the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 06 Apr 2018
| 06 Apr 2018
Impact of elevated precipitation, nitrogen deposition and warming on soil respiration in a temperate desert
Ping Yue
Xiaoqing Cui
Yanming Gong
Kaihui Li
Keith Goulding
Xuejun Liu
Soil respiration (Rs) is the most important source of carbon dioxide emissions from soil to atmosphere. However, it is unclear what the interactive response of Rs would be to environmental changes such as elevated precipitation, nitrogen (N) deposition and warming, especially in unique temperate desert ecosystems. To investigate this an in situ field experiment was conducted in the Gurbantunggut Desert, northwest China, from September 2014 to October 2016. The results showed that precipitation and N deposition significantly increased Rs, but warming decreased Rs, except in extreme precipitation events, which was mainly through its impact on the variation of soil moisture at 5 cm depth. In addition, the interactive response of Rs to combinations of the factors was much less than that of any single-factor, and the main response was a positive effect, except for the response from the interaction of increased precipitation and high N deposition (60 kg N ha−1 yr−1). Although Rs was found to show a unimodal change pattern with the variation of soil moisture, soil temperature and soil NH-N content, and it was significantly positively correlated to soil dissolved organic carbon (DOC) and pH, a structural equation model found that soil temperature was the most important controlling factor. Those results indicated that Rs was mainly interactively controlled by the soil multi-environmental factors and soil nutrients, and was very sensitive to elevated precipitation, N deposition and warming. However, the interactions of multiple factors largely reduced between-year variation of Rs more than any single-factor, suggesting that the carbon cycle in temperate deserts could be profoundly influenced by positive carbon–climate feedback.
- Article
(4086 KB) - Full-text XML
-
Supplement
(483 KB) - BibTeX
- EndNote





Global climate warming, changes in precipitation patterns and increased atmospheric nitrogen (N) deposition have all occurred since the Industrial Revolution, especially in temperate regions (IPCC, 2013), which will be expected to significantly change soil respiration (Rs), the most important source of carbon dioxide (CO2) from soil to atmosphere (Wu et al., 2011): the annual CO2 flux from Rs was 10-fold that of fossil fuel emissions (Eswaran et al., 1993; Batjes, 1996; Gougoulias et al., 2014). Therefore, even a small change in Rs will profoundly affect greenhouse gas balance and climate (Heimann and Reichstein, 2008). Although a number of experiments of the effects of warming, precipitation and N deposition on Rs have been conducted in alpine grasslands, tundra regions, peatlands and temperate forests (Lafleur and Humphreys, 2008; Strong et al., 2017; Yang et al., 2017; Zhao et al., 2017), studies in temperate desert ecosystems are scarce, especially those on the impact of these changes' interactions on Rs. A field study of multi-factor interactive effects on Rs was therefore conducted in a temperate desert ecosystem to help in understanding the response of Rs to climate change and N deposition in future and highlight the main driving factors.
Rs includes autotrophic respiration (RA), which is mainly from plant roots and mycorrhizal activities, and heterotrophic respiration (RH), which is mainly from the activities of microorganisms (Hanson et al., 2000). Soil moisture is a critical limiting factor for plant roots and microbial activities in desert ecosystems (Huang et al., 2015a): Rs significantly increased, by 47–70 %, in a degraded steppe in Inner Mongolia, China, due to increasing precipitation (Chen et al., 2013), with the effect especially strong in summer (Zhang et al., 2017). In addition, in arid ecosystems, increasing precipitation significantly stimulated plant growth, enhanced soil microbial activity and abundance (Huang et al., 2015a), and it changed soil nutrient and substrate concentration, such as dissolved organic carbon (DOC), inorganic nitrogen content, moisture and temperature (Huang et al., 2015b).
Warming significantly increased soil temperature, another important controlling factor for plant growth and microbial activity (Sheik et al., 2011; Huang et al., 2015a). Rs rates were significantly increased in a forest soil and Tibetan Plateau grassland by warming (Chen et al., 2017a), reducing Rs with decreasing soil moisture in the growing season, but increasing Rs in the non-growing season (Fang et al., 2017; Li et al., 2017); no significant impact was observed from warming (T. Liu et al., 2016). Therefore, how Rs is affected by warming induced variations in the soil environment is still unclear. In addition, low and short-term N deposition enhanced Rs, while higher and long-term N deposition inhibited Rs due to changes in plant growth and microbial activity (Zhu et al., 2017), but no impacts have also been reported (Luo et al., 2017; Zhang et al., 2017). A meta-analysis showed that the effects of N enrichment on soil CO2 fluxes depended on temperature and soil properties (Zhong et al., 2016); desert soils may be even more sensitive to its variation.
A nation-wide analysis showed that warming, elevated N deposition and precipitation significantly increased Rs in China (Feng et al., 2017). Some studies have shown that the warming effect on Rs mainly depended on the variation of soil moisture in a dry forest soil (Li et al., 2017). Luo et al. (2008), using a modeling analysis, found that interactive effects became increasingly weaker with increasing intensity of the factors, but a recent meta-analysis showed that interactive effects were much greater than an individual factor (L. Y. Zhou et al., 2016). Thus how multi-factor interactions impact Rs is still unclear. Therefore, an in situ experiment was carried out in the Gurbantunggut Desert to (1) investigate the single-factor and interactive responses of Rs to warming, precipitation and N deposition and (2) identify the main controlling factors on Rs.
2.1 Study site
A field experiment was carried out at the southern edge of the Gurbantunggut Desert (44∘26′ N, 87∘54′ E; 436.8 m a.s.l.), northwest China, from September 2014 to October 2016. This is the largest fixed/semi-fixed temperate desert in China. The mean annual temperature and precipitation are 7.1 ∘C and 215.6 mm, respectively (Cui et al., 2017), and annual potential evaporation exceeds 2000 mm. From late November to mid-March, 20–35 cm of snow covers the whole desert (equivalent to 38–64 mm rainfall; Huang et al., 2015c). The growing season is from April to October. This desert soil is of extremely low fertility and is highly alkaline (Cui et al., 2017). Soil organic carbon, total N content, soil NO-N, NH-N contents and C : N ratio are 2.21 ± 0.71 g kg−1, 0.08 ± 0.003 g kg−1, 4.49 ± 0.71 mg kg−1, 1.38 ± 0.74 mg kg−1 and 21.39 ± 1.84, respectively (Table 1; Cui et al., 2017). Plant species are dominated by Haloxylon ammodendron and Haloxylon persicum, and vegetation is extremely sparse, with only 30 % coverage, with some spring ephemeral plants (May–June), annuals and perennial herbaceous plants (July–August; R. Liu et al., 2016). Spring ephemerals account for > 60 % of the community cover and 85 % of the biomass. Summer ephemerals, annuals and perennials usually account for only a small proportion of the community biomass before June, but dominate the community after the die-back of the spring annuals (Huang et al., 2015c).
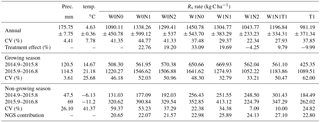
Table 1The annual, growing season (GS), non-growing season (NGS) and between-year fluxes and variation of soil respiration (Rs) over September 2014 to September 2016 (mean ± SE), including the contribution of GS and NGS, and the treatment effect. The positive values stand for increased Rs, and the negative value stand for reduced Rs. Dates are shown in YYYY.M format.
2.2 Experimental treatments
A striking N deposition rate (35.2 kg N ha−1 yr−1) exists in the Gurbantunggut Desert due to the rapid development of agriculture and industry, with the main form being ammonium nitrate (NH4NO3); deposition is split between wet (19.6 kg N ha−1 yr−1) and dry (15.6 kg N ha−1 yr−1; Song et al., 2015). In addition, according to the forecast of Galloway et al. (2008), atmospheric N deposition will double from the early 1990s to 2050; Liu et al. (2010) predict that precipitation in this region will increased by 30 % in next 30 years. In September 2014 to August 2016, an in situ complete block interactive experiment was therefore conducted to study the impact of N deposition and increased precipitation on Rs. The three levels of N deposition (0 kg N ha−1 yr−1, control, N0; 30 kg N ha−1 yr−1, low, N1; 60 kg N ha−1 yr−1, high, N2) and two levels of precipitation (“natural” precipitation, W0, and an increase of 30 % – an extra 60 mm precipitation annually, W1) were applied (Cui et al., 2017). Therefore there were six treatments (W0N0, W0N1, W0N2, W1N0, W1N1 and W1N2) with four replicates of each treatment. Each replicate plot was 10 m × 10 m with a 5 m wide buffer zone. The additional precipitation and N deposition (NH4NO3) were added 12 times in April, July and September, equivalent to 5 mm precipitation and 2.5 or 5 kg N ha−1 per application over a week. The NH4NO3 was diluted in 50 L water (equal to 0.5 mm precipitation), and evenly applied following the simulated precipitation. The same amount of water was applied to the control plots (W0N0).
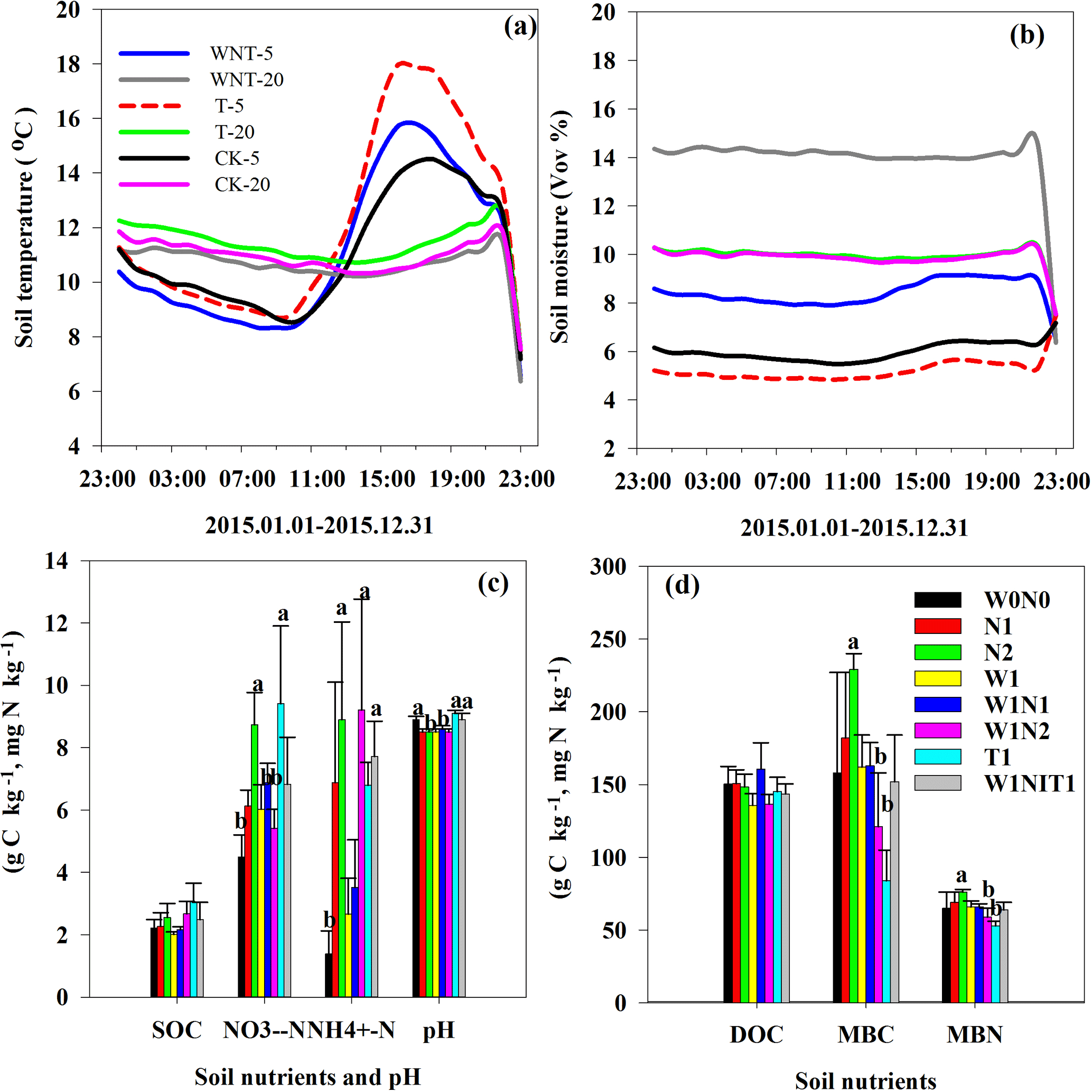
Figure 1Comparative effects of warming from open-top chambers (OTCs), increased precipitation and N deposition on soil temperature (a), soil moisture (b) at 5 and 20 cm depth; soil organic carbon (SOC), NH-N content, NO-N content and pH (c), dissolved organic carbon (DOC), microbial biomass carbon (MBC) and microbial biomass nitrogen (MBN, d). The data are mean ± SE, n = 4 in (c) and (d); different letters indicate significant effect at P < 0.05.
Rapid warming (0.6 ∘C per decade), increasing precipitation (3–5 mm yr−1 since 1979) and high N deposition (3 kg N ha−1 since 1980) are affecting the Gurbantunggut Desert (Liu et al., 2013; Li et al., 2015), which are excepted to affect rate of Rs. Therefore, another interactive experiment was established at the same time, simulating the three most likely climate scenarios in the future: (1) warming only (W0N0T1), (2) increased precipitation and N deposition without warming (W1N1T0), (3) the interaction of increasing precipitation, N deposition and warming (W1N1T1); all compared with the current climate (W0N0T0). Therefore, there were four treatments (W0N0T1, W1N1T0, W1N1T1, W0N0T0) with four replicates (plots) of each treatment. Open-top chambers (OTCs) were used to simulate warming. The OTCs were designed with 5 mm transparent tempered glass and stainless steel angle iron to the ITEX standard (Marion et al., 1997). They were 2 m high and 4 m in diameter, with each OTC area being 12 m2. However, the design was improved such that the top and bottom OTC areas were the same so that precipitation and snowfall were the same as that in the surrounding environment; this also avoids overheating inside the OTCs. The timings of water and N applications were the same as in the complete block interactive experiment.
2.3 Measurements
Rs in all plots were measured twice or thrice a week (continuous measurements over 3 days were made following simulated precipitation and N deposition) using gas chromatography and static chambers (50 cm × 50 cm× 10 cm) at locations where only spring ephemeral plants grow, without any annuals and perennials, in order to minimize the between-treatment spatial heterogeneity due to sparse annuals and perennials (Liu et al., 2012). Gas samples were collected between 10:00 and 12:00 (GMT + 8) throughout the experimental period; measurements from this period were close to the diurnal averages (Figs. 3b, d and S1 in the Supplement). Gas samples were collected from the headspace of each chamber 0, 10, 20 and 30 min after closing the chamber. The gas samples were analyzed within 3 days using a gas chromatograph (GC; Agilent 7890A, Agilent Technologies, Santa Clara, CA, USA) equipped with a flame ionization detector for quantitative Rs (Liu et al., 2012). Rs rates were calculated from four concentrations of the gas samples based on a first-order differential linear or nonlinear equation and were temperature and pressure corrected (Liu et al., 2012; Zhang et al., 2014). Soil samples were taken monthly from around the static chambers to a depth of 10 cm using an auger (3.5 cm in diameter). Fine roots and small stones were separated out using a 2 mm sieve. Dissolved organic carbon (DOC) was extracted with deionized water (soil : water ratio = 1 : 10) by a centrifuge at 10000 rpm for 5 min and analyzed using a TOC analyzer (multi N/C 3100, Jena, Germany; Jones and Willett, 2006). The chloroform fumigation extraction of Brookes (1985) was used to measure microbial biomass carbon (MBC) and microbial biomass nitrogen (MBN). Soil organic carbon (SOC) was measured using the potassium dichromate method (Jiang et al., 2014), and soil NON and NH-N was analyzed as per Yue et al. (2016). Soil pH was measured with a 1 : 5 soil : deionized water suspension using a pH meter (Seven Easy, Mettler-Toledo, Switzerland). Caipos Soil and Environment Monitoring Systems (Caipos GmbH, Austria) were used to monitor soil moisture/temperature at 5 and 20 cm depth every hour.
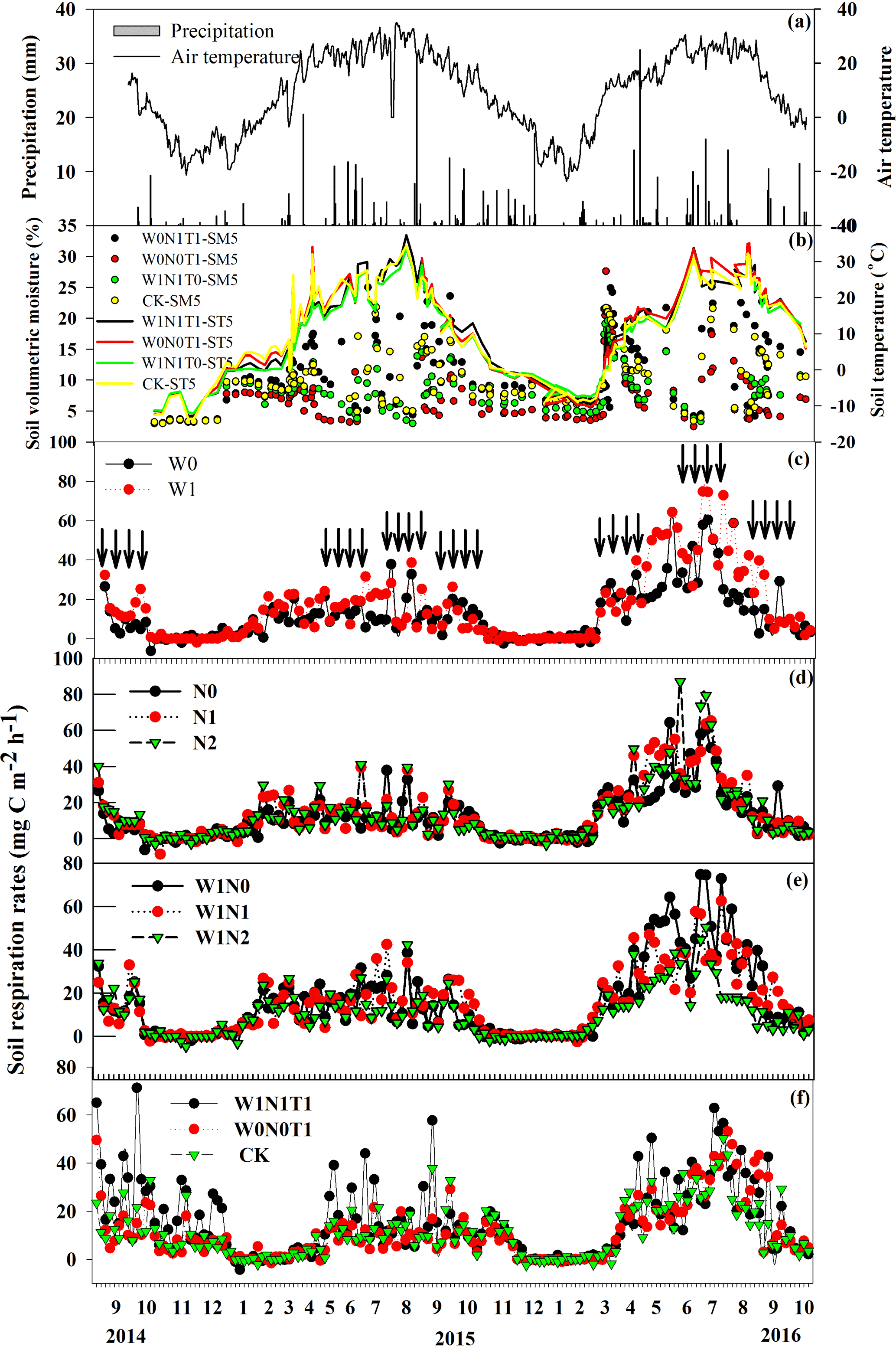
Figure 2Variation in rainfall (mm, a), and air temperature (∘C, a) from September 2014 to October 2016 at the Gurbantunggut Desert. Also shown are soil moisture and temperature responses to increasing precipitation and warming (b), and the response of Rs (mean, n = 4) to precipitation (c), N deposition (d, e) and warming (f). W0 and W1 indicate under ambient precipitation (without water addition) and 60 mm yr−1 precipitation addition; N0, N1 and N2 indicate 0, 30 and 60 kg N ha−1 yr−1 nitrogen addition; W1N0, W1N1 and W1N2 indicate 0, 30 and 60 kg N ha−1 yr−1 nitrogen addition under 60 mm yr−1 precipitation addition; W1N1T1, W0N0T1 and W0N0T0 indicate the interaction between increasing precipitation (60 mm yr−1), N deposition (30 kg N ha−1 yr−1) and warming by OTCs, warming alone (without increasing precipitation and N deposition) and control plots, respectively. Black arrows indicate simulated precipitation (5 mm) and N deposition (0.25 or 0.5 g N m−2). Each point represents the mean of four replications (chambers). Standard deviations for Rs are not shown for figure clarity.
2.4 Effects of each treatment on Rs
Each treatment effect was analyzed using the following formula to better evaluating the effect of precipitation, warming and N deposition on Rs (Yue et al., 2016).
where the treatment effect is W0N1, W0N2, W1N0, W1N1, W1N2, W1N1T1 or W0N0T1 effect on Rs (a positive value shows that the treatment has increased Rs and a negative value shows decrease of Rs), corresponding RT,s represents Rs from the W0N1, W0N2, W1N0, W1N1, W1N2, W1N1T1 or W0N0T1 plots (mg C m−2 h−1) and RC,s indicates the Rs from the control plots (W0N0, mg C m−2 h−1).
2.5 Statistical analyses
Treatment effect on SOC, NO-N, NH-N content, pH, DOC, MBC and MBN were examined in each treatment by least significant difference (LSD; p < 0.05). The single-factor and interaction effects of precipitation, warming and N deposition on Rs were detected using multi-way analysis of variance (ANOVA), and the cumulative effect of precipitation, warming and N deposition on Rs were tested by repeated measures ANOVA. In addition, the relationships of Rs and DOC, MBC, MBN, soil temperature, soil moisture, NH-N content, soil NO-N and pH were described using a linear or nonlinear regression model. The factors of key controls on Rs were analyzed with structural equation models (SEMs). SPSS software (version 20.0) was used to conduct all statistical analyses, and statistically significant differences were set at P < 0.05. All figures were created using the Sigmaplot software package (version 10.0), but SEM analyses were carried out using AMOS 22.0 (Amos Development Corporation, Chicago, IL, USA).
3.1 Treatment effects on soil environmental and properties
Soil temperatures at 5 cm depth mostly increased between 11:00 and 22:00 every day due to warming; the average annual soil temperatures at 5 and 20 cm depth increased significantly, by 4.41 and 3.67 ∘C, respectively (Fig. 1a). Soil moisture at 5 cm depth was decreased by warming by only 0.61 v∕v % (Fig. 1b), and a very small decrease of 0.01 v∕v % in soil moisture at 20 cm depth was observed (Fig. 1b). Soil moisture at 5 and 20 cm depth were largely increased by the increased precipitation (Fig. 1b). N deposition and warming significant increased soil NH-N and NO-N contents (Fig. 1c), but no significant change was found from increased precipitation. Soil MBC and MBN were greatly increased by N deposition, but significant negative effects on soil MBC and MBN were observed by warming and the interaction of precipitation and N deposition (Fig. 1d). No significant change in SOC and DOC was observed in any treatment (Fig. 1c and d).
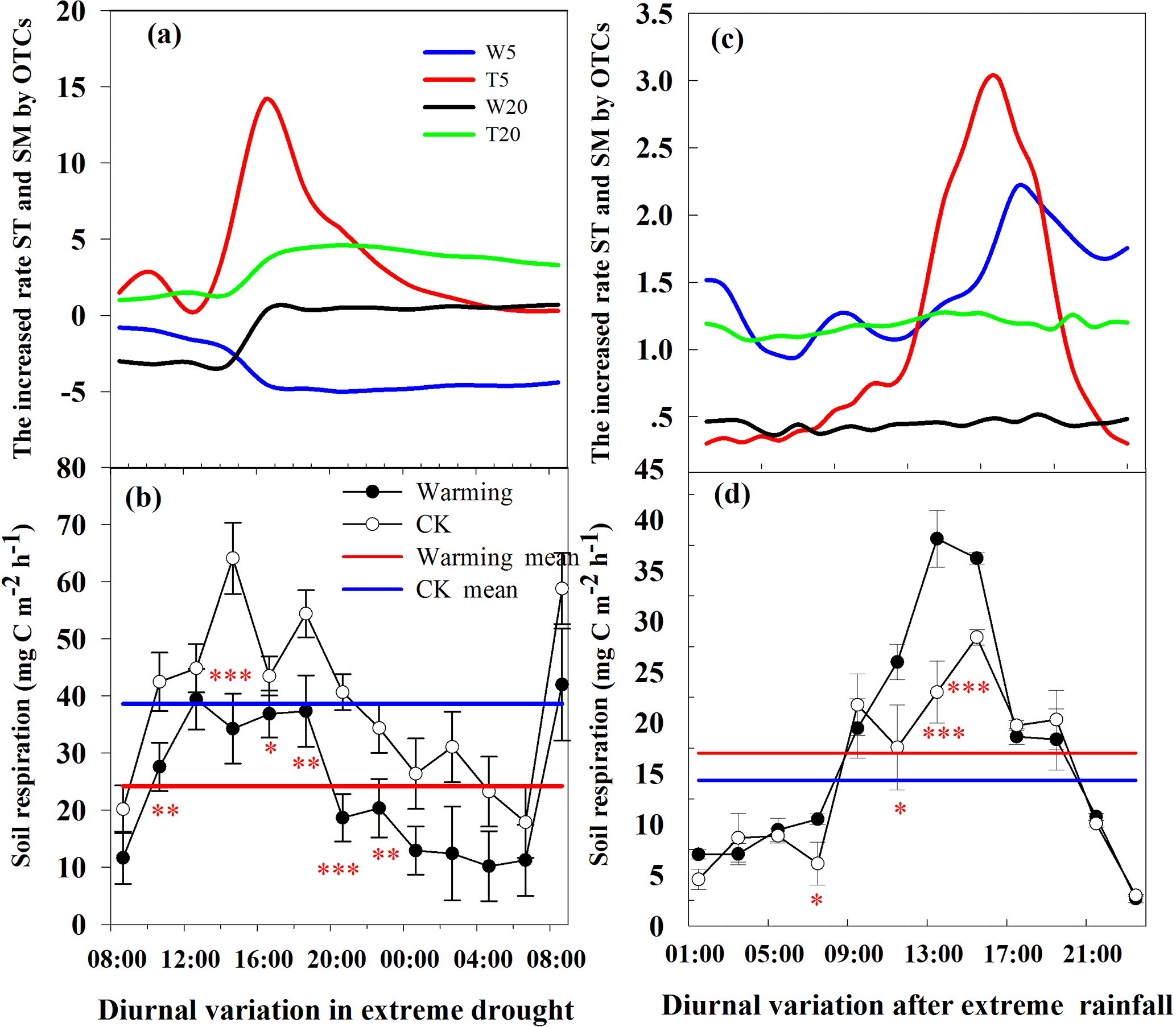
Figure 3Post-extreme-rainfall diurnal variation in Rs (mean ± SE, n = 4, d) with variation in soil temperature and soil moisture (c), and extreme-drought variation in Rs (mean ± SE, n = 4, b) with variation in soil temperature (T5, T20, a) and soil moisture at 5 cm (W5) or 20 cm (W20) depth caused by warming in OTCs. Positive values indicate an increase due to warming, and negative values indicate decline. A red straight line indicates the average value of Rs inside the OTCs in (b) and (d), and a blue straight line represents the average value of Rs out of OTCs in (b) and (d). Red *, ** and *** symbols indicate significant effects at P < 0.05, P < 0.01 and P <0.001, respectively.
3.2 Precipitation, warming and N deposition effects on Rs
In our study, a weak Rs emission rate (−2.46 to 50.26 mg C m−2 h−1) was observed at control plots with an average emission rate of 12.18 mg C m−2 h−1 from September 2014 to October 2016 (Fig. 2c). Annual cumulative rate of Rs was 1090.11 ± 450.78 kg C ha−1, with the non-growing season accounting for 20.7 % of the annual emission (Table 1). Rs was significantly enhanced by increasing 5 mm precipitation and N deposition from 12.18 to 16.23 and to 14.97 mg C m−2 h−1(average), respectively (P < 0.001; Fig. 2c and d; Table 2), with annual Rs increased by 33.1 % and 19.2–22.8 %, respectively (Table 1). The low N deposition effect on Rs was much higher than that of high N deposition (Fig. 2c and d). However, Rs was reduced mostly by warming, although not significant (P=0.084; Table 2). The high temperatures and low humidity at times of peak sunshine during the diurnal variation significantly inhibited the emission rate (Figs. 3a and b, S2), but it was also significantly increased by warming following extreme rainfall events that increased soil moisture (Fig. 3c and d). The diurnal trend in Rs was consistent with that of soil temperature at 5 cm depth (Fig. 3). In addition, the interactive responses of Rs to increasing precipitation, warming and N deposition were much lower than that from any single-factor (Table 1), and with the interaction of 60 kg ha−1 N and extra precipitation decreasing Rs by 4.25 % (Table 1). Overall, annual Rs rates were significantly impacted by precipitation, N deposition, and their interaction (Table 2), but no significant net change was caused by warming (Table 2), although Rs rates were decreased by 9.99 % (Table 1).
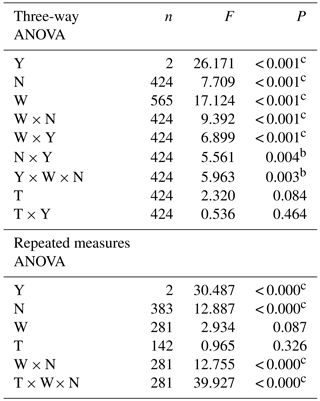
Table 2Tests of significance of year (Y), warming (T), precipitation (W) and nitrogen addition (N) on soil respiration (Rs) using multivariate ANOVA (F and P values). The cumulative effect of precipitation, N deposition and warming on Rs in 2015 and 2016 (F and P values) as assessed by repeated measures ANOVA. Superscripts a, b and c indicate significant effects at P < 0.05, 0.01 and 0.001, respectively.
3.3 Temporal variation and its control
The results of repeated measures ANOVA showed that significant cumulative effects on Rs were found to be due to N deposition and interaction between N deposition and precipitation or warming rather than increasing precipitation and warming alone (Table 2). A large between-year variation in Rs was observed with a coefficient of variation (CV) up to 41.4 % (a much higher Rs rate was observed in 2016 than 2015), but variation was reduced by increasing precipitation, N deposition and warming and their interaction, except with an increase in N deposition of 30 kg ha−1 (Table 1). The results of regression analysis showed that Rs was significantly increased by increases in pH and DOC (Fig. 4e and f), but no significant relationships were found with MBC, MBN or NO-N content (Fig. 4a, b and c). In addition, different response characteristics of Rs in the impacts of increased soil moisture, soil temperature and NH-N content were found. Soil moisture was the most important controlling factor when it was < 4.2 % and soil temperature was > 26.5 ∘C (Figs. 4g, h and S2). Secondly, soil temperature was the most important limiting factor when soil moisture was > 15.9 % and soil temperature < 2.7 ∘C (Figs. 4g, h and S2). Thirdly, there was no significant impact on Rs when soil NH-N content was < 6.3 mg N kg−1. A significant increase in Rs occurred when soil NH-N content was between 6.3 and 12.6 mg N kg−1, but Rs was inhibited when soil NH-N content was between 12.6 and 31.6 mg N kg−1 (Fig. 4d).
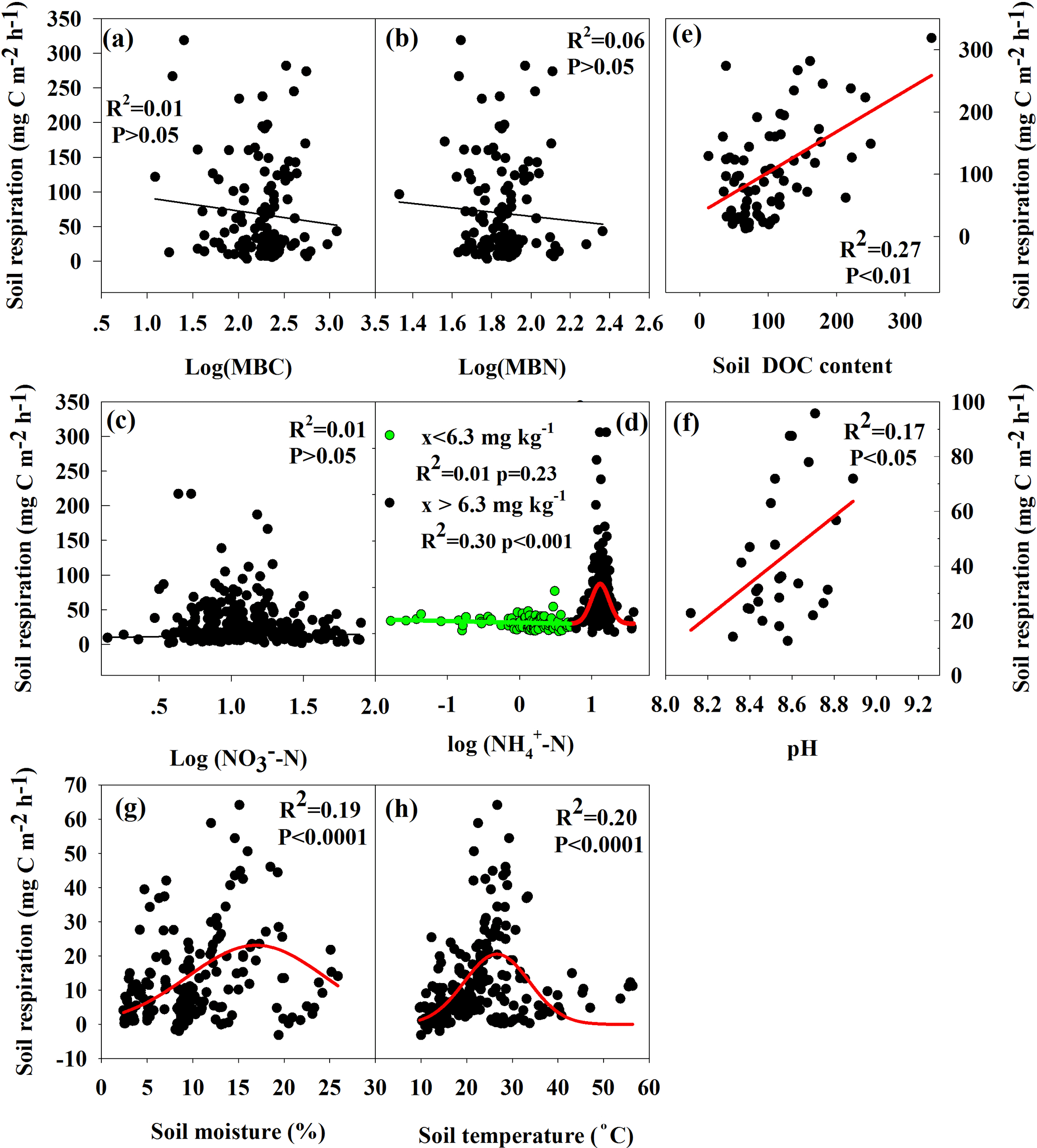
Figure 4The relationship of soil respiration with microbial biomass carbon (MBC, a); microbial biomass carbon (MBN, b); soil NO3-N (c); NH-N content (x, d); soil dissolved organic carbon (DOC, e); pH (f); soil moisture (g) and soil temperature (h).
4.1 Single-factor impacts of precipitation, N deposition and warming on Rs
Annual Rs was 1090 kg C ha−1 in this temperate desert, with the non-growing season accounting for 20.7 % of the annual flux (Table 1). This is consistent with previous studies here (Zhou et al., 2014; Huang et al., 2015a) because SOC content was very low (Fig. 1c), and vegetation was very sparse in this desert (R. Liu et al., 2016). Increasing precipitation significantly increased Rs (Fig. 2c). It is also consistent with the results of a meta-analysis and previous studies here (Huang et al., 2015a ; L. L. Liu et al., 2016). This is because the growth of desert plants and microbial activity are significantly activated by increasing precipitation (Huang et al., 2015a), and microbial biomass, mass-specific respiration, microbial biomass carbon (MBC) and nitrogen (MBN), and microbial PLFAs (phospholipid fatty acids) were consistently significantly enhanced by increased precipitation (Zhang et al., 2013; Huang et al., 2015a). However, Rs in our study was much higher in moderate soil moisture conditions than with too little or too much soil moisture (Fig. 4g). This suggests that Rs is mainly RH rather than RA in this desert, namely from soil microorganism, because (1) too little or too much soil moisture could significantly inhibit microbial activity due to variation of soil temperature and soil properties (Ma et al., 2013), while moderate soil moisture could significantly enhance microbial activity (Skopp et al., 1990) and (2) the biomass of fine roots was not significantly enhanced at our sites by increased precipitation (Cui et al., 2017). This is consistent with results from a desert steppe in northern China where the contribution of RH (78.1 %) was significantly higher than that of RA (21.9 %) under increasing precipitation (T. Liu et al., 2016).
N deposition also significantly increased Rs, especially in low N deposition (Fig. 2d). This is consistent with results from an alpine meadow and in the Loess Plateau (Fang et al., 2017; Zong et al., 2017), and with a meta-analysis showing that N deposition increased Rs by 8.8 % (L. Y. Zhou et al., 2016). This is because N deposition, on the one hand, could increase fine root biomass, although this was not significant in our study (Cui et al., 2017), and on the other hand, increases microbial activity and abundance by low N deposition (Huang et al., 2015b). However, this was inconsistent with a young Cunninghamia lanceolata forest (Wang et al., 2017), and beneath shrubs of H. ammodendron, soil high N content had the opposite effect as in our study site (Chen et al., 2013; Huang et al., 2015b). Additionally, the results of nonlinear regression analysis showed that higher Rs rates occurred at moderate soil NH-N contents (between 6.3 and 12.6 mg N kg−1), while lower Rs occurred in much lower (< 6.3 mg N kg−1) or much higher ( > 12.6 mg N kg−1) soil NH-N contents (Fig. 4d); however, this effect of N deposition on Rs is not consistent with other ecosystems (Burton et al., 2004; Chen et al., 2013; Liu et al., 2015; Chen et al., 2017b). This is because the desert soil is extremely limited compared to other ecosystem (Adams, 2003), so low N deposition enhanced plant growth and microbial activity, but high N inhibited microbial activity and community composition, and reduced Rs (Zhou et al., 2014; Huang et al., 2015b). Overall, soil NH-N content was an important controlling factor for Rs because microbial activity, abundance and species diversity were regulated by soil NH-N content in this desert, and Rs was very sensitive to variation of N deposition.
Warming decreased Rs (Fig. 2f), although not significantly (P=0.084; Table 2), which was consistent with results from a semi-arid alfalfa pasture of the Loess Plateau (Fang et al., 2017). In addition, significant decreases in Rs were observed on extreme drought or hot sunny days when soil moisture was reduced, and sharply reduced Rs occurred when soil temperature reached 37 ∘C (Figs. 3a, b and S1). This is because (i) microbial activity is significantly inhibited by extreme temperatures and low soil moisture may reduce population size by 50–80 % (Sheik et al., 2011) and (ii) fine root growth is inhibited in high temperature and low soil moisture. Others have noted this phenomenon as occurring at about 16:00 each day (Ma et al., 2013), but in our study the effect was advanced to 14:00 by warming, which may reduce carbon emission from soil to atmosphere. However, this is not consistent with results from a tundra ecosystem, subtropical forest or alpine regions where Rs was significantly increased by warming due to the limitation of soil temperature in these ecosystems, and not to significant change in soil moisture (Noh et al., 2016; Wu et al., 2016; Y. M. Zhou et al., 2016). In addition, a significant increase in Rs was found following enhanced precipitation with warming (Fig. 3c and d), which indicates that soil moisture was the most important controlling factor for Rs under a warming climate. This is consistent with other studies (Chen et al., 2017a; Zhao et al., 2017). However, statistical analysis showed that overall no significant impact on Rs was found during the experimental period due to warming (it was reduced by 9.99 %). This is because our gas samples were taken at 10:00–12:00 each day, when average soil temperatures were increased by about one degree. Thus mean annual Rs was not sensitive to temperature changes this small in contrast to the very significant effects of short-term diurnal changes in soil temperature observed between 12:00 and 17:00 (Fig. 3a and c). However, gas samples during 10:00–12:00 in this study was able to catch the mean warming effects on Rs, except in some extreme precipitation and drought events in summer (Fig. S1), which require further systematic evaluation. Those results indicated that Rs depends mainly on variations of soil moisture and temperature in the context of warming, and climate change is likely to have a very significant effect on temperate deserts.
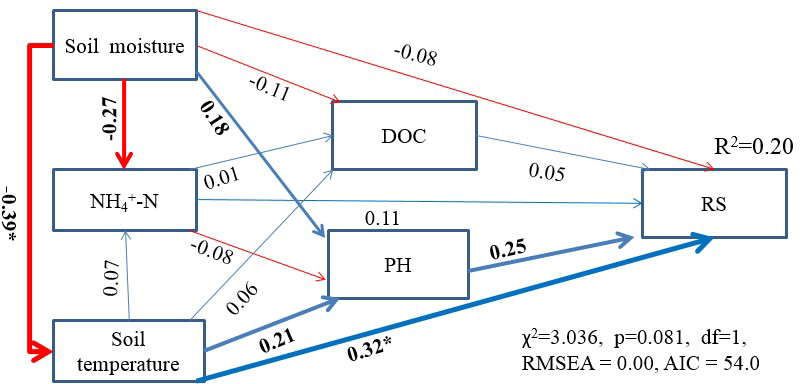
Figure 5Structure equation model (SEM) testing the multivariate (soil moisture, soil temperature, soil NH-N content, DOC and pH) effects on Rs (n = 34). Arrows show the effects of different key controls on Rs analyzed. The blue arrows indicated positive effects, and red arrows showed negative effects. The width of the arrows indicate the strength of the relationship. The numbers are standardized path coefficients, which can show the importance of the variables in the model. Goodness-of-fit statistics for the model are shown below the model. Asterisks indicate significant effect at P < 0.05.
4.2 The interactive effects of precipitation, N deposition and warming on Rs
Interactive responses of Rs were much lower than those of any single-factor, but still increased Rs overall, except with the interaction between precipitation and high N deposition (Table 1). This is consistent with results in dry ecosystems (Morillas et al., 2015; Martins et al., 2016), but not with the results of a meta-analysis of precipitation and N deposition interactive experiments which showed greater positive effects on Rs (L. Y. Zhou et al., 2016). This can be explained in our study due to soil MBC or MBN being much lower in interactive treatments than those of single-factor (Fig. 1d), showing that the number of microorganisms was much smaller in interactive treatments than that of single-factor due to the much stronger N effect. As we found that Rs was reduced with increasing N deposition and precipitation by as much as 4.25 % in W1N2 plots (Table 1), this showed that the inhibiting effect of soil NH-N content was much stronger when there was sufficient soil moisture (Fig. 2e). This is consistent with the results in a Populus euphratica community in a desert ecosystem (He et al., 2015). This was because (i) microbial activity was inhibited by high or low soil moisture and high soil NH-N or NO-N content (Burton et al., 2004) and (ii) high N content to reduce extracellular enzyme activity and the fungal population (Maris et al., 2015). In addition, the interactive effect of the three factors on Rs in this desert was much lower than interaction of two factors of precipitation and N deposition (Table 1), and is consistent with the results of modeled interactive effects, which showed that three-factor interactions were rare while two-factor interactions were more common (Luo et al., 2008). Fortunately, the interactive effect of three factors or two factors (precipitation, N deposition and warming) in this desert could largely reduce between-year variation on Rs (Table 2), which may because (i) the limits of soil moisture, soil temperature and soil N content were lifted for key biological processes by increasing precipitation, N deposition and warming (Huang et al., 2015a; R. Liu et al., 2016) and (ii) various factors counteracting each other (L. Y. Zhou et al., 2016). However, the effects of the variation on Rs in the growing season on Rs can be increased by warming, elevated precipitation and N deposition because of their dominant effects on plant growth and microbial activity (Huang et al., 2015b), but it was the exact opposite in the non-growing season due to the reduced temperature limitation (Zeng et al., 2016). Those results showed that Rs would be reduced under interactive effect of increasing rainfall, temperature and N deposition in the future, and positive carbon–climate feedback took place.
4.3 Temporal effects of the variation in treatments on Rs and controlling factors
Significant cumulative effects on Rs were found by elevated N deposition rather than increasing precipitation and warming alone (Table 2). A previous study here showed that Rs decreased due to N addition over the experiment's duration (Zhou et al., 2014), which was inconsistent with our results (Fig. 2d) because in our study, we used relatively lower rates of N addition than that Zhou et al. (2014), and the composition of microbial community and soil properties were altered gradually by long-term and high N deposition (Fig. 1c and d; Huang et al., 2015b; Zong et al., 2017). In addition, significant cumulative effects in the interaction between N deposition and precipitation or warming on Rs were also found (Table 2), and Rs decreased by 4.25 % by interaction between increasing precipitation and high N deposition (Table 1), which indicated that the response of Rs to N deposition is largely dependent on soil moisture in desert soil. This may be attributed to the antagonistic interaction between elevated N deposition and precipitation on Rh (L. Y. Zhou et al., 2016). Those results indicated that N deposition produced strong cumulative effects on Rs in this desert, and was enhanced largely with increasing soil moisture, which would reduce carbon emission from soil to atmosphere.
Regression analysis shows that Rs exhibited a unimodal change pattern with variations of soil NH-N (Fig. 4d), moisture (Fig. 4g), and temperature (Fig. 4h), and Rs was significantly positively correlated to soil dissolved organic carbon (DOC) and pH (Fig. 4e and f). However, structural equation modeling indicated that soil temperature was a more important controlling factor than soil NH-N and soil moisture (Fig. 5). This is consistent with most research results (Wu et al., 2016; Y. M. Zhou et al., 2016; Chen et al., 2017a). In addition, a large interannual variation was observed (CV = 41.4 %) during our experiment (Table 1), while the variation of annual precipitation and air temperature were only 4.41 and 7.78 %, respectively (Table 1), but close to the CV of spring root biomass of ephemeral plants (47.14 %), with 24 times greater aboveground biomass of spring ephemeral plants in 2016 than in 2015 (Cui et al., 2017), which indicated that the increase of Rs in 2016 was mainly from the root respiration of ephemeral plants. This is consistent with previous studies showing that ephemeral plants mediated the interannual variations of carbon fluxes in this desert (Huang et al., 2015c; R. Liu et al., 2016). It is different from other ecosystems where interannual variations of Rs were mainly dependent on variations in annual precipitation and air temperature (Gerard et al., 1999; Asensio et al., 2007; Chen et al., 2012). Overall, our results indicate that annual variation in Rs in this temperate desert is mainly controlled by soil temperature, but between-year variation in Rs is mainly controlled by ephemeral plants.
Climate change and elevated N deposition play important roles in controlling Rs in temperate deserts. We found that increasing precipitation and N deposition significantly increased Rs in the Gurbantunggut Desert, but warming reduced Rs, mostly because of the variation of soil moisture. In addition, we found that the interactive responses of Rs were much lower for the combination of precipitation, N deposition and warming than for any single factor. Additionally, Rs is mainly mediated by soil moisture, soil temperature and soil NH-N content, but soil temperature is the most important, with between-year variation in Rs mainly controlled by ephemeral plants. These results showed that Rs is very sensitive to increasing precipitation, N deposition and warming, and their interactive effects could reduce soil carbon emissions and thus the impacts of climate change.
All data used to generate the figures are available upon request to the corresponding author.
The supplement related to this article is available online at: https://doi.org/10.5194/bg-15-2007-2018-supplement.
The authors declare that they have no conflict of interest.
This article is part of the special issue “The 10th International Carbon Dioxide Conference (ICDC10) and the 19th WMO/IAEA Meeting on Carbon Dioxide, other Greenhouse Gases and Related Measurement Techniques (GGMT-2017) (AMT/ACP/BG/CP/ESD inter-journal SI)”. It is a result of the 19th WMO/IAEA Meeting on Carbon Dioxide, Other Greenhouse Gases, and Related Measurement Techniques (GGMT-2017), Empa Dübendorf, Switzerland, 27–31 August 2017.
This work was financially supported by the Chinese National Basic Research
Program (2014CB954202), the National Natural Science Foundation of China
(41603084, 41703131, 41425007), the special training program for science and
technology talents of ethnic minorities in Xinjiang (2016D0201) and the
Ten-Thousand Talent Program (Xuejun Liu). Keith Goulding was supported by the
Sino-UK cooperative project on nitrogen (CINAg).
Edited by: Christoph
Heinze
Reviewed by: two anonymous referees
Adams, M. B.: Ecological issues related to N deposition to natural ecosystems: research needs, Environ. Int., 29, 189–199, https://doi.org/10.1016/S0160-4120(02)00179-4, 2003.
Asensio, D., Penuelas, J., Llusia, J., Ogaya, R., and Filella, L: Interannual and interseasonal Soil CO2 efflux and VOC exchange rates in a Mediterranean holm oak forest in response to experimental drought, Soil Biol. Biochem., 39, 2471–2484, https://doi.org/10.1016/j.soilbio.2007.04.019, 2007
Batjes, N. H.: Total carbon and nitrogen in the soils of the world, Eur. J. Soil Sci., 47, 151–163, https://doi.org/10.1111/j.1365-2389.1996.tb01386.x, 1996.
Brookes, P. C.: Microbial biomass and activity measurements in soil, J. Sci. Food Agric., 36, 269–270, 1985.
Burton, A. J., Pregitzer, K. S., Crawford, J. N., Zogg, G. P., and Zak, D. R.: Simulated chronic NO deposition reduces soil respiration in northern hardwood forests, Glob. Change Biol., 10, 1080–1091, https://doi.org/10.1111/j.1365-2486.2004.00737.x, 2004.
Chen, J., Zhou, X. H., Hruska, T., Cao, J. J., Zhang, B. C., Liu, C., Liu, M., Shelton, S., Guo, L., Wei, Y. L., Wang, J. F., Xiao, S., and Wang, P.: Asymmetric diurnal and monthly responses of ecosystem carbon fluxes to experimental warming, Clean-Soil Air Water, 45, 1600557, https://doi.org/10.1002/clen.201600557, 2017a.
Chen, S. T., Huang, Y., Zou, J. W., Shi, Y. S., Lu, Y. Y., Zhang, W., and Hu, Z. H.: Interannual variability in soil respiration from terrestrial ecosystems in China and its response to climate change, Sci. China Earth. Sci., 55, 2091–2098, 2012.
Chen, W. W., Zheng, X. H., Chen, Q., Wolf', B., Butterbach-Bahl, K., Bruggemann, N., and Lin, S.: Effects of increasing precipitation and nitrogen deposition on CH4 and N2O fluxes and ecosystem respiration in a degraded steppe in Inner Mongolia, China, Geoderma, 192, 335–340. https://doi.org/10.1016/j.geoderma.2012.08.018, 2013.
Chen, Z. J., Setala, H., Geng, S. C., Han, S. J., Wang, S. Q., Dai, G. H., and Zhang, J. H.: Nitrogen addition impacts on the emissions of greenhouse gases depending on the forest type: a case study in Changbai Mountain, Northeast China, J. Soil Sediment, 17, 23–34, https://doi.org/10.1007/s11368-016-1481-7, 2017b.
Cui, X., Yue, P., Gong, Y., Li, K., Tan, D., Goulding, K., and Liu, X.: Impacts of water and nitrogen addition on nitrogen recovery in Haloxylon ammodendron dominated desert ecosystems, Sci. Total Environ., 601, 1280–1288, https://doi.org/10.1016/j.scitotenv.2017.05.202, 2017.
Eswaran, H., Van Den Berg, E., and Reich, P.: Organic carbon in soils of the world, Soil Sci. Soc. Am. J., 57, 192–194, https://doi.org/10.2136/sssaj1993.03615995005700010034x, 1993.
Fang, C., Ye, J. S., Gong, Y. H., Pei, J. Y., Yuan, Z. Q., Xie, C., Zhu, Y. S., and Yu, Y. Y.: Seasonal responses of soil respiration to warming and nitrogen addition in a semi-arid alfalfa-pasture of the Loess Plateau, China, Sci. Total Environ., 590, 729–738, https://doi.org/10.1016/j.scitotenv.2017.03.034, 2017.
Feng, J. G., Wang, J. S., Ding, L. B., Yao, P. P., Qiao, M. P., and Yao, S. C.: Meta-analyses of the effects of major global change drivers on soil respiration across China, Atmos. Environ., 150, 181–186, https://doi.org/10.1016/j.atmosenv.2016.11.060, 2017.
Galloway, J. N., Townsend, A. R., Erisman, J. W., Bekunda, M., Cai, Z., Freney, J. R., and Sutton, M. A. . Transformation of the nitrogen cycle: recent trends, questions, and potential solutions, Science, 320, 889–892. https://doi.org/10.1126/science.1136674, 2008.
Gerard, J. C., Nemry, B., Francois, L. M., and Warnant, P.: The interannual change of atmospheric CO2: contribution of subtropical ecosystems?, Geophys. Res. Lett., 26, 243–246, https://doi.org/10.1029/1998GL900269, 1999.
Gougoulias, C., Clark, J. M., and Shaw, L. J.: The role of soil microbes in the global carbon cycle: tracking the below – microbial processing of plant – carbon for manipulating carbon dynamics in agricultural systems, J. Sci. Food Agr., 94, 2362–2371, https://doi.org/10.1002/jsfa.6577, 2014.
Hanson, P. J., Edwards, N. T., Garten, C. T., and Andrews, J. A.: Separating root and soil microbial contributions to soil respiration: A review of methods and observations, Biogeochemistry, 48, 115–146, https://doi.org/10.1023/A:1006244819642, 2000.
He, X., Lv, G., Qin, L., Chang, S., Yang, M., Yang, J., and Yang, X.: Effects of simulated nitrogen deposition on soil respiration in a Populus euphratica community in the Ebinur Lake area, a desert ecosystem of northwestern China, PloS one, 10, 1–16, https://doi.org/10.1371/journal.pone.0137827, 2015.
Heimann, M. and Reichstein, M.: Terrestrial ecosystem carbon dynamics and climate feedbacks, Nature, 451, 289–292, https://doi.org/10.1038/nature06591, 2008.
Huang, G., Li, Y., and Su, Y. G.: Effects of increasing precipitation on soil microbial community composition and soil respiration in a temperate desert, Northwestern China, Soil Biol. Biochem., 83, 52–56, https://doi.org/10.1016/j.soilbio.2015.01.007, 2015a.
Huang, G., Cao, Y. F., Wang, B., and Li, Y.; Effects of nitrogen addition on soil microbes and their implications for soil C emission in the Gurbantunggut Desert, Center of the Eurasian Continent, Sci. Total Environ., 515, 215–224, https://doi.org/10.1016/j.scitotenv.2015.01.054, 2015b.
Huang, G., Li, Y., and Padilla, F. M.: Ephemeral plants mediate responses of ecosystem carbon exchange to increased precipitation in a temperate desert, Agr. Forest Meteorol., 201, 141–152, https://doi.org/10.1016/j.agrformet.2014.11.011, 2015c.
IPCC: Climate Change 2013: the Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge University Press, Cambridge, UK and New York, NY, USA, 1535 pp., 2013.
Jiang, X., Cao, L., and Zhang, R.: Changes of labile and recalcitrant carbon pools under nitrogen addition in a city lawn soil, J. Soils Sediments, 14, 515–524, https://doi.org/10.1007/s11368-013-0822-z, 2014.
Jones, D. L. and Willett, V. B.: Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil, Soil Biology and Biochemistry, 38, 991–999, https://doi.org/10.1016/j.soilbio.2005.08.012, 2006.
Lafleur, P. M. and Humphreys, E. R.,: Spring warming and carbon dioxide exchange over low Arctic tundra in central Canada, Glob. Change Biol., 14, 740–756, https://doi.org/10.1111/j.1365-2486.2007.01529.x, 2008.
Li, G., Kim, S., Han, S. H., Chang, H., and Son, Y.: Effect of soil moisture on the response of soil respiration to open-field experimental warming and precipitation manipulation, Forests, 8, 1–10, 2017.
Liu, C., Wang, K., and Zheng, X.: Responses of N2O and CH4 fluxes to fertilizer nitrogen addition rates in an irrigated wheat-maize cropping system in northern China, Biogeosciences, 9, 839–850, https://doi.org/10.5194/bg-9-839-2012, 2012.
Liu, L. L., Wang, X., Lajeunesse, M. J., Miao, G. F., Piao, S. L., Wan, S. Q., Wu, Y. X., Wang, Z. H., Yang, S., Li, P., and Deng, M. F.: A cross-biome synthesis of soil respiration and its determinants under simulated precipitation changes, Glob. Change Biol., 22, 1394–1405, https://doi.org/10.1111/gcb.13156, 2016.
Liu, L. T., Hu, C. S., Yang, P. P., Ju, Z. Q., Olesen, J. E., and Tang, J. W.: Effects of experimental warming and nitrogen addition on soil respiration and CH4 fluxes from crop rotations of winter wheat-soybean/fallow, Agr. Forest Meteorol., 207, 38–47, https://doi.org/10.1016/j.agrformet.2015.03.013, 2015.
Liu, R., Cieraad, E., Li, Y., and Ma, J.: Precipitation pattern determines the inter-annual variation of herbaceous layer and carbon fluxes in a phreatophyte-dominated desert ecosystem, Ecosystems, 19, 601–614, https://doi.org/10.1007/s10021-015-9954-x, 2016.
Liu, T., Xu, Z. Z., Hou, Y. H., and Zhou, G. S.: Effects of warming and changing precipitation rates on soil respiration over two years in a desert steppe of northern China, Plant Soil., 400, 15–27, https://doi.org/10.1007/s11104-015-2705-0, 2016.
Liu, Y., Li, X., Zhang, Q., Guo, Y., Gao, G., and Wang, J.: Simulation of regional temperature and precipitation in the past 50 years and the next 30 years over China, Quatern. Int., 212, 57–63 https://doi.org/10.1016/j.quaint.2009.01.007, 2010.
Luo, C. Y., Wang, S. P., Zhao, L., Xu, S. X., Xu, B. R. B. Y., Zhang, Z. H., Yao, B. Q., and Zhao, X. Q.: Effects of land use and nitrogen fertilizer on ecosystem respiration in alpine meadow on the Tibetan Plateau, J. Soil Sediment., 17, 1626–1634, https://doi.org/10.1016/j.quaint.2009.01.007, 2017.
Luo, Y., Gerten, D., Le Maire, G., Parton, W. J., Weng, E., Zhou, X., Keough, C., Beier, C., Ciais, P., and Cramer, W.: Modeled interactive effects of precipitation, temperature, and CO2 on ecosystem carbon and water dynamics in different climatic zones, Glob. Change Biol., 14, 1986–1999, https://doi.org/10.1111/j.1365-2486.2008.01629.x, 2008.
Ma, J., Wang, Z.-Y., Stevenson, B. A., Zheng, X.-J., and Li, Y.: An inorganic CO2 diffusion and dissolution process explains negative CO2 fluxes in saline/alkaline soils, Sci. Rep-Uk., 3, 2025, https://doi.org/10.1038/srep02025, 2013.
Marion, G. M., Henry, G. H. R., Freckman, D. W., Johnstone, J., Jones, G., Jones, M. H., and Virginia, R. A.: Open-top designs for manipulating field temperature in high-latitude ecosystems, Glob. Change Biol., 1997, 3, 20–32, https://doi.org/10.1111/j.1365-2486.1997.gcb136.x, 2003.
Maris, S. C., Teira-Esmatges, M. R., Arbonés, A., and Rufat, J.: Effect of irrigation, nitrogen application, and a nitrification inhibitor on nitrous oxide, carbon dioxide and methane emissions from an olive (Olea europaea L.) orchard, Sci. Total Environ., 538, 966–978, https://doi.org/10.1016/j.scitotenv.2015.08.040, 2015.
Martins, C. S. C., Macdonald, C. A., Anderson, I. C., and Singh, B. K.: Feedback responses of soil greenhouse gas emissions to climate change are modulated by soil characteristics in dryland ecosystems, Soil Biol. Biochem., 100, 21–32, https://doi.org/10.1016/j.soilbio.2016.05.007, 2016.
Morillas, L., Duran, J., Rodriguez, A., Roales, J., Gallardo, A., Lovett, G. M., and Groffman, P. M.: Nitrogen supply modulates the effect of changes in drying-rewetting frequency on soil C and N cycling and greenhouse gas exchange Global Change Biol., 21, 3854–3863, https://doi.org/10.1111/gcb.12956, 2015.
Noh, N. J., Kuribayashi, M., Saitoh, T. M., Nakaji, T., Nakamura, M., Hiura, T., and Muraoka, H.: Responses of soil, heterotrophic, and autotrophic respiration to experimental open-field soil warming in a cool-temperate deciduous forest, Ecosystems, 19, 504–520, https://doi.org/10.1007/s10021-015-9948-8, 2016.
Sheik, C. S., Beasley, W. H., Elshahed, M. S., Zhou, X., Luo, Y., and Krumholz, L. R.: Effect of warming and drought on grassland microbial communities, The ISME Journal, 5, 1692–1700, https://doi.org/10.1038/ismej.2011.32, 2011.
Skopp, J., Jawson, M. D., and Doran, J. W.: Steady-state aerobic microbial activity as a functionof soil watercontent, Soil Sci. Soc. Am., 54, 1619–1625, https://doi.org/10.2136/sssaj1990.03615995005400060018x, 1990.
Song ,W.:Characteristics of Chemical Composition and Deposition Fluxes of Atmospheric Particles and Reactive Nitrogen in Typical Ecosystems of Xinjiang, China, PhD Thesis, Xinjiang Institute of Ecology and Geography, China, 2015.
Strong, A. L., Johnson, T. P., Chiariello, N. R., and Field, C. B.: Experimental fire increases soil carbon dioxide efflux in a grassland long-term multifactor global change experiment, Global Change Biol., 23, 1975–1987, https://doi.org/10.1111/gcb.13525, 2017.
Wang, Q. K., Zhang, W. D., Sun, T., Chen, L. C., Pang, X. Y., Wang, Y. P., and Xiao, F. M.: N and P fertilization reduced soil autotrophic and heterotrophic respiration in a young Cunninghamia lanceolata forest, Agr. Forest Meteorol., 232, 66–73, https://doi.org/10.1016/j.agrformet.2016.08.007, 2017.
Wu, C. S., Liang, N. S., Sha, L. Q., Xu, X. L., Zhang, Y. P., Lu, H. Z., Song, L., Song, Q. H., and Xie, Y. N.: Heterotrophic respiration does not acclimate to continuous warming in a subtropical forest, Sci. Rep-Uk, 6, 21561, https://doi.org/10.1038/srep21561, 2016.
Wu, Z. T., Dijkstra, P., Koch, G. W., Penuelas, J., and Hungate, B. A.: Responses of terrestrial ecosystems to temperature and precipitation change: a meta-analysis of experimental manipulation, Glob. Change Biol., 17, 927–942, https://doi.org/10.1111/j.1365-2486.2010.02302.x, 2011.
Yang, G., Wang, M., Chen, H., Liu, L. F., Wu, N., Zhu, D., Tian, J. Q., Peng, C. H., Zhu, Q. A., and He, Y. X.: Responses of CO2 emission and pore water DOC concentration to soil warming and water table drawdown in Zoige Peatlands, Atmos. Environ., 152, 323–329, https://doi.org/10.1016/j.atmosenv.2016.12.051, 2017.
Yue, P., Li, K., Gong, Y., Hu, Y., Mohammat, A., Christie, P., and Liu, X.: A five-year study of the impact of nitrogen addition on methane uptake in alpine grassland, Sci. Rep-UK, 6, 32064, https://doi.org/10.1038/srep32064, 2016.
Zeng, X. H., Song, Y. G., Zeng, C. M., Zhang, W. J., and He, S. B.: Partitioning soil respiration in two typical forests in semi-arid regions, North China, Catena, 147, 536–544, https://doi.org/10.1016/j.catena.2016.08.009, 2016.
Zhang, N., Liu, W., Yang, H., Yu, X., Gutknecht, J. L. M., Zhang, Z., Wan, S., and Ma, K.: Soil microbial responses to warming and increased precipitation and their implications for ecosystem C cycling, Oecologia, 173, 1125–1142, https://doi.org/10.1007/s00442-013-2685-9, 2013.
Zhang, W., Liu, C. Y., Zheng, X. H., Fu, Y. F., Hu, X. X., Cao, G. M., and Butterbach-Bahl, K.: The increasing distribution area of zokor mounds weaken greenhouse gas uptakes by alpine meadows in the Qinghai-Tibetan Plateau, Soil Biol. Biochem., 71, 105–112, https://doi.org/10.1016/j.soilbio.2014.01.005, 2014.
Zhang, X. L., Tan, Y. L., Zhang, B. W., Li, A., Daryanto, S., Wang, L. X., and Huang, J. H.: The impacts of precipitation increase and nitrogen addition on soil respiration in a semiarid temperate steppe, Ecosphere, 8, e01655, https://doi.org/10.1002/ecs2.1655, 2017.
Zhao, Z. Z., Dong, S. K., Jiang, X. M., Liu, S. L., Ji, H. Z., Li, Y., Han, Y. H., and Sha, W.: Effects of warming and nitrogen deposition on CH4, CO2 and N2O emissions in alpine grassland ecosystems of the Qinghai-Tibetan Plateau, Sci. Total Environ., 592, 565–572, https://doi.org/10.1016/j.scitotenv.2017.03.082, 2017.
Zhong, Y. Q. W., Yan, W. M., and Shangguan, Z. P.: The effects of nitrogen enrichment on soil CO2 fluxes depending on temperature and soil properties, Global Ecol. Biogeogr., 25, 475–488, https://doi.org/10.1111/geb.12430, 2016.
Zhou, L. Y., Zhou, X. H., Shao, J. J., Nie, Y. Y., He, Y. H., Jiang, L. L., Wu, Z. T., and Bai, S. H.: Interactive effects of global change factors on soil respiration and its components: a meta-analysis, Glob. Change Biol., 22, 3157–3169, https://doi.org/10.1111/gcb.13253, 2016.
Zhou, X. B. and Zhang, Y. M.: Seasonal pattern of soil respiration and gradual changing effects of nitrogen addition in a soil of the Gurbantunggut Desert, northwestern China, Atmos. Environ., 85, 187–194, https://doi.org/10.1016/j.atmosenv.2013.12.024, 2014.
Zhou, Y. M., Hagedorn, F., Zhou, C. L., Jiang, X. J., Wang, X. X., and Li, M. H.: Experimental warming of a mountain tundra increases soil CO2 effluxes and enhances CH4 and N2O uptake at Changbai Mountain, China, Sci Rep-UK, 6, 21108, https://doi.org/10.1038/srep21108, 2016.
Zhu, J., Kang, F. F., Chen, J., Cheng, X. Q., and Han, H. R.: Effect of nitrogen addition on soil respiration in a Larch Plantation, Pol. J. Environ. Stud., 26, 1403–1412, https://doi.org/10.15244/pjoes/67687, 2017.
Zong, N., Shi, P. L., Chai, X., Jiang, J., Zhang, X. Z., and Song, M. H.: Responses of ecosystem respiration to nitrogen enrichment and clipping mediated by soil acidification in an alpine meadow, Pedobiologia, 60, 1–10, https://doi.org/10.1016/j.pedobi.2016.11.001, 2017.