the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 19 Sep 2022
| 19 Sep 2022
Effect of the presence of plateau pikas on the ecosystem services of alpine meadows
Ying Ying Chen
Huan Yang
Gen Sheng Bao
Xiao Pan Pang
Zheng Gang Guo
The activity of small mammalian herbivores influences grassland ecosystem services in arid and semi-arid regions. Plateau pika (Ochotona curzoniae) was considered to be a focal organism to investigate the effect of small mammalian herbivores on meadow ecosystem services in alpine regions. In this study, a home-range scale was used to measure the forage available to livestock, water conservation, carbon sequestration and soil nutrient maintenance (total nitrogen, phosphorus and potassium) in the topsoil layer, and a quadrat scale was used to assess the biodiversity conservation of alpine meadows. This study showed that the forage available to livestock and water conservation was 19 % and 16 % lower in the presence of plateau pikas than in their absence, and biodiversity conservation, carbon sequestration, soil nitrogen and phosphorus maintenance was 15 %, 29 %, 10 % and 8.9 % higher in the presence of plateau pikas than in their absence. In contrast, it had no impact on soil potassium maintenance of meadow ecosystems in alpine regions. The forage available to livestock, biodiversity conservation and soil nutrient maintenance of meadow ecosystems in alpine regions had maximum values as the disturbance intensity of plateau pikas increased; the water conservation tended to decrease linearly with the increasing disturbance intensity of plateau pikas. These results present a pattern of plateau pikas influencing the ecosystem services of meadow ecosystems in alpine regions, enriching our understanding of the small mammalian herbivores in relation to grassland ecosystem service.
- Article
(1649 KB) - Full-text XML
- BibTeX
- EndNote





Grasslands provide multiple ecosystem services, mainly including provisioning services of food and water, regulating services of carbon sequestration and water conservation, supporting services of soil nutrient maintenance and biodiversity conservation, and cultural services of landscapes and recreation tourism (Millennium Ecosystem Assessment, 2005; Bai and Cotrufo, 2022; Buisson et al., 2022; Strömberg and Staver, 2022). These ecosystem services sustain animal production, flora and fauna, and other human welfare (Costanza et al., 1997; Zhang et al., 2018; Dong et al., 2020); however, they are affected by multiple biotic factors, such as soil microbial communities (Van Eekeren et al., 2010), grazing by large herbivores (Lu et al., 2017) and the presence of small herbivores (Delibes-Mateos et al., 2011; Martínez-Estévez et al., 2013).
Small mammalian herbivores are common biotic factors (Davidson et al., 2012). These herbivores usually create extensive disturbances on grassland vegetation and soil (Pang et al., 2020a, b) by developing burrow systems (Delibes-Mateos et al., 2008; Sun et al., 2015), excreting feces and urine (Zhang et al., 2016), consuming plants (Eldridge and Myers, 2001; Liu et al., 2017), clipping tall plants (Zhang et al., 2020), and producing bare soil patches (Guo et al., 2012a, b; Yu et al., 2017a, b) or mounds (Yang et al., 2021). Previous studies have shown that the presence of prairie dogs (Cynomys ludovicianus) can increase the forage available to livestock, water conservation, carbon sequestration and biodiversity conservation of grassland ecosystems in arid regions (Ceballos et al., 1999; Martínez-Estévez et al., 2013), whereas the presence of European rabbit (Oryctolagus cuniculus) can decrease the forage available to livestock (Delibes-Mateos et al., 2008; Eldridge and Myers, 2001) and increase the biodiversity conservation (Delibes-Mateos et al., 2008) and nitrogen maintenance (Willott et al., 2000) of grassland ecosystems in semi-arid regions. In addition to grasslands in arid and semi-arid regions, vast alpine meadows exist in high-latitude and high-altitude regions throughout the world (Zhang et al., 2018; Dong et al., 2020). However, how small mammalian herbivores influence the ecosystem services in alpine meadows as much as they do in arid and semi-arid regions has not been well documented.
The plateau pika (Ochotona curzoniae) is a common, small mammalian herbivore that mainly lives in alpine meadows of the Qinghai–Tibetan Plateau (Smith and Foggin, 1999). This small mammalian herbivore with an average weight of 150 g is diurnally active and non-hibernating (Smith and Wang, 1991; Fan et al., 1999) and preferentially consumes dicotyledons (Zhao et al., 2013; Pang and Guo, 2017). Plateau pikas, a sexual monomorphism (Dobson et al., 1998), often construct a family warren with numerous burrow entrances and develop a complex burrow system with an average length and depth of 13 m and 30 cm (Fan et al., 1999). This mammalian herbivore is social and philopatric (Dobson et al., 1998), and its young offspring stay with their family during their birth year (Wang et al., 2020). Plateau pikas are generally considered a pest in China (Harris, 2010; Pang and Guo, 2017) as they often exacerbate the degradation of alpine meadows (Liu et al., 2013; Zhang et al., 2016). However, some studies have argued that plateau pika is a key species in alpine meadow ecosystems (Smith and Foggin, 1999; Delibes-Mateos et al., 2011). This disagreement has encouraged professionals to re-evaluate the role of plateau pikas in alpine meadow ecosystems. Thus, the effects of the presence of plateau pikas on ecosystem services of alpine meadows allow insight into the role of plateau pikas in alpine meadow ecosystems. Previous studies have demonstrated that the presence of plateau pikas decreases (Liu et al., 2013) or has no significant effect on plant biomass (Pang and Guo, 2017), increases (Liu et al., 2017; Pang and Guo, 2017) or decreases (Sun et al., 2015) plant-species richness, and increases (Yu et al., 2017a; Pang et al., 2020a, b) or decreases (Sun et al., 2015) soil carbon and nutrients. In addition, previous studies have shown that the disturbance intensity of plateau pikas affects plant-species richness and soil nutrient stocks of alpine meadows (Yu et al., 2017a; Pang and Guo, 2018). These findings imply that plateau pikas may have an impact on the ecosystem services of alpine meadows. Thus, further studies are needed to test whether the presence of plateau pikas and their disturbance intensity influence the ecosystem services of alpine meadows, which can enrich our understanding of the presence of small mammalian herbivores in relation to grassland ecosystem services.
Since soil carbon and nutrients differ between vegetated and bare soil patches in the home range (Yu et al., 2017b), Pang et al. (2020a, b) proposed that the home-range scale is a better proxy than the quadrat scale to estimate the complete effects of the presence of plateau pikas on soil carbon and nutrient stocks. Although the provisioning, regulation, support and cultural services of alpine meadows can be estimated by multiple indicators (Egoh et al., 2012; Brown et al., 2014), one or two can be used to verify whether the presence and intensity of plateau pikas influence each ecosystem service. In previous studies, palatable plant biomass for livestock has been used to evaluate the provisioning services (Martínez-Estévez et al., 2013; Wen et al., 2013), soil-water storage and soil organic carbon stock have been used to evaluate the regulating services (Wen et al., 2013; Li and Xie, 2015; Tang et al., 2019), and plant-species richness and soil total nutrient stocks can be used to evaluate the supporting services (Wen et al., 2013). Notably, cultural services are particularly related to the spatial scale, as many are perceived visually over distant views (Norton et al., 2012). The plateau pika is territorial, and its habitat use is patchy within a given area (Pang et al., 2020a), which leads to mismatches between the spatial scale and cultural services (De Groot et al., 2010). Therefore, the present study used ecological services of forage available to livestock, water conservation, carbon sequestration and soil nutrient maintenance, and biodiversity conservation to test how the presence of plateau pikas influences the ecosystem services of alpine meadows across five sites. In this study, we hypothesized that (1) the presence of plateau pikas leads to lower forage available to livestock because of lower palatable plant biomass in the presence of small mammalian herbivores, (2) the presence of plateau pikas leads to higher water conservation and carbon sequestration because small mammalian herbivores can increase soil-water storage and carbon stocks, and (3) the presence of plateau pikas leads to higher biodiversity conservation and soil nutrient maintenance because small mammalian herbivores can increase plant-species richness and soil nutrient stocks.
2.1 Study site descriptions
Plateau pikas can live in various habitats with different soil types, topographies and microclimates on the Qinghai–Tibetan Plateau. To determine how the presence of plateau pikas generally influences the ecosystem services of alpine meadows, five survey sites were selected in Luqu (34∘15′51′′ N, 102∘22′12′′ E), Gangcha (37∘36′12′′ N, 100∘26′26′′ E), Haiyan (36∘57′50′′ N, 100∘54′33′′ E), Qilian (37∘43′26′′ N, 100∘34′48′′ E) and Gonghe (36∘43′48′′ N, 99∘47′11′′ E) counties on the Qinghai–Tibetan Plateau. These five survey sites have a similar typical plateau continental climate, with elevations ranging from 3194 m at the Gonghe survey site to 3550 m at the Luqu survey site. Based on 5-year weather data, the mean annual temperatures are 3.1, 0.9, 1.9, 2.2 and 3.3 ∘C at Luqu, Gangcha, Haiyan, Qilian and Gonghe, respectively, of which the average temperatures in the warm season are 9.3, 8.3, 9.6, 10.3 and 9.9 ∘C and in the cold season are −3.1, −6.5, −5.8, −5.9 and −3.4 ∘C. The mean annual precipitation is 439.5, 258.9, 257.4, 257.0 and 239.8 mm, of which the warm season accounts for 83.4 %, 92.8 %, 89.3 %, 91.5 % and 91.4 % at Luqu, Gangcha, Haiyan, Qilian and Gonghe, respectively. According to the Chinese soil classification system (Gong, 2001), the soil type at each site is alpine meadow soil, similar to Cambisol in the World Reference Base (WRB) soil classification system.
Animal husbandry is the dominant use of alpine meadows on the Qinghai–Tibetan Plateau, and herders traditionally graze their livestock seasonally on cold and warm grasslands. The survey sites in this study were all situated in cold grasslands, in which alpine meadows were fenced from mid-April to September, and fences were opened to grazing yaks from mid-October to early April (Zhang et al., 2020). All field data were collected in August, when the annual population of plateau pikas was the highest, and reproduction had largely ceased (Qu et al., 2013; Pang et al., 2020a, b). In addition, the growing season for plants is short on the Qinghai–Tibetan Plateau, and some plants do not turn green until July. Therefore, sampling in August is optimal because August is a good time to identify all plants and ensure an accurate survey of the plant species. Notably, the small burrowing herbivore at each survey site was only plateau pikas.
2.2 Field survey design
The adult dispersal of plateau pikas is a gradual process (Pang et al., 2020a); it is easy to identify reference sites without plateau pikas, even though these sites might be potential as suitable habitats. In this study, a home-range scale was used to calculate the forage available to livestock, water conservation, carbon sequestration and soil nutrient maintenance, and a quadrat scale was used to calculate the biodiversity conservation.
A stratified random and paired design was used to select plots. The home range of the plateau pika was approximately 1262.5 m2 (Fan et al., 1999), and the plot size was 35 × 35 m, which was similar to the average area of the plateau pika's home range. At each of the five sites, this study first selected 10 plots where plateau pikas were present or where active burrow entrances were observed. The second plot was identified along the road when the first plot with plateau pikas was selected. The distance between the two plots with plateau pikas was more than 3 km, which ensured that plateau pikas of the same family would not appear in two plots at the same time. Second, a paired adjacent plot without plateau pikas and active burrow entrances was selected for each plot with plateau pikas. The plots without plateau pikas were in any direction of plots with plateau pikas. The distance between each plot with plateau pikas and its paired plot without plateau pikas ranged from 500 to 1000 m. If the distance between each paired plot was too close, the plateau pikas could move between plots with and without plateau pikas. To ensure that each plot with plateau pikas was paired with a plot without plateau pikas, each paired plot shared the alpine meadow with the same dominant plant, with no obvious differences in soil type, topography or microclimate. In total, there were 10 paired plots at each site and 100 plots across 5 sites, including 50 with and 50 without plateau pikas. Each paired plot shared the same grazing intensity during the cold season; however, 50 paired plots consisted of different yak grazing intensity, and this can permit the general pattern relating to the effect of plateau pika disturbance on alpine meadow ecosystem services.
2.3 Field sampling
Field surveys and sampling were conducted in early August 2020. First, the active burrow entrance at each plot with plateau pikas was estimated by the “plugging tunnels method”, in which the burrow entrances were plugged with hay for 3 d (Sun et al., 2015), and the number of plugs cleared by the plateau pikas to allow access to the meadow surface was recorded (Guo et al., 2012a). The average number of burrow entrances with cleared plugs after 3 d was taken as the density of active burrow entrances. For plots with plateau pikas, the density of active burrow entrances was used as a proxy for the intensity of the disturbance (Guo et al., 2012a; Sun et al., 2015). Second, this study was restricted to plateau pikas in relation to the ecosystem services of alpine meadows. However, bare soil patches caused by other factors (no plateau pikas) simultaneously existed on the vegetated surface in the presence or absence of plateau pikas. To actually quantify the effect of plateau pikas on ecosystem services of alpine meadows, this study only measured the area of bare soil patches caused by plateau pikas, although there exist multiple types of bare soil patches in alpine meadows. The bare soil patches caused by plateau pikas are easy to identify because one bare soil patch caused by plateau pikas is paired with a visible burrow entrance (Pang et al., 2021a). Other bare soil patches are not paired with a visible burrow entrance. The area of each bare soil patch (created by plateau pikas) in the plot with plateau pikas was measured. Each bare soil patch was identified as a regular shape or irregular shape. If one bare soil patch was identified as a regular shape, such as a rectangle, circle or trapezoid, a ruler was used to measure its length, width, height, diameter, and upper and lower bottom, and these data were used to calculate the area of that bare soil patch. If one bare soil patch was identified as an irregular shape, this bare soil patch was divided into several regular shapes; the areas of these regular shapes were calculated, respectively, and the area sum of these regular shapes from the irregular bare soil patch was considered to be the area of that irregular bare soil patch (Han et al., 2011). Then, the sum of all bare soil patch areas in each plot with plateau pikas was calculated and defined as the bare soil area for that plot. Third, five vegetated quadrats (1×1 m) were placed on the vegetated surface approximately 8 m apart along the shape of a W pattern in all plots (with or without plateau pikas). In plots with plateau pikas, if the quadrat was justly covered with the bare patches caused by plateau pikas, the quadrat was slightly moved to avoid it; if the quadrat was justly covered with the bare patches caused by other factors, the quadrat was not moved. Fourth, alpine meadows in plots with plateau pikas consisted of bare and vegetated surface, and a bare soil patch was selected as a paired bare soil quadrat for each vegetated quadrat in the plot with plateau pikas, and the distance between each paired bare soil quadrat and vegetated quadrat was as short as possible (less than 1 m). Bare-soil-patch quadrats and each vegetated quadrat were beneficial to accurately measure the soil nutrients, carbon concentrations and plant biomass, which reflected the effect of the presence of plateau pika on ecosystem services in alpine meadows by comparing the parameters between plots with and without plateau pikas at the home-range scale. Thus, there were five paired quadrats, consisting of five vegetated and five bare soil quadrats in each plot with plateau pikas. Additionally, there were five vegetated quadrats in each plot without plateau pikas, since this study focused on bare soil patches induced by plateau pikas.
In each vegetated quadrat of the plot with or without plateau pikas, all vascular plant species were identified, and the number of plant species was recorded as plant-species richness. Then, all plants rooted in a quadrat were harvested into palatable and unpalatable plants, in which palatable plants or unpalatable plants were for yak and Tibetan sheep (Pang and Guo, 2017). Finally, all palatable plant samples were placed in envelopes and transported to the laboratory.
Generally, most burrows derived from the activities of plateau pikas are less than 20 cm in depth (Yu et al., 2017b), although a few burrows extend to depths of 60 cm (Fan et al., 1999). In addition, the majority of plant roots in alpine meadows of the Qinghai–Tibetan Plateau are in the top 20 cm of the soil. The soil samples were collected at a depth of 20 cm. Soil samples were collected from vegetated and bare soil quadrats for each plot with plateau pikas and vegetated quadrats for each plot without plateau pikas. Before collecting the soil samples, plants and litter were removed from the soil surface. First, a 5 cm diameter soil auger was used to collect soil samples, which were used to measure soil organic carbon and soil nutrient concentrations (total nitrogen, phosphorus and potassium). Second, soil profiles in each quadrat were obtained using a spade, and a stainless-steel cutting ring (with a volume of 100 cm3) was used to collect soil cores to determine soil bulk density and soil-water content. Soil samples used to determine soil bulk density were packed into aluminum boxes with recorded weights, and each aluminum box was numbered. The aluminum boxes containing fresh soil were immediately weighed, recorded, stored at 4 ∘C and then transported to the laboratory. Thus, in this study, 10 soil samples were collected to analyze the soil carbon, nitrogen, phosphorus, and potassium concentrations, and 10 soil samples were obtained to determine the soil bulk density in each plot with plateau pikas. Because this study is constricted with bare soil patch caused by plateau pikas, bare soil quadrats was not considered in plot without plateau pikas; therefore, five soil samples were used to determine the soil carbon, nitrogen, phosphorus, and potassium concentrations, and five samples were obtained for the analysis of soil bulk density in each plot without plateau pikas. Five soil samples in each plot were individually measured. The average value of five soil samples in one plot was considered to be the representative data of that plot.
2.4 Analysis of samples
In the laboratory, palatable plant samples were dried in an oven at 80 ∘C for 48 h and weighed. The soil samples used to measure soil bulk density and soil-water content were dried to a constant weight at 105 ± 2 ∘C, the aluminum boxes with dry soil were weighed, and the values were recorded. The soil samples used to measure soil organic carbon and total nitrogen, phosphorus and potassium concentrations were air-dried; gravel and roots were manually picked out; and the proportion of gravel larger than 2.0 mm in the soil sample was determined by passing through a 2.0 mm sieve. Finally, soil samples were sieved at 0.15 mm to analyze soil organic carbon, nitrogen, phosphorus and potassium concentrations in the laboratory. Soil organic carbon was measured using the dichromate heating–oxidation (Nelson and Sommers, 1982). Soil total nitrogen concentration was measured using the Kjeldahl procedure. Soil total phosphorus concentration was measured using the Molybdenum blue colorimetric method. Soil total potassium concentration was measured using flame photometry.
Soil bulk density, soil organic carbon and nutrient concentrations (total nitrogen, phosphorus and potassium) were used to calculate the soil organic carbon, total nitrogen, phosphorus and potassium stocks. Soil bulk density and soil-water content were used to calculate the soil-water storage (Jia et al., 2020).
2.5 Calculations
The bare soil area consisted of all bare soil patches, and the vegetated surface area was estimated from the plot areas minus the bare soil areas. This study only measured the area of bare soil patches caused by plateau pikas, although there were other kinds of bare soil patches in alpine meadows. Therefore, bare soil areas caused by plateau pikas were considered to be zero in each plot without plateau pikas, and the vegetated surface area was considered to be 100 %.
The palatable plant biomass was calculated using the following equation:
where GB, Bq and δva are the palatable plant biomass of the plot, palatable plant biomass on the quadrat scale (g m−2) and vegetated surface area, respectively.
The plant-species richness in a quadrat (1 × 1 m) was measured using the species number of each quadrat.
Soil-water storage was determined using the method recommended by Jia et al. (2020), and it was calculated by the following equation:
where SWSpika, SWCBA, BDBA and θBA are soil-water storage in a plot with plateau pikas, water content (g kg−1), soil bulk density (g cm−3) and soil fraction of gravel larger than 2 mm in bare soil areas of plots with plateau pikas, respectively; BA is the percentage of bare soil areas in plots with plateau pikas; SWCVA, BDVA and θVA are water content (g kg−1), soil bulk density (g cm−3) and soil fraction of gravel larger than 2 mm in vegetated areas of a plot with plateau pikas, respectively; T is soil thickness (20 cm); and VA is the percentage of vegetated surface area in plots with plateau pikas. SWCBA and SWCVA were measured by the oven drying method.
where SWCno pika, SWCno pika, BDno pika and θno pika are soil-water storage in a plot without plateau pikas, soil-water content (g kg−1), soil bulk density (g cm−3) and soil fraction of gravel larger than 2 mm in plots without plateau pikas, respectively, and T is soil thickness (20 cm).
The soil organic carbon stock per plot was estimated using the method described by Pang et al. (2020b), and it was calculated by the following equation:
where SOCSpika is soil organic carbon stock in a plot with plateau pikas (kg m−2); SOCBA, BDBA and θBA are soil organic carbon concentration (g kg−1), soil bulk density (g cm−3) and soil fraction of gravel larger than 2 mm in bare soil areas of plots with plateau pikas, respectively; BA is the percentage of bare soil areas in plots with plateau pikas; SOCVA, BDVA and θVA are organic carbon concentration (g kg−1), soil bulk density (g cm−3) and soil fraction of gravel larger than 2 mm in vegetated areas of a plot with plateau pikas, respectively; T is soil thickness (20 cm); and VA is the percentage of vegetated surface area in plots with plateau pikas.
where SOCSno pika is soil organic carbon stock in the plot without plateau pikas (kg m−2); SOCno pika, BDno pika and θno pika are soil organic carbon concentration (g kg−1), soil bulk density (g cm−3) and soil fraction of gravel larger than 2 mm in plots without plateau pikas, respectively; and T is soil thickness (20 cm).
The soil total nitrogen, phosphorus and potassium stocks per plot were quantified using the method described by Pang et al. (2020a), and it was calculated by the following equation:
where SNSipika is soil total nitrogen, phosphorus and potassium stock in plots with plateau pikas (kg m−2); SNiBA, BDBA and θBA are soil nutrient concentration (g kg−1), soil bulk density (g cm−3) and soil fraction of gravel larger than 2 mm in bare soil areas of plots with plateau pikas, respectively; BA is the percentage of bare soil areas in plots with plateau pikas; SNiVA, BDVA and θVA are soil nutrient concentration (g kg−1), soil bulk density (g cm−3) and soil fraction of gravel larger than 2 mm in vegetated areas of a plot with plateau pikas, respectively; T is soil thickness (20 cm); and VA is the percentage of vegetated surface area in plots with plateau pikas.
where SNSino pika is soil total nitrogen, phosphorus and potassium stock in the plot without plateau pikas (kg m−2); SNino pika, BDno pika and θno pika are soil nutrient concentration (g kg−1), soil bulk density (g cm−3) and soil fraction of gravel larger than 2 mm in plots without plateau pikas, respectively; and T is soil thickness (20 cm).
2.6 Data analysis
Data from 50 disturbed plots and 50 undisturbed plots were used to examine the difference in ecosystem services of alpine meadows between the presence of plateau pikas and the absence of plateau pikas, and then data from 50 disturbed plots were used to examine the responses of each ecosystem service of alpine meadows to the disturbance intensity of plateau pikas.
All data variables (palatable plant biomass, plant-species richness, soil-water storage, soil organic carbon stock, soil total nitrogen stock, soil total phosphorus stock and soil total potassium stock) were assessed for the normality and homogeneity by using the Shapiro–Wilk test. If necessary, the data were base-10 log-transformed to fit the assumption of normality and homogeneity for further variance analysis.
A linear mixed model (LMM) with the function “lmer” from the lme4 package was used to examine differences in palatable plant biomass, plant-species richness, soil-water storage, soil organic carbon stock, soil total nitrogen stock, soil total phosphorus stock and soil total potassium stock between the presence and absence of plateau pikas across the five sites. In linear mixed models, the abovementioned parameters acted as response variables, the absence and presence were introduced as fixed factors, and the paired plots nested within each site as a random factor.
To clarify the responses of palatable plant biomass, plant-species richness, soil-water storage, soil organic carbon stock, soil total nitrogen stock, soil total phosphorus stock and soil total potassium stock to the disturbance caused by plateau pikas, a linear model (LM) was used to examine the relationships between these variables and active burrow entrance densities in all plots with plateau pikas. The densities of active burrow entrances by plateau pikas were considered to be the fixed factor and were used to construct the regression analysis between palatable plant biomass, plant-species richness, soil-water storage, soil organic carbon stock, soil total nitrogen stock, soil total phosphorus stock and active burrow entrance densities. To select the final regression models, likelihood ratio tests were used to compare simple linear regression and polynomial regression models. After likelihood ratio tests, the models with p<0.05 and the smaller Akaike information criterion (AIC) were used as the final regression models.
Bonferroni's test was used to adjust P values and to correct for experiment-wise error rates. All statistical analyses were performed with R 4.0.2 (R Foundation for Statistical Computing, Vienna, Austria).
3.1 Effects of the presence of plateau pikas on the ecosystem services of alpine meadows
The palatable plant biomass (Fig. 1a) and soil-water storage (Fig. 1b) were 19 % and 16 % lower in the plots with plateau pikas than in the plots without plateau pikas, whereas soil organic carbon stock (Fig. 1c), plant-species richness (Fig. 1d), soil total nitrogen (Fig. 1e) and total phosphorus stocks (Fig. 1f) in the plots with plateau pikas were 29 %, 15 %, 10 % and 8.9 % higher than those in the plots without plateau pikas. In addition, there was no difference in the soil total potassium stock between the plots with and without plateau pikas (Fig. 1g).
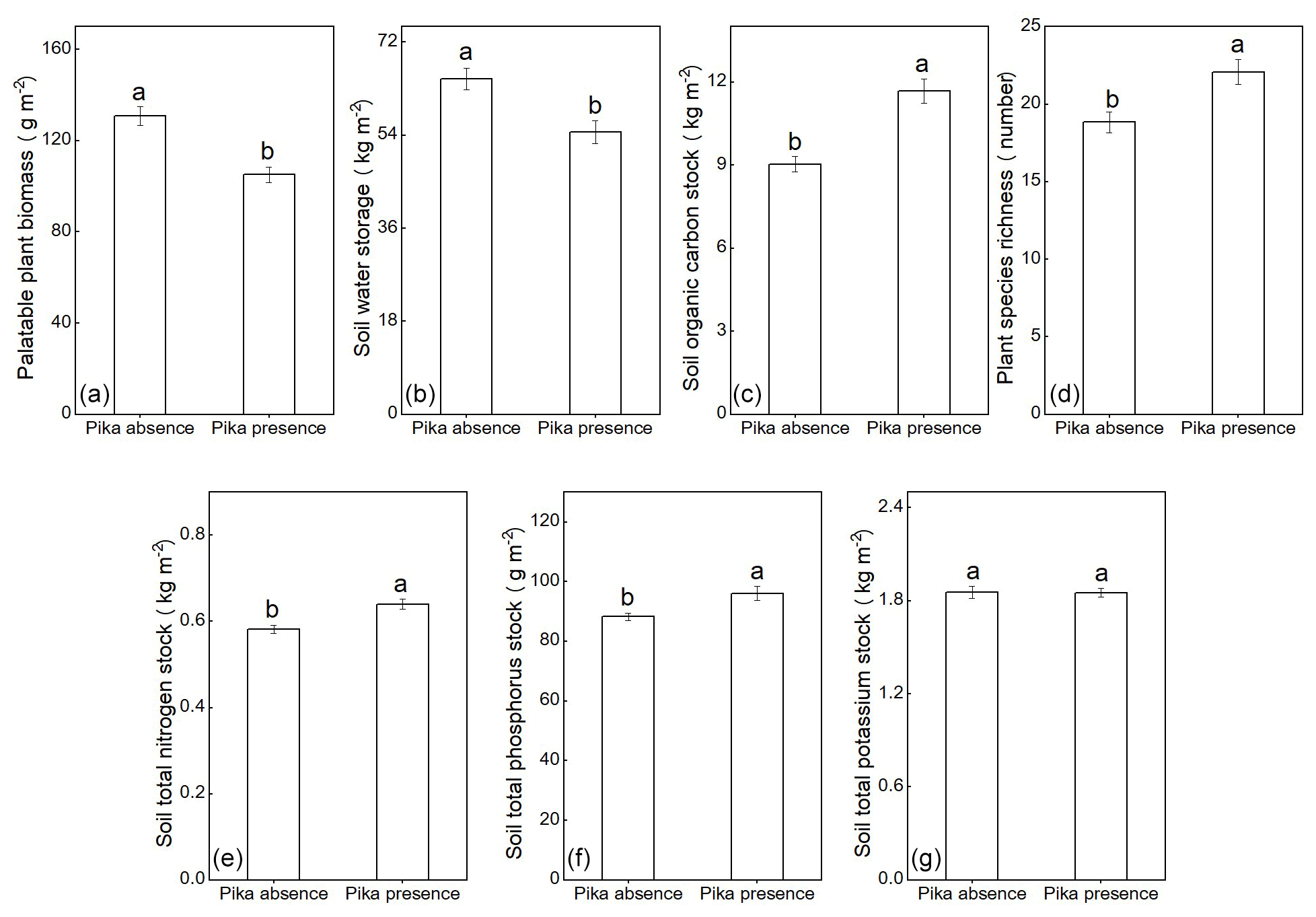
Figure 1Palatable plant biomass (F = 46, p<0.001) (a), soil-water storage (F = 35, p<0.001) (b), soil organic carbon stock (F = 88, p<0.001) (c), plant-species richness (F = 64, p=0.003) (d), soil total nitrogen stock (F = 22, p<0.001) (e), soil total phosphorus stock (F = 12, p=0.004) (f) and soil total potassium stock (F = 0.03, p=0.88) (g) of plots with and without plateau pikas (mean ± standard error). Lower case represents a significant difference between the absence and presence of pika based on an LMM.
3.2 Effects of plateau pikas' disturbance intensity on the ecosystem services of alpine meadows
The palatable plant biomass (Fig. 2a), soil organic carbon stock (Fig. 2c), plant-species richness (Fig. 2d), soil total nitrogen (Fig. 2e) and phosphorus (Fig. 2f) stocks had maximum values as the plateau pikas' disturbance intensity increased, while the soil-water storage of the topsoil layer (Fig. 2b) decreased linearly with increasing disturbance intensity of plateau pikas. In addition, the disturbance intensity of plateau pikas had no obvious relationship with soil total potassium (Fig. 2g).
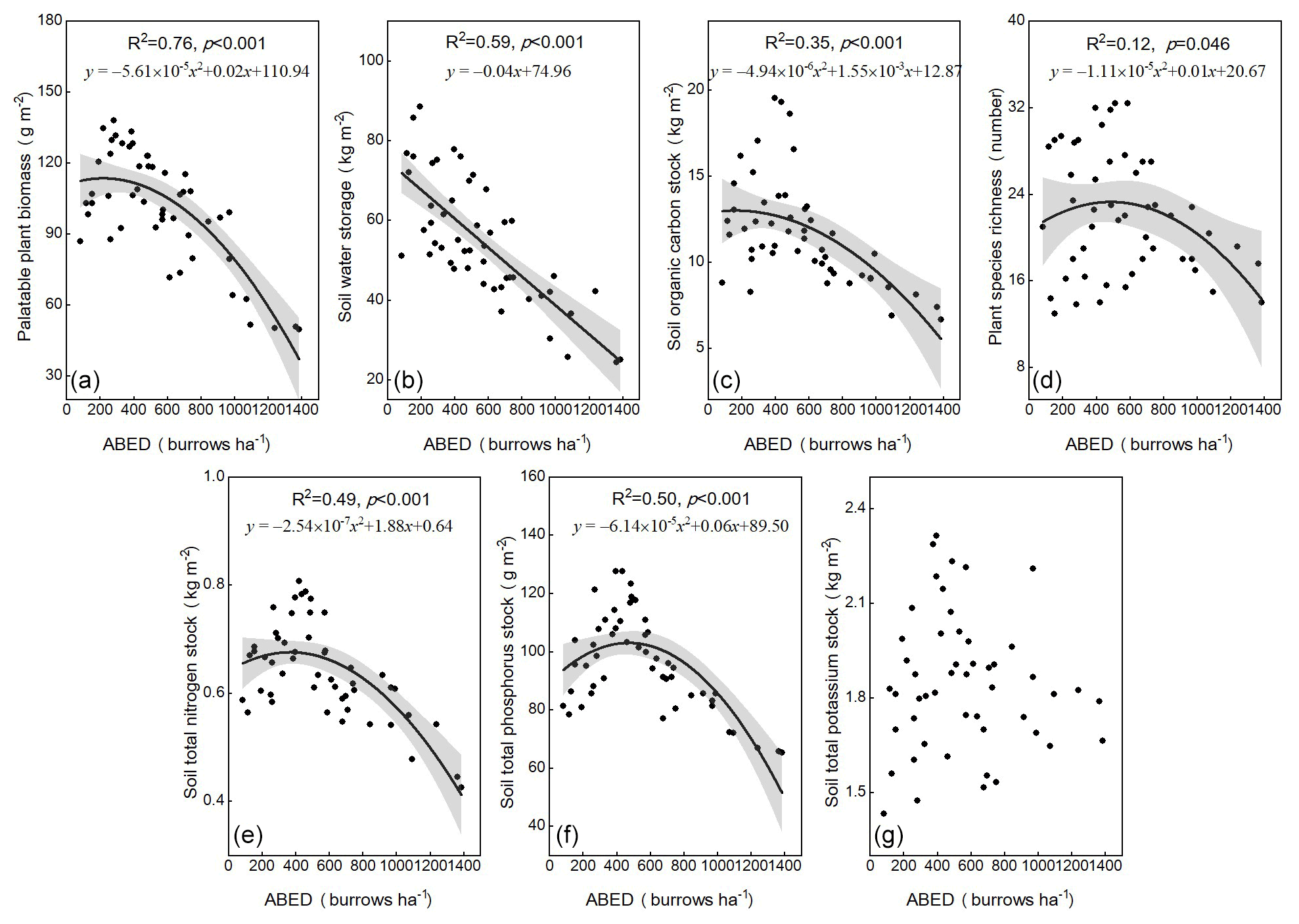
Figure 2The palatable plant biomass (F = 69) (a), soil-water storage (F = 69) (b), soil organic carbon stock (F = 13) (c), plant-species richness ( F = 3.3) (d), soil total nitrogen stock (F = 23) (e), soil total phosphorus stock (F = 24) (f) and soil total potassium stock (g) for different disturbance intensity of plateau pikas based on linear models (LMs). An adjusted local smoothed regression line (black) with its 95 % confident interval (gray) was used to determine the relationship between the disturbance intensity of plateau pikas and the indicators above. ABED: active burrow entrance densities.
Prairie dogs and European rabbits have been shown to affect grassland ecosystem services in arid and semi-arid regions (Delibes-Mateos et al., 2011; Martínez-Estévez et al., 2013). This study combined the home-range scale and the quadrat scale to test how the presence of plateau pikas and their disturbance intensity influence the ecosystem services of alpine meadows and found that the presence of plateau pikas and their disturbance intensity indeed impact the ecosystem services of alpine meadows, similar to prairie dogs and European rabbits in grassland ecosystem services in arid and semi-arid regions.
Lower palatable plant biomass in the presence of plateau pikas indicates that the presence of plateau pikas reduces the forage available to livestock, which is consistent with the results of European rabbits in semi-arid regions (Eldridge and Myers, 2001; Delibes-Mateos et al., 2008) and is not consistent with results from prairie dogs in arid regions (Martínez-Estévez et al., 2013). Prairie dogs benefit perennial plants in arid grasslands, in which blue grama (Bouteloua gracilis) and vine mesquite (Panicum obtusum) are palatable perennials for livestock (Sierra-Corona et al., 2015), whereas European rabbits increase unpalatable plants (Marrubium vulgare and Colchicum melitensis) because they prefer grasses (Leigh et al., 1989; Eldridge and Myers, 2001). Plateau pikas enable more unpalatable broad-leaved plants to grow in alpine meadows (Pang and Guo, 2018) and can bury many plants (Pang and Guo, 2017). However, their consumption patterns can benefit the growth of palatable plants (Pang and Guo, 2017) because plateau pikas preferentially consume unpalatable dicotyledons (Zhao et al., 2013; Pang and Guo, 2017). The tradeoff between the decrease and increase in palatable plant biomass contributes to a negative effect on palatable plant biomass on a home-range scale, resulting in a decrease in the forage available to livestock. These results demonstrate that the presence of small mammalian herbivores affects the forage available to livestock of grassland ecosystems may be related to environmental conditions. Specific performance is that the presence of small mammalian herbivores is disadvantageous to the forage available to livestock in semi-arid and alpine regions, but it is beneficial to forage available to livestock in arid regions.
The presence of plateau pikas has different impacts on regulating services of alpine meadows, when assessed by different indicators. The presence of plateau pikas leads to lower soil-water storage, resulting in a decrease in the water conservation, whereas the presence of plateau pikas can lead to higher soil organic carbon stock, implying that the presence of plateau pikas can increase the carbon sequestration. Lower water conservation of alpine meadows in relation to the presence of plateau pikas is consistent with the effect of the presence of European rabbits on the water conservation of grasslands in semi-arid regions (Eldridge et al., 2010), whereas it is inconsistent with the presence of prairie dogs in relation to the water conservation in arid regions (Martínez-Estévez et al., 2013). This difference is ascribed to evaluation indicators for the water conservation. The water infiltration rate is considered to be an index to evaluate the effect of prairie dogs on the water conservation of grasslands in arid regions (Martínez-Estévez et al., 2013). In contrast, the water storage of topsoil is used to evaluate the effects of European rabbits and plateau pikas on the water conservation of grasslands in semi-arid grasslands and alpine meadows (Eldridge et al., 2010). The activities of European rabbits and plateau pikas can reduce the crust cover of grasslands and increase water infiltration from topsoil to deep soil in semi-arid regions (Eldridge et al., 2010; Li et al., 2015), contributing to a negative effect on the water conservation in the topsoil layer. This study shows that the presence of plateau pikas leads to higher carbon sequestration in alpine meadows, similar to the effect of the presence of prairie dogs in arid regions (Martínez-Estévez et al., 2013) and European rabbits in semi-arid regions (Delibes-Mateos et al., 2011). Plateau pikas can input extra organic matter through the deposition of uneaten food (Liu et al., 2009; Zhang et al., 2016; Yu et al., 2017a) and the excretion of urine and feces (James et al., 2009; Yu et al., 2017b), which increases the soil organic carbon stock and contributes to an increase in the carbon sequestration of alpine meadows. These results indicate that the presence of small mammalian herbivores can increase the carbon sequestration of grasslands.
Higher plant-species richness in the presence of plateau pikas shows that the presence of plateau pikas can lead to higher biodiversity conservation, similar to the effect of European rabbits in semi-arid regions (Delibes-Mateos et al., 2008) and prairie dogs in arid regions (Davidson et al., 2012). The mechanisms by which small mammalian herbivores lead to higher plant-species richness have been discussed in many previous studies (Zhang et al., 2020; Pang et al., 2021b). The presence of plateau pikas can lead to higher soil total nitrogen and total phosphorus stocks, demonstrating that plateau pikas can increase the soil nitrogen and phosphorus maintenance. In addition, there was no difference in the soil total potassium stock between the areas with and without plateau pikas, indicating that the presence of plateau pikas had no effect on the soil potassium maintenance. This effect was also observed with prairie dogs and European rabbits in arid (Delibes-Mateos et al., 2011) and semi-arid regions (Delibes-Mateos et al., 2008; Willott, 2001). Some of the following factors explain the higher soil nitrogen and phosphorus stocks caused by plateau pikas. The presence of plateau pikas can increase the input of soil organic material (Liu et al., 2013; Zhang et al., 2016; Pang et al., 2020a). Secondly, the presence of plateau pikas can result in higher organic nitrogen and phosphorus stocks (Yu et al., 2017b), which contributes to higher soil nitrogen and phosphorus maintenance. These results suggest that a general pattern can be identified regarding the effect of the presence of small mammalian herbivores on the supporting services of biodiversity conservation, soil nitrogen and phosphorus maintenance.
This study also shows that the disturbance intensity of plateau pikas also affects the forage available to livestock, biodiversity conservation, water conservation, carbon sequestration, and soil total nitrogen and phosphorus maintenance, and these effects are related to disturbance intensity of plateau pikas. In this case, the active burrow entrances caused by plateau pikas were used to indicate the disturbance intensity of plateau pikas. However, active burrow entrances in disturbed plots were greatly changeable. This study just uses the field survey data in this experiment to simulate the effect of disturbance intensity of plateau pikas on the palatable plant biomass; soil organic carbon stock; plant-species richness; and soil total nitrogen, phosphorus and potassium stocks. As found in plant-species richness and aboveground plant productivity (Dial and Roughgarden, 1998; Gao and Carmel, 2020), the response of plant-species richness and palatable plant biomass to the disturbance intensity of plateau pikas follows the pattern for the intermediate disturbance hypothesis in this study. In addition, the soil organic carbon stock and soil total nitrogen and phosphorus stocks at the home-range scale also support the intermediate disturbance hypothesis. However, the topsoil-water storage does not confirm the intermediate disturbance hypothesis.
At lower disturbance intensity, stronger competition of dominant sedges often prevents the grass from growing well (Pang and Guo, 2018) and rare plants from coexisting (Wang et al., 2012), which leads the forage available to livestock and biodiversity conservation of alpine meadows to be maintained at a low level. Although the presence of plateau pikas can increase the input of soil organic matter, this increase is low (Pang and Guo, 2017; Pang et al., 2020b), which enables the soil organic carbon sequestration and soil nitrogen and phosphorus maintenance of alpine meadows to maintain a relatively low level.
At intermediate disturbance intensity, the activities of plateau pikas improve the growth potential of grass plants (Wang et al., 2012) and increase the input of organic matter, soil total nitrogen (Li et al., 2014) and organic carbon accumulation (Yu et al., 2017b), which contributes to higher biodiversity conservation, forage available to livestock, carbon sequestration, and soil total nitrogen and phosphorus maintenance services.
At higher disturbance intensity of plateau pikas, frequent bioturbation can put all species at risk of going extinct (Dial and Roughgarden, 1998). Low soil-water content in alpine meadows (Liu et al., 2013) only sustains the xerophytes and mesophytes, most of which are unpalatable (Pang and Guo, 2018). This contributes to relatively lower forage available to livestock and biodiversity conservation. Low vegetation biomass decreases the input resources of soil organic matter (Sun et al., 2015; Pang and Guo, 2017), contributing to a decrease in the soil organic carbon sequestration and soil nitrogen and phosphorus maintenance of alpine meadows.
Additionally, the linearly negative relationship between the water conservation of alpine meadows and disturbance intensity of plateau pikas is ascribed to evaporation and more water infiltration on bare soil patches, as the amount of water evaporation and infiltration tends to increase as the area of bare soil increases (Liu et al., 2013).
Together with previous studies (Delibes-Mateos et al., 2011; Martínez-Estévez et al., 2013), this study demonstrates that the presence of small mammalian herbivores has similar impacts on the biodiversity conservation, soil nutrient maintenance and carbon sequestration of grasslands throughout the arid, semi-arid and alpine regions, whereas the effects of the presence of small mammalian herbivores on the forage available to livestock and water conservation are dependent on environmental conditions. This study further verifies that the disturbance intensity of plateau pikas also has a significant impact on the ecosystem services of alpine ecosystems. These results concur with the findings in research fields of small mammalian herbivores in relation to grassland ecosystem services.
This study focused on plateau pikas to investigate the responses of forage available to livestock, water conservation, carbon sequestration, soil nutrient maintenance and biodiversity conservation of meadow ecosystems to the presence of a small mammalian herbivore and its disturbance intensity across five sites. This will provide insight into the relationship between small mammalian herbivores and ecosystem services of grasslands. The results of this study showed that the presence of plateau pikas led to higher biodiversity conservation, soil nitrogen and phosphorus maintenance, and carbon sequestration of alpine meadows, whereas it led to lower forage available to livestock and water conservation of alpine meadows. Furthermore, this study found that the effect of plateau pikas' disturbance intensity on the forage available to livestock, biodiversity conservation, soil maintenance of nitrogen and phosphorus, and carbon sequestration also conformed to the moderate disturbance hypothesis. These results verified that plateau pikas could affect the ecosystem services of meadow ecosystems in alpine regions and present a relatively complete pattern of small mammalian herbivores influencing grassland ecosystem services.
The code used in this study can be accessed by contacting the first author (chenyy2019@lzu.edu.cn) based on a reasonable request.
Data used in this study are archived by the authors and are available on request.
YYC and ZGG conceived the ideas and designed the methodology; YYC, XPP, GSB and HY collected the data; YYC analyzed the data; YYC and ZGG wrote the manuscript. All of the authors contributed critically to the drafts and gave their final approval for publication.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The authors would like to thank Jing Zhang, Qian Wang, Hai Peng Xu, Wen Na Zhang, Juan Wang, Ding Yang, Jie Li, Fu Yun Qiao, Di Gang Zhi, Hao Hao Qi, Gan Lin Feng and Yuan Yuan Duan from Lanzhou University for the contributions made to this study through their field assistance and laboratory analysis.
This study was funded by the National Natural Science Foundation of China (grant no. 32171675); the Key Laboratory of Superior Forage Germplasm in the Qinghai–Tibetan Plateau (grant no. 2020-ZJ-Y03); and the Open Project of the State Key Laboratory of Plateau Ecology and Agriculture, Qinghai University.
This paper was edited by Paul Stoy and reviewed by Shixiong Li, Xiaojun Yu and one anonymous referee.
Bai, Y. F. and Cotrufo, M. F.: Grassland soil carbon sequestration: Current understanding, challenges, and solutions, Science, 377, 603–608, https://doi.org/10.1126/science.abo2380, 2022.
Buisson, E., Archibald, S., Fidelis, A., and Suding, K. N.: Ancient grasslands guide ambitious goals in grassland restoration, Science, 377, 594–598, https://doi.org/10.1126/science.abo4605, 2022.
Brown, C., Reyers, B., Ingwall-King, L., Mapendembe, A., Nel, J., O'Farrell, P., Dixon, M., and Bowles-Newark, N. J.: Measuring Ecosystem Services: Guidance on developing ecosystem service indicators, UNEP World Conservation Monitoring Centre, 72, https://doi.org/10.13140/RG.2.2.11321.83043, 2014.
Ceballos, G., Pacheco, J., and List, R.: Influence of prairie dogs (Cynomys ludovicianus) on habitat heterogeneity and mammalian diversity in Mexico, J. Arid. Environ., 41, 161–172, https://doi.org/10.1006/jare.1998.0479, 1999.
Costanza, R., D'Arge, R., De Groot, R., Farber, S., Grasso, M., Hannon, B., Limburg, K., Naeem, S., O'Neill, R., Paruelo, J., Raskin, R. G., Sutton, P., and Van Den Belt, M.: The value of the world's ecosystem services and natural capital, Nature, 387, 253–260, https://doi.org/10.1038/387253a0, 1997.
Davidson, A. D., Detling, J. K., and Brown, J. H.: Ecological roles and conservation challenges of social, burrowing, herbivorous mammals in the world's grasslands, Front. Ecol. Environ., 10, 477–486, https://doi.org/10.1890/110054, 2012.
Delibes-Mateos, M., Delibes, M., Ferreras, P., and Villafuerte, R.: Key role of European rabbits in the conservation of the western Mediterranean Basin hotspot, Conserv. Biol., 22, 1106–1117, https://doi.org/10.1111/j.1523-1739.2008.00993.x, 2008.
Delibes-Mateos, M., Smith, A. T., Slobodchikoff, C. N., and Swenson, J. E.: The paradox of keystone species persecuted as pests: A call for the conservation of abundant small mammals in their native range, Biol. Conserv., 144, 1335–1346, https://doi.org/10.1016/j.biocon.2011.02.012, 2011.
De Groot, R. S., Alkemade, R., Braat, L., Hein, L., and Willemen, L.: Challenges in integrating the concept of ecosystem services and values in landscape planning, management and decision making, Ecol. Complex., 7, 260–272, https://doi.org/10.1016/j.ecocom.2009.10.006, 2010.
Dial, R. and Roughgarden, J.: Theory of marine communities: The intermediate disturbance hypothesis, Ecology, 79, 1412–1424, https://doi.org/10.1890/0012-9658(1998)079[1412:TOMCTI]2.0.CO;2, 1998.
Dobson, F. S., Smith, A. T., and Gao, W. X.: Social and ecological influences on dispersal and philopatry in the plateau pika (Ochotona curzoniae), Behav. Ecol., 9, 622–635, https://doi.org/10.1093/beheco/9.6.622, 1998.
Dong, S. K., Shang, Z. H., Gao, J. X., and Boone, R. B.: Enhancing sustainability of grassland ecosystems through ecological restoration and grazing management in an era of climate change on Qinghai-Tibetan Plateau, Agr. Ecosyst. Environ., 287, 106684, https://doi.org/10.1016/j.agee.2019.106684, 2020.
Egoh, B., Drakou, E. G., Dunbar, M. B, Maes, J., and Willemen, L.: Indicators for mapping ecosystem services: a review, Ispra, Italy: European Commission, Joint Research Centre (JRC), 14–21, https://doi.org/10.2788/41823, 2012.
Eldridge, D. J. and Myers, C. A.: The impact of warrens of the European rabbit (Oryctolagus cuniculus L.) on soil and ecological processes in a semi-arid Australian woodland, J. Arid. Environ., 47, 325–337, https://doi.org/10.1006/jare.2000.0685, 2001.
Eldridge, D. J., Bowker, M. A., Maestre, F. T., Alonso, P., Mau, R. L., Papadopoulos, J., and Escudero, A.: Interactive effects of three ecosystem engineers on infiltration in a semi-arid Mediterranean grassland, Ecosystems, 13, 499–510, https://doi.org/10.1007/s10021-010-9335-4, 2010.
Fan, N., Zhou, W., Wei, W., Wang, Q., and Jiang, Y.: Rodent pest management in the Qinghai-Tibet alpine meadow ecosystem, in: Ecologically-Based Rodent Management, edited by: Singleton, G. R., Hinds, L. A., Leirs, H., and Zhang, Z., Australian Centre for International Agricultural Research, Canberra, Australia, 285–304, 1999.
Gao, J. and Carmel, Y.: Can the intermediate disturbance hypothesis explain grazing-diversity relations at a global scale?, Oikos, 129, 493–502, https://doi.org/10.1111/oik.06338, 2020.
Gong, Z.: Chinese soil taxonomy, Science Press, China, ISBN 9787030067173, 2001 (in Chinese).
Guo, Z. G., Zhou, X. R., and Hou, Y.: Effect of available burrow densities of plateau pika (Ochotona curzoniae) on soil physicochemical property of the bare land and vegetation land in the Qinghai-Tibetan Plateau, Acta Ecol. Sin., 32, 104–110, https://doi.org/10.1016/j.chnaes.2012.02.002, 2012a.
Guo, Z. G., Li, X. F., Liu, X. Y., and Zhou, X. R.: Response of alpine meadow communities to burrow density changes of plateau pika (Ochotona curzoniae) in the Qinghai-Tibet Plateau, Acta Ecol. Sin., 32, 44–49, https://doi.org/10.1016/j.chnaes.2011.12.002, 2012b.
Han, L. H., Shang, Z. H., Ren, G. H., Wang, Y. L., Ma, Y. S., Li, X. L., and Long, R. J.: The response of plants and soil on black soil patch of the Qinghai-Tibetan Plateau to variation of bare-patch areas, Acta Prataculturae Sinica, 20, 1–6, 2011.
Harris, R. B.: Rangeland degradation on the Qinghai-Tibetan plateau: A review of the evidence of its magnitude and causes, J. Arid. Environ., 74, 1–12, https://doi.org/10.1016/j.jaridenv.2009.06.014, 2010.
James, A. I., Eldridge, D. J., and Hill, B. M.: Foraging animals create fertile patches in an Australian desert shrubland, Ecography, 32, 723–732, https://doi.org/10.1111/j.1600-0587.2009.05450.x, 2009.
Jia, Q. M., Xu, R. R., Chang, S. H., Zhang, C., Liu, Y. J., Shi, W., Peng, Z. C., and Hou, F. J.: Planting practices with nutrient strategies to improves productivity of rain-fed corn and resource use efficiency in semi-arid regions, Agr. Water Manage., 228, 105879, https://doi.org/10.1016/j.agwat.2019.105879, 2020.
Leigh, J. H., Wood, D. H., Slee, A. V, and Stanger, M. G.: Effects of rabbit and kangaroogra zing on two semi-arid grassland communities in central-western new south wales, Aust. J. Bot., 37, 375–396, https://doi.org/10.1071/BT9890375, 1989.
Li, J., Zhang, F. W., Lin, L., Li, H. Q., Du, Y. G., Li, Y. K., and Cao, G. M.: Response of the plant community and soil water status to alpine Kobresia meadow degradation gradients on the Qinghai-Tibetan Plateau, China, Ecol. Res., 30, 589–596, https://doi.org/10.1007/s11284-015-1258-2, 2015.
Li, S. M. and Xie, G. D.: Spatial and temporal heterogeneity of water conservation service for meadow ecosystem, Chinese Journal of Grassland, 37, 88–93, 2015.
Liu, W., Zhang, Y., Wang, X., Zhao, J. Z., Xu, Q. M., and Zhou, L.: The relationship of the harvesting behavior of plateau pikas with the plant community, Acta Theriol. Sin., 29, 40–49, https://doi.org/10.16829/j.slxb.2009.01.007, 2009.
Liu, Y. S., Fan, J. W., Harris, W., Shao, Q. Q., Zhou, Y. C., Wang, N., and Li, Y. Z.: Effects of plateau pika (Ochotona curzoniae) on net ecosystem carbon exchange of grassland in the Three Rivers Headwaters region, Qinghai-Tibet, China, Plant. Soil., 366, 491–504, https://doi.org/10.1007/s11104-012-1442-x, 2013.
Liu, Y. S., Fan, J. W., Shi, Z. J., Yang, X. H., and Harris, W.: Relationships between plateau pika (Ochotona curzoniae) densities and biomass and biodiversity indices of alpine meadow steppe on the Qinghai-Tibet Plateau China, Ecol. Eng., 102, 509–518, https://doi.o rg/10.1016/j.ecoleng.2017.02.026, 2017.
Lu, X. Y., Kelsey, K. C., Yan, Y., Sun, J., Wang, X. D., Cheng, G. W., and Neff, J. C.: Effects of grazing on ecosystem structure and function of alpine grasslands in Qinghai-Tibetan Plateau: A synthesis, Ecosphere, 8, e01656, https://doi.org/10.1002/ecs2.1656, 2017.
Martínez-Estévez, L., Balvanera, P., Pacheco, J., and Ceballos, G.: Prairie dog decline reduces the supply of ecosystem services and leads to desertification of semiarid grasslands, PLoS One, 8, e75229, https://doi.org/10.1371/journal.pone.0075229, 2013.
Millennium Ecosystem Assessment.: Ecosystems and human well-being: Synthesis, Island Press, Washington, DC, ISBN 9241563095, 2005.
Nelson, D. W. and Sommers, L. E.: Total carbon, organic carbon, and organic matter. Methods of soil analysis, in: Part 3-Chemical and Microbiological Properties, 539–579, https://doi.org/10.2134/agronmonogr9.2.2ed.c29, 1982.
Norton, L. R., Inwood, H., Crowe, A., and Baker, A.: Trialling a method to quantify the “cultural services” of the English landscape using Countryside Survey data, Land Use Policy, 29, 449–455, https://doi.org/10.1016/j.landusepol.2011.09.002, 2012.
Pang, X. P. and Guo, Z. G.: Plateau pika disturbances alter plant productivity and soil nutrients in alpine meadows of the Qinghai-Tibetan Plateau, China, Rangeland J., 39, 133–144, https://doi.org/10.1071/RJ16093, 2017.
Pang, X. P. and Guo, Z. G.: Effects of plateau pika disturbance levels on the plant diversity and biomass of an alpine meadow, Grassland Sci., 64, 159–166, https://doi.org/10.1111/grs.12199, 2018.
Pang, X. P., Yu, C. Q., Zhang, J., Wang, Q., Guo, Z. G., and Tian, Y.: Effect of disturbance by plateau pika on soil nitrogen stocks in alpine meadows, Geoderma, 372, 114392, https://doi.org/10.1016/j.geoderma.2020.114392, 2020a.
Pang, X. P., Wang, Q., Zhang, J., Xu, H. P., Zhang, W. N., Wang, J., and Guo, Z. G.: Responses of soil inorganic and organic carbon stocks of alpine meadows to the disturbance by plateau pikas, Eur. J. Soil Sci., 71, 706–715, https://doi.org/10.1111/ejss.12895, 2020b.
Pang, X. P., Yang, H., Wei, X. X., and Guo, Z. G.: Effect of plateau pika (Ochotona curzoniae) bioturbation on soil C-N-P stoichiometry in alpine meadows, Geoderma, 397, 115098, https://doi.org/10.1016/j.geoderma.2021.115098, 2021a.
Pang, X. P., Wang, Q., and Guo, Z. G.: The impact of the plateau pika on the relationship between plant aboveground biomass and plant species richness, Land. Degrad. Dev., 32, 1205–1212, https://doi.org/10.1002/ldr.3790, 2021b.
Qu, J. P., Li, W. J., Yang, M., Ji, W. H., and Zhang, Y. M.: Life history of the plateau pika (Ochotona curzoniae) in alpine meadows of the Tibetan Plateau, Mamm. Biol., 78, 68–72, https://doi.org/10.1016/j.mambio.2012.09.005, 2013.
Sierra-Corona, R., Davidson, A., Fredrickson, E. L., Luna-Soria, H., Suzan-Azpiri, H., Ponce-Guevara, E., and Ceballos, G.: Black-tailed prairie dogs, cattle, and the conservation of North America's Arid Grasslands, PLoS One, 10, e0118602, https://doi.org/10.1371/journal.pone.0118602, 2015.
Smith, A. T. and Wang, X. G.: Social relationships of adult black-lipped pikas (Ochotona curzoniae), J. Mammal., 72, 231–247, https://doi.org/10.2307/1382094, 1991.
Smith, A. T. and Foggin, J. M.: The plateau pika (Ochotona curzoniae) is a keystone species for biodiversity on the Tibetan plateau, Anim. Conserv., 2, 235–240, https://doi.org/10.1017/S1367943099000566, 1999.
Strömberg, C. A. E. and Staver, A. C.: The history and challenge of grassy biomes, Science, 377, 592–593, https://doi.org/10.1126/science.add1347, 2022.
Sun, F. D., Chen, W. Y., Liu, L., Liu, W., Cai, Y. M., and Smith, P.: Effects of plateau pika activities on seasonal plant biomass and soil properties in the alpine meadow ecosystems of the Tibetan Plateau, Grassland Sci., 61, 195–203, https://doi.org/10.1111/grs.12101, 2015.
Tang, Y. K., Wu, Y. T., Wu, K., Guo, Z. W., Liang, C. Z., Wang, M. J., and Chang, P. J.: Changes in trade-offs of grassland ecosystem services and functions under different grazing intensities, Chin. J. Plant Ecol., 43, 408–417, https://doi.org/10.17521/cjpe.2018.0289, 2019.
Van Eekeren, N., de Boer, H., Hanegraaf, M., Bokhorst, J., Nierop, D., Bloem, J., Schouten, T., de Goede, R., and Brussaard, L.: Ecosystem services in grassland associated with biotic and abiotic soil parameters, Soil. Biol. Biochem., 42, 1491–1504, https://doi.org/10.1016/j.soilbio.2010.05.016, 2010.
Wang, C. T., Wang, G. X., Liu, W., Wang, Q. L., and Xiang, Z. Y.: Vegetation roots and soil physical and chemical characteristics in degeneration succession of the Kobresia pygmaea meadow, Ecol. Environ. Sci., 21, 409–416, https://doi.org/10.16258/j.cnki.1674-5906.2012.03.002, 2012.
Wang, Q., Guo, Z. G., Pang, X. P., Zhang, J., and Yang, H.: Effects of small-herbivore disturbance on the clonal growth of two perennial graminoids in alpine meadows, Alpine. Bot., 130, 115–127, https://doi.org/10.1007/s00035-020-00240-9, 2020.
Wen, L., Dong, S. K., Li, Y. Y., Li, X. Y., Shi, J. J., Wang, Y. L., Liu, D. M., and Ma, Y. S.: Effect of degradation intensity on grassland ecosystem services in the alpine region of Qinghai-Tibetan Plateau, China, PLoS One, 8, e58432, https://doi.org/10.1371/journal.pone.0058432, 2013.
Willott, S. J., Miller, A. J., Incoll, L. D., and Compton, S. G.: The contribution of rabbits (Oryctolagus cuniculus) to soil fertility semi-arid, Biol. Fert. Soils., 31, 379–384, https://doi.org/10.1007/s003749900183, 2000.
Yang, D., Pang, X. P., Jia, Z. F., and Guo, Z. G.: Effect of plateau zokor on soil carbon and nitrogen concentrations of alpine meadows, CATENA, 207, 105625, https://doi.org/10.1016/j.catena.2021.105625, 2021.
Yu, C., Pang, X. P., Wang, Q., Jin, S. H., Shu, C. C., and Guo, Z. G.: Soil nutrient changes induced by the presence and intensity of plateau pika (Ochotona curzoniae) disturbances in the Qinghai-Tibet Plateau, China, Ecol. Eng., 106, 1–9, https://doi.org/10.1016/j.ecoleng.2017.05.029, 2017a.
Yu, C., Zhang, J., Pang, X. P., Wang, Q., Zhou, Y. P., and Guo, Z. G.: Soil disturbance and disturbance intensity: Response of soil nutrient concentrations of alpine meadow to plateau pika bioturbation in the Qinghai-Tibetan Plateau, China, Geoderma, 307, 98–106, https://doi.org/10.1016/j.geoderma.2017.07.041, 2017b.
Zhang, Y., Dong, S. K., Gao, Q. Z., Liu, S. L., Liang, Y., and Cao, X. J.: Responses of alpine vegetation and soils to the disturbance of plateau pika (Ochotona curzoniae) at burrow level on the Qinghai-Tibetan Plateau of China, Ecol. Eng., 88, 232–236, https://doi.org/10.1016/j.ecoleng.2015.12.034, 2016.
Zhang, H. Y., Fan, J. W., Cao, W., Zhong, H. P., Harris, W., Gong, G. L., and Zhang, Y. X.: Changes in multiple ecosystem services between 2000 and 2013 and their driving factors in the Grazing Withdrawal Program, China, Ecol. Eng., 116, 67–79, https://doi.org/10.1016/j.ecoleng.2018.02.028, 2018.
Zhang, W. N., Wang, Q., Zhang, J., Pang, X. P., Xu, H. P., Wang, J., and Guo, Z. G.: Clipping by plateau pikas and impacts to plant community, Rangeland, Ecol. Manag., 73, 368–374, https://doi.org/10.1016/j.rama.2020.01.010, 2020.
Zhao, G. Q., Li, G. Y., Ma, W. H., Zhao, D. Z., and Li, X. Y.: Impacts of Ochotona pallasi disturbance on alpine grassland community characteristics, Chin. J. Appl. Ecol., 24, 2122–2128, https://doi.org/10.13287/j.1001-9332.2013.0398, 2013.