the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 26 Mar 2021
| 26 Mar 2021
On the barium–oxygen consumption relationship in the Mediterranean Sea: implications for mesopelagic marine snow remineralization
Stéphanie H. M. Jacquet
Dominique Lefèvre
Christian Tamburini
Marc Garel
Frédéric A. C. Le Moigne
Nagib Bhairy
Sophie Guasco
In the ocean, remineralization rate associated with sinking particles is a crucial variable. Since the 1990s, particulate biogenic barium (Baxs) has been used as an indicator of carbon remineralization by applying a transfer function relating Baxs to O2 consumption (Dehairs's transfer function, Southern Ocean-based). Here, we tested its validity in the Mediterranean Sea (ANTARES/EMSO-LO) for the first time by investigating connections between Baxs, prokaryotic heterotrophic production (PHP) and oxygen consumption (JO2-Opt; optodes measurement). We show that (1) higher Baxs (409 pM; 100–500 m) occurs in situations where integrated PHP (PHP) is located deeper, (2) higher Baxs occurs with increasing JO2-Opt, and (3) there is similar magnitude between JO2-Opt (3.14 mmol m−2 d−1; 175–450 m) and JO2-Ba (4.59 mmol m−2 d−1; transfer function). Overall, Baxs, PHP and JO2 relationships follow trends observed earlier in the Southern Ocean. We conclude that such a transfer function could apply in the Mediterranean Sea.
- Article
(944 KB) - Full-text XML
- BibTeX
- EndNote





Ocean ecosystems play a critical role in the Earth's carbon (C) cycle (IPCC, 2014). The quantification of their impacts of both present conditions and future predictions remains one of the greatest challenges in oceanography (Siegel et al., 2016). In essence, the biological C pump is termed for the numerous processes involved in maintaining the vertical gradient in dissolved inorganic C. This includes processes such as organic matter production at the surface, its export and subsequent remineralization. Most of marine snow organic C conversion (i.e., remineralization) into CO2 by heterotrophic organisms (i.e., respiration) occurs in the mesopelagic zone (100–1000 m) (Martin et al., 1987; Buesseler and Boyd, 2009). Globally, the flux of C exported below 1000 m depth is the key determinant of ocean carbon storage capacity (Henson et al., 2011). However, there is no consensus on C transfer efficiency estimations from field experiments, leading to an imbalance of the water column C budget (Giering et al., 2014). Resolving this imbalance is at the core of numerous studies in the global ocean, but also regionally, especially in the Mediterranean Sea (MedSea). Due to limited exchanges with the adjacent basin and the existence of an intense overturning circulation qualitatively resembling the global one (but with shorter timescales), the MedSea is often considered a laboratory to observe and understand the impact of transient climate variability on ecosystems and biogeochemical cycles (Malanotte-Rizzoli et al., 2014). In a context of climate changes, better constraining C fluxes and the ocean C storage capacity is of crucial importance.
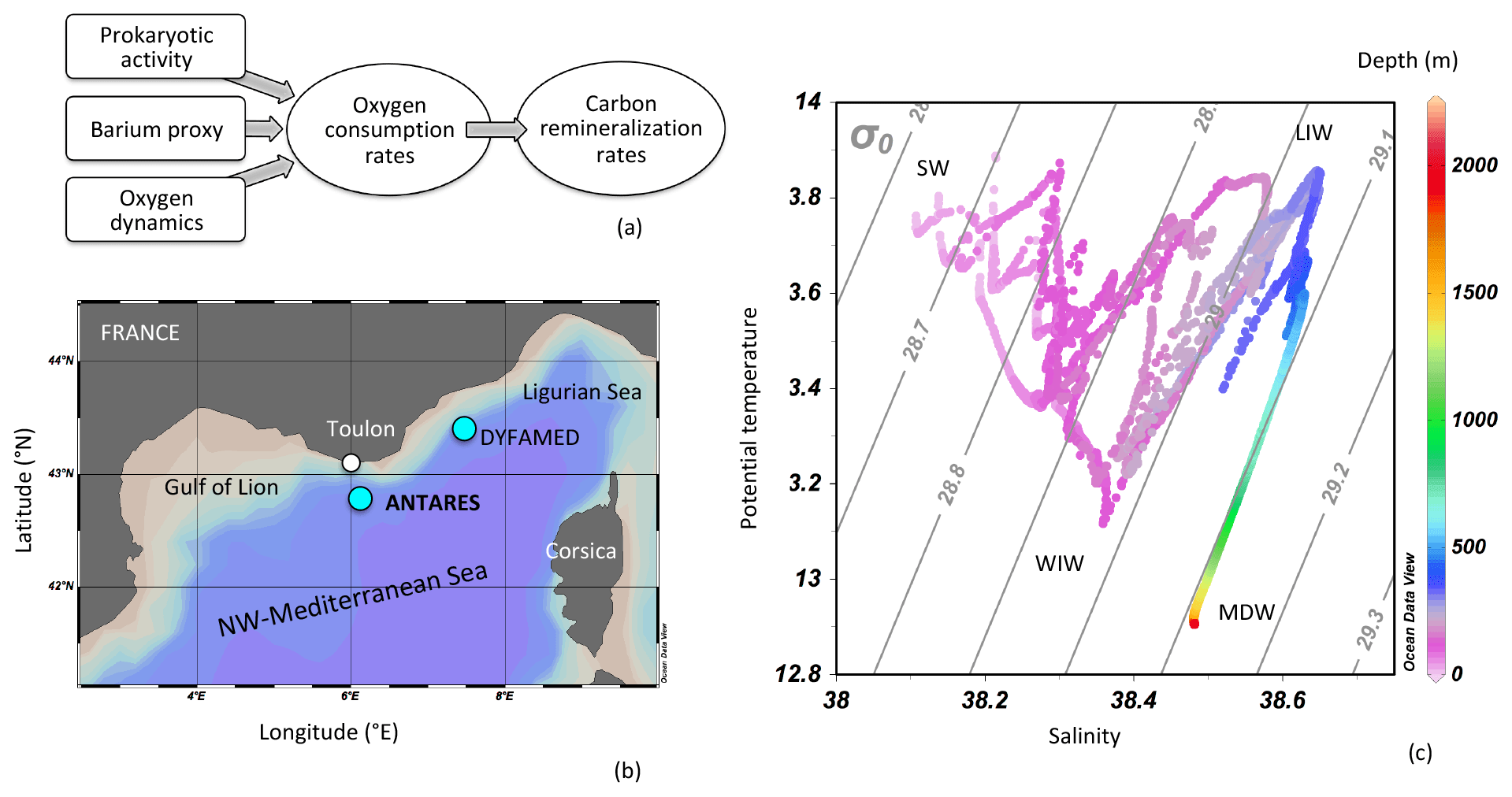
Figure 1(a) Schematic representation of the convergence of the different estimators of oxygen consumption and C remineralization rates from the “oxygen dynamics”, “barium proxy” and “prokaryotic activity” tools; (b) location of the BATMAN cruise at the ANTARES observatory site in the NW Mediterranean Sea (42∘48′ N, 6∘10′ E). The location of the DYFAMED station is reported for comparison (Sternberg et al., 2008); (c) potential temperature–salinity–depth plots and isopycnals for BATMAN profiles. SW: Surface Water; WIW: Winter Intermediate Water; LIW: Levantine Intermediate Water; DMW: Deep Mediterranean Water. Graph constructed using Ocean Data View (Schlitzer, 2002; Ocean Data View; http://www.awi-bremerhaven.de/GEO/ODV, last access: 2 March 2021).
Particulate barium in excess (Baxs, i.e., biogenic Ba from total particulate Ba after correction for lithogenic Ba) is a geochemical tracer of particulate organic carbon (POC) remineralization in the mesopelagic layer (Dehairs et al., 1997). Baxs mostly occurs in the form of barite microcrystals (BaSO4) at these depths. In a global ocean undersaturated with respect to barite, studies report that Baxs would precipitate inside oversaturated biogenic micro-environments during POC degradation by heterotrophic prokaryotes in the mesopelagic zone, through sulfate and/or barium enrichment (Bertram and Cowen, 1997). The first-ever studies on mesopelagic Baxs reported coinciding Baxs maxima with depths of dissolved O2 minimum and pCO2 maximum (Dehairs et al., 1987, 1997). By using an 1D advection–diffusion model applied to O2 profiles in the Atlantic sector of the Southern Ocean (ANTX/6 cruise; Shopova et al., 1995), Dehairs et al. (1997) established an algorithm converting mesopelagic Baxs concentration into O2 consumption rate (JO2) and remineralized organic C (POC remineralization rate). This transfer function has been widely used until now (Cardinal et al., 2001, 2005; Dehairs et al., 2008; Jacquet et al., 2008a, b, 2011, 2015). Yet its validity has never been tested in other oceanic provinces. In the North Atlantic, Lemaitre et al. (2018) reported a Baxs–JO2 (obtained from apparent oxygen utilization divided by the water mass age) relationship not significantly different to that reported in Dehairs et al. (1997). Furthermore, significant progress was made in relating Baxs and O2 dynamics to prokaryotic heterotrophic activity (Jacquet et al., 2015). These advancements clearly show that Baxs is closely related with the vertical distribution of prokaryote heterotrophic production (PHP) (the rate of change with depth), reflecting the temporal progression of POC remineralization processes. Also, in a first attempt to test the validity of the Dehairs transfer function in other locations, Jacquet et al. (2015) compared oxygen consumption rates (JO2) from direct measurements (dark community respiration, DCR) to derived JO2 from Baxs data (using the transfer function) in the Kerguelen area (Indian sector of the Southern Ocean). We revealed good convergence of JO2 rates from these two approaches, further supporting the Dehairs function to estimate POC remineralization rates in different biogeochemical settings of the Southern Ocean.
Here, we further investigate relationships between the mesopelagic Baxs proxy, prokaryotic activity and oxygen dynamics (Fig. 1a) in the northwestern Mediterranean Sea (MedSea), a different biogeochemical setting to those already studied (see references above). Today, observations of the various components of the MedSea biological C pump provide organic C remineralization fluxes varying by at least an order of magnitude (Santinelli et al., 2010; Ramondenc et al., 2016). Malanotte-Rizzoli et al. (2014) reviewing unsolved issues and future directions for MedSea research highlighted the need to further investigate biogeochemical processes at intermediate (mesopelagic) and deep layers to reconciliate the C budget in the Mediterranean basin. The previous particulate Baxs dataset is very scarce in the NW MedSea, with generally very low vertical sampling resolution (Sanchez-Vidal et al., 2005) or very restricted studied areas (Dehairs et al., 1987; Sternberg et al., 2008). Here we discuss Baxs, PHP and JO2 (from optode measurement during incubations) at the ANTARES / EMSO-LO observatory site (Fig. 1a and b). We hypothesize that the Dehairs transfer function converting Baxs into POC remineralization also applies in a different ocean ecosystem from the Southern Ocean. We suggest that the Baxs proxy can be used as routine tracer to estimate local-scale processes of mesopelagic POC remineralization in the Mediterranean basin.
2.1 Study site
The BATMAN cruise (https://doi.org/10.17600/16011100, 10–16 March 2016, R/V L'EUROPE) took place at the ANTARES/EMSO-LO observatory site (42∘48′ N, 6∘10′ E; Tamburini et al., 2013), 40 km off the coast of Toulon, southern France (Fig. 1b). The hydrological and biogeochemical conditions at this site are monitored monthly in the framework of the MOOSE (Mediterranean Ocean Observing System for the Environment) program and of the EMSO (European Multidisciplinary Subsea Observatory) observation program. The hydrography displays the general three-layer MedSea system with surface, intermediate and deep waters (Hainbucher et al., 2014). Briefly, the main water masses can be distinguished (see potential temperature–salinity diagram during the BATMAN cruise in Fig. 1c): (1) Surface Water (SW); (2) Winter Intermediate Water (WIW); (3) Levantine Intermediate water (LIW); (4) Mediterranean Deep Water (MDW). LIW is present at intermediate depths (around 400 m at ANTARES) and is characterized by temperature and a salinity maxima.
2.2 Sampling and analyses
For particulate barium, 4 to 7 L of seawater sampled using Niskin bottles was filtered onto 47 mm polycarbonate membranes (0.4 µm porosity) under slight overpressure supplied by filtered air. Filters were rinsed with a few milliliters of Milli-Q grade water to remove sea salt, dried (50 ∘C) and stored in petri dishes. Thirteen depths between the surface and 1000 m were sampled by combining different casts sampled closely in time and space (total of 28 samples) with similar potential temperature–salinity profiles. No major changes in water mass characteristics occurred over the 3 d sampling period (Fig. 1c). In the laboratory, we performed a total digestion of filters using a concentrated triacid (0.5 mL HF, 1.5 mL HNO3, HCl 1 mL; all Optima grade) mixture in closed teflon beakers overnight at 95 ∘C in a clean pressurized room. After evaporation close to dryness, samples were re-dissolved into 10 mL of HNO3 2 %. The solutions were analyzed for Ba and other elements of interest (Na and Al) by HR-ICP-MS (high-resolution inductively coupled plasma mass spectrometry; ELEMENT XR Thermo Fisher). Based on analyses of external certified reference standards, accuracy and reproducibility were both within ±5 %. Details on sample processing and analysis are given in Cardinal et al. (2001) and Jacquet et al. (2015). The presence of sea salt was checked by analyzing Na, and the sea salt particulate Ba contribution was found negligible (<0.1 % of total Ba). Particulate biogenic barium in excess (hereafter referred to as Baxs) was calculated as the difference between total Ba and lithogenic Ba using Al as the lithogenic reference element. The lithogenic Ba concentration was determined using Al concentration and the upper continental crust (UCC) Ba : Al molar ratio (Taylor and McLennan, 1985). The biogenic Ba fraction ranged from 51 % to 91 % of the total particulate Ba signal (see Sect. 3.1). The standard uncertainty (Ellison, 2000) on Baxs concentration ranges between 5.0 % and 5.5 %. The term “in excess” is used to indicate that concentrations are larger than the Baxs background. The background (or residual value) is considered to be “preformed” Baxs at zero oxygen consumption left over after transfer and partial dissolution of Baxs produced during degradation of previous phytoplankton growth events. The background is set at 130 pM in this study.
Oxygen concentrations were measured using oxygen optode Aanderaa® 4330 for at least 24 h (on a 30 s time step) on samples taken at four depths in the mesopelagic layer (175, 250, 450 and 1000 m). Samples were placed into sealed 1 L borosilicate glass bottles incubated at a constant temperature of 13 ∘C in thermoregulated baths. Optodes were calibrated using a homemade calibration facility. Oxygen consumption rates (later referred to as JO2-Opt) were derived from a linear model calculation. Associated errors to the linear model fit are below 0.01 µM O2 h−1. Each oxygen consumption experiment has been duplicated for each depth. Average and standard deviation of the duplicates are reported in Fig. 3a. The larger associated errors are related to the differences between each duplicate, especially at the surface, reflecting potential heterogeneity of the microbial community during sampling.
Prokaryotic heterotrophic production (PHP) estimation was measured over time course experiments at in situ temperature (13 ∘C) following the protocol described in Tamburini et al. (2002). 3H-leucine-labeled tracer (Kirchman, 1993) was used. For water sample collected with Niskin bottles, we have performed measurements in three replicates of 20 and 40 mL seawater volume for the depth ranging from 0 to 800 m with 20 nM of leucine at final concentration. Concerning depth above 800 m depth, PHP was measured in three replicate of 40 mL of seawater with 10nM at final concentration of Leucine. Samples were incubated 2, 6 and 10 h respectively for samples ranging between 0–200 m, 200–600 m and up to 800 m depth. To calculate prokaryotic heterotrophic production, we used the empirical conversion factor of 1.55 ng C pmol−1 of incorporated leucine according to Simon and Azam (1989), assuming that isotope dilution was negligible under these saturating concentrations.
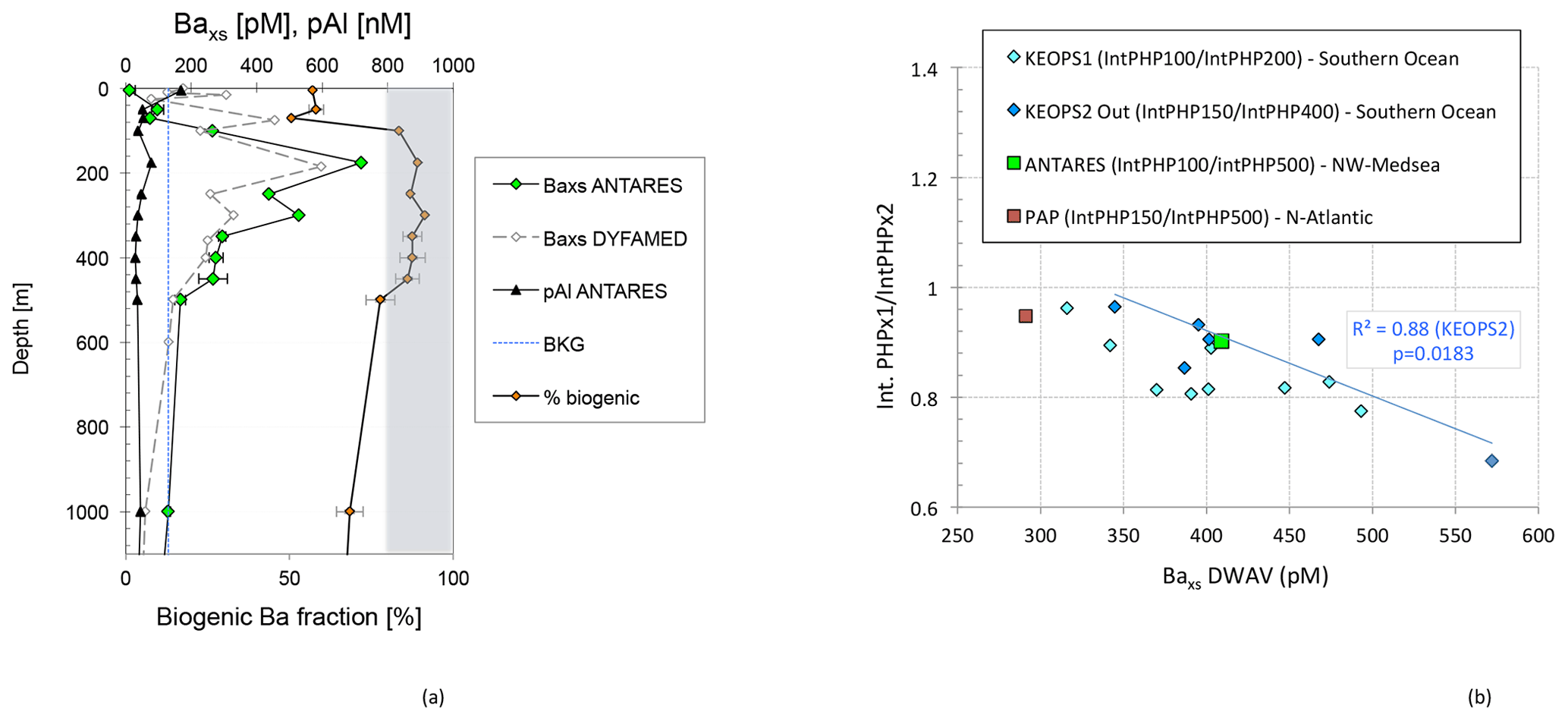
Figure 2(a) Particulate biogenic Baxs (pM) and particulate Al (nM) profiles next to the biogenic Ba fraction (%) in the upper 1000 m at ANTARES. The grey area represents a biogenic Ba fraction larger than 80 %. BKG: Baxs background. Baxs profile (pM) at DYFAMED: data from Sternberg et al. (2008). (b) ANTARES ratio plot (green square) of integrated PHP in the upper 100 m over integrated PHP in the upper 500 m versus depth-weighted average (DWA) mesopelagic Baxs (pM) over the 100–500 m depth interval. Regression of the same ratio is reported for KEOPS1 (light blue symbols; out plateau stations) and KEOPS2 (dark blue symbols; Southern Ocean, Jacquet et al., 2015) and DY032 (red square; PAP station, NE Atlantic; pers. data) cruises. The blue line represents the trend obtained during KEOPS2 (Jacquet et al., 2015).
3.1 Particulate Baxs vertical distribution
Particulate biogenic Baxs, particulate Al (pAl) and biogenic Ba fraction profiles in the upper 1000 m at ANTARES are reported in Fig. 2a. Baxs concentrations range from 12 to 719 pM. The biogenic Ba fraction ranges from 51 % to 91 % of the total particulate Ba signal. Particulate Al concentrations (pAl) range from 8 to 170 nM. Baxs concentrations are low in surface water (<100 pM) where the lithogenic fraction reaches 43 % to 49 % in the upper 70 m. From previous studies we know that Baxs in surface waters is distributed over different, mainly non-barite biogenic phases and incorporated into or adsorbed onto phytoplankton material. As such these do not reflect POC remineralization processes, in contrast to mesopelagic waters where Baxs is mainly composed of barite formed during prokaryotic degradation of organic matter. Focus in the present study is on the mesopelagic layer. The Baxs profile at ANTARES indeed displays a mesopelagic Baxs maximum between 100 and 500 m, reaching up to 719 pM at 175 m. Ba is mostly biogenic at these depths (>80 %). Baxs concentrations then decrease below 500 m to reach a background value of around 130 pM (see BKG in Fig. 2). Note that the MedSea is largely undersaturated with respect to barite, with saturation state ranging between 0.2 and 0.6 over the basin (Jacquet et al., 2016; Jullion et al., 2017). For comparison, the Baxs background value in the Southern Ocean reaches 180 to 200 pM below 1000 m (Dehairs et al., 1997; Jacquet et al., 2015). Previously, Sternberg et al. (2008) reported the seasonal evolution of Baxs profiles at the DYFAMED station (43∘25′ N, 7∘52′ E; BARMED project) northeast from ANTARES (Fig. 1c) in the NW MedSea. The present Baxs profile at ANTARES (March 2016) is very similar to the Baxs profile measured in March 2003 at DYFAMED (Fig. 2a). The slight difference between Baxs profiles in the upper 75 m suggests more Ba bounded and/or adsorbed onto phytoplankton material during BARMED. Both profiles present a Baxs maximum in the upper mesopelagic zone between 150 and 200 m. Below this maximum, Baxs concentrations gradually decrease to reach around 130 pM between 500 and 1000 m (this study). A similar value was reached between 500 and 600 m at the DYFAMED station over the whole studied period (between February and June 2003; Sternberg et al., 2008).
3.2 Prokaryotic heterotrophic production
The particulate Ba in excess is centered in the upper mesopelagic zone between 100 and 500 m and reflects the fact that POC remineralization mainly occurred at this depth layer (Fig. 2a). Depth-weighted average (DWA) Baxs content (409 pM), i.e., the Baxs inventory divided by the depth layer considered, was calculated between 100 and 500 m. Figure 2b shows the column-integrated PHP at 100 m over that at 500 m (PHP). Our PHP ratio at ANTARES station is 0.90 and is compared to results obtained during KEOPS1 (summer) and KEOPS2 (spring; out plateau stations) cruises in the Southern Ocean (Jacquet et al., 2008a, 2015) and the DY032 cruise (July 2015, R/V Discovery) at the PAP (Porcupine Abyssal Plain) observatory in the northeast Atlantic (49∘ N, 16.5∘ W) (personal data). Results at the ANTARES/EMSO-LO site follow the trend previously reported in the Southern Ocean (blue line in Fig. 2b; Jacquet et al., 2015), indicating higher DWA Baxs in situations where a significant part of column-integrated PHP is located deeper in the water column (high Int. PHPx1 IntPHPx2 ratio; Fig. 2b). These previous studies revealed that the shape of the column-integrated PHP profile (i.e., the attenuation gradient) is important in setting the Baxs signal in the mesopelagic zone (Dehairs et al., 2008; Jacquet et al., 2008a, 2015). Indeed, mesopelagic DWA Baxs appears reduced when most of the column-integrated PHP is limited to the upper layer, i.e., indicating an efficient remineralization at the surface. In contrast, mesopelagic DWA Baxs appears higher when most of the column-integrated PHP is located in the mesopelagic layer, i.e., reflecting significant deep PHP activity, POC export and subsequent remineralization (Fig. 2b). Results at the PAP site reflect a similar situation as observed during KEOPS2 for time series stations at the plateau site and in a meander of the polar front area (not show in Fig. 2b). At these stations, Jacquet et al. (2015) reported a shift toward the KEOPS1 trend reflecting the temporal evolution (season advancement) and patchiness of the establishment of mesopelagic remineralization processes within the same area. Overall, our MedSea result is located along the trend defined in the Southern Ocean during the KEOPS1 cruise. It is generally considered that Baxs (barite) forms inside sulfate and/or barium oversaturated biogenic micro-environments during POC degradation by heterotrophic prokaryotes. However, it is unclear whether barite formation at mesopelagic depths is (directly or indirectly) bacterially induced or bacterially influenced (Martinez-Ruiz et al., 2018, 2019). In any case our results strengthen the close link between the water column Baxs distribution and respiration (organic matter degradation).
3.3 Oxygen–barium relationship
The relationship we obtained at ANTARES between Baxs concentrations and oxygen consumption rates from optode measurements (JO2-Opt) is reported in Fig. 3a. JO2-Opt ranges from 0.11 to 5.85 µmol L−1 d−1. The relationship indicates higher Baxs concentrations with increasing JO2-Opt. An interesting feature is the intercept at zero JO2-Opt (around 128 pM) which further supports the Ba BKG value at ANTARES (130 pM) determined from measured Baxs profiles (Fig. 3a).
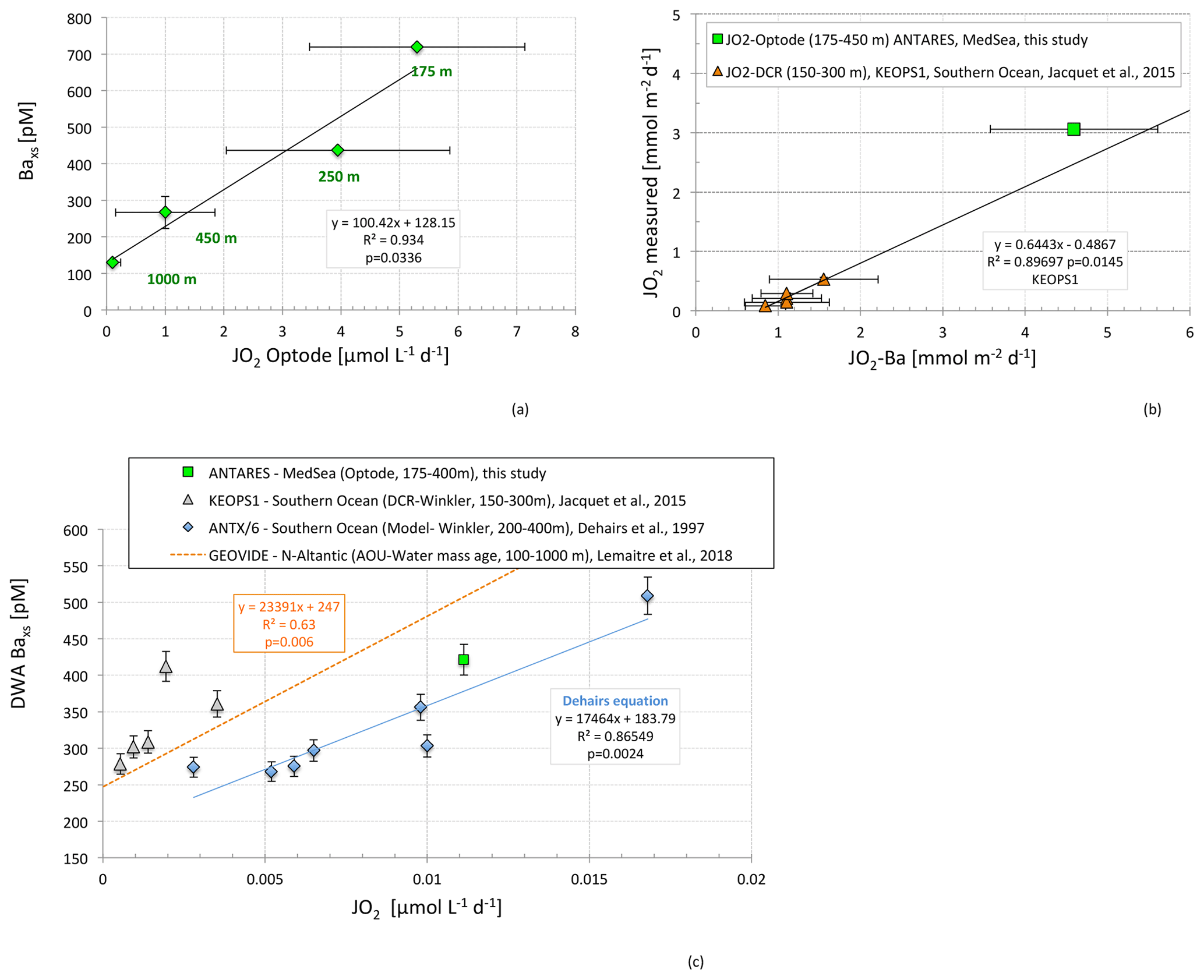
Figure 3(a) Relationship between Baxs concentrations (pM) and oxygen consumption rates (µmol L−1 d−1) from optode measurements (JO2-Opt) at ANTARES; (b) confrontation of oxygen consumption rates (JO2; mmol m−2 d−1) obtained from different methods: optode measurements (this study; green square), dark community respiration (DCR) (Winkler titration; red triangles; JO2-DCR; Jacquet et al., 2015; KEOPS1) and Dehairs's transfer function calculation (Dehairs et al., 1997) based on Baxs contents. The black line corresponds to the correlation obtained during KEOPS1 (Jacquet et al., 2015); (c) Dehairs's relationship between depth-weighted average (DWA) mesopelagic Baxs (pM) and JO2 (µmol L−1 d−1) compared to the ANTARES result (this study), KEOPS1 data (Southern Ocean; Jacquet et al., 2015) and GEOVIDE correlation (North Atlantic; Lemaitre et al., 2018).
In Fig. 3b we applied a similar approach as reported in Jacquet et al. (2015) where we show the correlation between JO2 obtained from dark community respiration DCR (Winkler titration, JO2-DCR) data integration in the water column and JO2 based on Baxs content (Dehairs's transfer function, later referred to as JO2-Ba). Similarly, to estimate JO2-Ba in the present study, we used the following equation (Dehairs et al., 1997) (Fig. 3c):
A Ba BKG value of 130 pM was used (see above). JO2-Ba is compared to JO2-Opt integrated over the same layer depth (between 175 and 450 m; Fig. 3b). JO2 rates are of the same order of magnitude (JO2-Ba = 4.59 mmol m−2 d−1 and JO2-opt = 3.14 mmol m−2 d−1). The slight difference could be explained by the integration time of both methods: a few hours to days for the incubations vs. a few days to weeks for Baxs (seasonal build-up; Jacquet et al., 2007). JO2 rates calculated in the present work are 3 times higher than those reported in the Southern Ocean during KEOPS1 (Jacquet et al., 2015), but they are in good agreement with the Baxs vs. JO2 trend (Fig. 3b). DWA Baxs and JO2 measured during KEOPS1 (Jacquet et al., 2015) and at the ANTARES site (this study) are compared to Dehairs's relationship in Fig. 3c. The correlation obtained in Lemaitre et al. (2018) in the North Atlantic is also reported (JO2 was calculated from apparent oxygen utilization divided by water mass age). Note that this relationship is not significantly different from the Dehairs equation (Lemaitre et al., 2018). Overall, results at the ANTARES site are in line with the Southern Ocean Baxs–JO2 correlation. This further supports the validity of the Dehairs transfer function in the present study.
3.4 Estimated particle remineralization rates and implications
In order to provide a Baxs-derived estimate of POC remineralization rate (MR) at the ANTARES/EMSO-LO observatory during the BATMAN cruise, we converted JO2-Ba into C respired using the Redfield (RR) C O2 molar ratio (; Broecker et al., 1985) multiplied by the depth layer considered (Z, 175–450 m) (Dehairs et al., 1997):
We obtain a POC remineralization rate of 11 mmol C m−2 d−1 (10 % RSD, relative standard deviation). This is within the range of dissolved Ba-derived fluxes of POC remineralization (13 to 29 mmol C m−2 d−1) reported in the Mediterranean Sea previously (Jacquet et al., 2016; Jullion et al., 2017). Following calculations reported in Jullion et al. (2017), our MR rate would correspond to a Baxs flux of around 0.01 µmol m−2 d−1. This is in reasonable agreement with barium fluxes (0.01 to 0.08 µmol m−2 d−1) presented in Jullion et al. (2017). Previously published barium fluxes from sediment traps range from 0.27 to 0.36 µmol m−2 d−1 at the DYFAMED station (Sternberg et al., 2007) and from 0.39 to 1.07 µmol m−2 d−1 in the Alboran Sea (Sanchez-Vidal et al., 2005). The POC remineralization rate from the present study is in the range of previously published carbon export fluxes (a few to tens of mmol m−2 d−1) from thorium-derived data (Speicher et al., 2006) or from combining drifting sediment traps and underwater vision profilers (Ramondenc et al., 2016). Constraining POC flux attenuation and remineralization rates in the Mediterranean is far from being achieved, especially regarding seasonal changes and inter-basin variations, but the concordance of the different approaches is promising.
This paper presents the first insight into the connections of Baxs, PHP and JO2 at the ANTARES/EMSO-LO observatory site in the northwestern Mediterranean Sea during the BATMAN (2016) cruise. Our results reveal a strong relationship between Baxs contents and measured JO2 rates. Also, DWA Baxs vs. column-integrated PHP and measured vs. Baxs-based JO2 relationships follow trends previously reported in the Southern Ocean where the Dehairs function was first established to estimate POC remineralization rate. Results from the present study would indicate that this function can also be applied in the Mediterranean basin provided that adequate Baxs background values are estimated. From a global climate perspective, the Baxs tool will help to better balance the MedSea water column C budget. It will contribute to gaining focus on the emerging picture of the C transfer efficiency (strength of the biological pump).
All data and metadata will be made available at the French INSU/CNRS LEFE CYBER database (scientific coordinator: Hervé Claustre; data manager, webmaster: Catherine Schmechtig) (INSU/CNRSLEFE CYBER, 2020).
SHMJ and DL designed the experiment for JO2. SHMJ, CT and MG designed the experiments for PHP measurements. SHMJ and FACLM managed barium sampling during the cruise. NB managed CTD deployment at sea. MG, SG and MR managed PHP. All co-authors contributed to writing.
The authors declare that they have no conflict of interest.
We thank the officers and crew of R/V L'EUROPE for their assistance during work at sea.
This research was supported by the French national LEFE/INSU “REPAP” project (PI is Stéphanie H. M. Jacquet). It was co-funded by the “ROBIN” project (PIs are Christian Tamburini, Frédéric A. C. Le Moigne) of Labex OT-Med (ANR-11-LABEX-0061) funded by the Investissements d'Avenir and the French Government project of the ANR, through the A*Midex project (ANR-11-IDEX-0001-02). The authors have benefited from the support of the SNO-MOOSE and SAM-MIO. BATMAN is a contribution to the “AT – POMPE BIOLOGIQUE” of the Mediterranean Institute of Oceanography (MIO) and to the international IMBER program. The instrument (ELEMENT XR, Thermo Fisher) was provided in 2012 by the European Regional Development Fund (ERDF).
This paper was edited by Carolin Löscher and reviewed by two anonymous referees.
Bertram, M. and Cowen, J.: Morphological and compositional evidence for biotic precipitation of marine barite, J. Mar. Res., 55, 577–593, 1997.
Broecker, W. S.: “NO”, a conservative water-mass tracer, Earth Planet. Sc. Lett., 23, 100–107, https://doi.org/10.1016/0012-821X(74)90036-3, 1974.
Broecker, W. S., Takahashi, T., and Takahashi, T.: Sources and flow patterns of deep-ocean waters as deduced from potential temperature, salinity and initial phosphate concentration, J. Geophys. Res., 90, 6925–6939, 1985.
Buesseler, K. O. and Boyd, P. W.: Shedding light on processes that control particle export and flux attenuation in the twilight zone of the open ocean, Limnol. Oceanogr., 54, 1210–1232, https://doi.org/10.4319/lo.2009.54.4.1210, 2009.
Cardinal, D., Dehairs, F., Cattaldo, T., and André, L.: Geochemistry of suspended particles in the Subantarctic and Polar Frontal zones south of Australia: Constraints on export and advection processes, J. Geophys. Res., 106, 31637–31656, https://doi.org/10.1029/2000JC000251, 2001.
Cardinal, D., Savoye, N., Trull, T. W., André, L., Kopczynska, E. E., and Dehairs, F.: Variations of carbon remineralisation in the Southern Ocean illustrated by the Baxs proxy, Deep-Sea Res. Pt. I, 52, 355–370, https://doi.org/10.1016/j.dsr.2004.10.002, 2005.
Dehairs, F., Lambert, C. E., Chesselet, R., and Risler, N.: The biological production of marine suspended barite and the barium cycle in the Western Mediterranean Sea, Biogeochemistry, 4, 119–140, https://doi.org/10.1007/BF02180151, 1987.
Dehairs, F., Shopova, D., Ober, S., Veth, C., and Goeyens, L.: Particulate barium stocks and oxygen consumption in the Southern Ocean mesopelagic water column during spring and early summer: relationship with export production, Deep-Sea Research Pt. II, 44, 497–516, https://doi.org/10.1016/S0967-0645(96)00072-0, 1997.
Dehairs, F., Jacquet, S., Savoye, N., Van Mooy, B. A. S., Buesseler, K. O., Bishop, J. K. B., Lamborg, C. H., Elskens, M., Baeyens, W., Boyd, P. W., Casciotti, K. L., and Monnin, C.: Barium in twilight zone suspended matter as a potential proxy for particulate organic carbon remineralization: Results for the North Pacific, Deep-Sea Res. Pt. II, 55, 1673–1683, https://doi.org/10.1016/j.dsr2.2008.04.020, 2008.
Ellison, S. L. R.: Eurachem/CITAC Guide CG4, Quantifying Uncertainty in Analytical Measurement, 2nd Edn., edited by: Ellison, S. L. R., Rosslein, M., and Williams, A., 120 pp., ISBN 0948926 15 5, 2000.
Giering, S. L. C., Sanders, R., Lampitt, R. S., Anderson, T. R., Tamburini, C., Boutrif, M., Zubkov, M. V., Marsay, C. M., Henson, S. A., Saw, K., Cook, K., and Mayor, D. J.: Reconciliation of the carbon budget in the ocean/'s twilight zone, Nature, 507, 480–483, 2014.
Hainbucher, D., Rubino, A., Cardin, V., Tanhua, T., Schroeder, K., and Bensi, M.: Hydrographic situation during cruise M84/3 and P414 (spring 2011) in the Mediterranean Sea, Ocean Sci., 10, 669–682, https://doi.org/10.5194/os-10-669-2014, 2014.
Henson, S. A., Sanders, R., Madsen, E., Morris, P. J., Le Moigne, F., and Quartly, G. D.: A reduced estimate of the strength of the ocean's biological carbon pump, Geophys. Res. Lett., 38, L04606, https://doi.org/10.1029/2011GL046735, 2011.
IPCC: 5th Assessment Report (AR5) Climate Change 2013, Working Group 1, January 2014.
Jacquet, S. H. M., Henjes, J., Dehairs, F., Worobiec, A., Savoye, N., and Cardinal, D.: Particulate Ba-barite and acantharians in the Southern Ocean during the European Iron Fertilization Experiment (EIFEX), J. Geophys. Res., 112, G04006, https://doi.org/10.1029/2006JG000394, 2007.
Jacquet, S. H. M., Dehairs, F., Savoye, N., Obernosterer, I., Christaki, U., Monnin, C., and Cardinal, D.: Mesopelagic organic carbon remineralization in the Kerguelen Plateau region tracked by biogenic particulate Ba, Deep-Sea Res. Pt. II, 55, 868–879, https://doi.org/10.1016/j.dsr2.2007.12.038, 2008a.
Jacquet, S. H. M., Savoye, N., Dehairs, F., Strass, V. H., and Cardinal, D.: Mesopelagic carbon remineralization during the European Iron Fertilization Experiment, Global Biogeochem. Cy., 22, GB1023, https://doi.org/10.1029/2006GB002902, 2008b.
Jacquet, S. H. M., Dehairs, F., Dumont, I., Becquevort, S., Cavagna, A.-J., and Cardinal, D.: Twilight zone organic carbon remineralization in the Polar Front Zone and Subantarctic Zone south of Tasmania, Deep-Sea Res. Pt. II, 58, 2222–2234, https://doi.org/10.1016/j.dsr2.2011.05.029, 2011.
Jacquet, S. H. M., Dehairs, F., Lefèvre, D., Cavagna, A. J., Planchon, F., Christaki, U., Monin, L., André, L., Closset, I., and Cardinal, D.: Early spring mesopelagic carbon remineralization and transfer efficiency in the naturally iron-fertilized Kerguelen area, Biogeosciences, 12, 1713–1731, https://doi.org/10.5194/bg-12-1713-2015, 2015.
Jacquet, S. H. M., Monnin, C., Riou, V., Jullion, L., and Tanhua, T.: A high resolution and quasi-zonal transect of dissolved Ba in the Mediterranean Sea, Mar. Chem., 178, 1–7, https://doi.org/10.1016/j.marchem.2015.12.001, 2016.
Jullion, L., Jacquet, S. H. M., and Tanhua, T.: Untangling biogeochemical processes from the impact of ocean circulation: First insight on the Mediterranean dissolved barium dynamics, Global Biogeochem. Cy., 31, 1256–1270, https://doi.org/10.1002/2016GB005489, 2017.
Kirchman, D. L.: Leucine incorporation as a measure of biomass production by heterotrophic bacteria, in: Handbooks of methods in aquatic microbial ecology,, edited by: Kemp, P. F., Sherr, B. F., Sherr, E. B., and Cole, J. J., Lewis Publishers, Boca Raton, Ann Arbor, London, Tokyo, 509–512, 1993.
Lemaitre, N., Planquette, H., Planchon, F., Sarthou, G., Jacquet, S., García-Ibáñez, M. I., Gourain, A., Cheize, M., Monin, L., André, L., Laha, P., Terryn, H., and Dehairs, F.: Particulate barium tracing of significant mesopelagic carbon remineralisation in the North Atlantic, Biogeosciences, 15, 2289–2307, https://doi.org/10.5194/bg-15-2289-2018, 2018.
Malanotte-Rizzoli, P., Artale, V., Borzelli-Eusebi, G. L., Brenner, S., Crise, A., Gacic, M., Kress, N., Marullo, S., Ribera d'Alcalà, M., Sofianos, S., Tanhua, T., Theocharis, A., Alvarez, M., Ashkenazy, Y., Bergamasco, A., Cardin, V., Carniel, S., Civitarese, G., D'Ortenzio, F., Font, J., Garcia-Ladona, E., Garcia-Lafuente, J. M., Gogou, A., Gregoire, M., Hainbucher, D., Kontoyannis, H., Kovacevic, V., Kraskapoulou, E., Kroskos, G., Incarbona, A., Mazzocchi, M. G., Orlic, M., Ozsoy, E., Pascual, A., Poulain, P.-M., Roether, W., Rubino, A., Schroeder, K., Siokou-Frangou, J., Souvermezoglou, E., Sprovieri, M., Tintoré, J., and Triantafyllou, G.: Physical forcing and physical/biochemical variability of the Mediterranean Sea: a review of unresolved issues and directions for future research, Ocean Sci., 10, 281–322, https://doi.org/10.5194/os-10-281-2014, 2014.
Martin, J. H., Knauer, G. A., Karl, D. M., and Broenkow, W. W.: VERTEX: carbon cycling in the northeast Pacific, Deep-Sea Res., 34, 267–285, https://doi.org/10.1016/0198-0149(87)90086-0, 1987.
Martinez-Ruiz, F., Jroundi, F., Paytan, A., Guerra-Tschuschke, I., del Abad, M. M., and González-Muñoz, M. T.: Barium bioaccumulation by bacterial biofilms and implications for Ba cycling and use of Ba proxies, Nat. Commun., 9, 1619, https://doi.org/10.1038/s41467-018-04069-z, 2018.
Martinez-Ruiz, F., Paytan, A., Gonzalez-Muñoz, M. T., Jroundi, F., Abad, M. M., Lam, P. J., Bishop, J. K. B., Horner, T. J., Morton, P. L., and Kastner, M.: Barite formation in the ocean: Origin of amorphous and crystalline precipitates, Chem. Geol., 511, 441–451, https://doi.org/10.1016/j.chemgeo.2018.09.011, 2019.
Ramondenc, S., Madeleine, G., Lombard, F., Santinelli, C., Stemmann, L., Gorsky, G., and Guidi, L.: An initial carbon export assessment in the Mediterranean Sea based on drifting sediment traps and the Underwater Vision Profiler data sets, Deep-Sea Res. Pt. I, 117, 107–119, https://doi.org/10.1016/j.dsr.2016.08.015, 2016.
Sanchez-Vidal, A., Collier, R. W., Calafat, A., Fabres, J., and Canals, M.: Particulate barium fluxes on the continental margin: a study from the Alboran Sea (Western Mediterranean), Mar. Chem., 93, 105–117, https://doi.org/10.1016/j.marchem.2004.07.004, 2005.
Santinelli, C., Nannicini, L., and Seritti, A.: DOC dynamics in the meso and bathypelagic layers of the Mediterranean Sea, Deep-Sea Res. Pt. II, 57, 1446–1459, https://doi.org/10.1016/j.dsr2.2010.02.014, 2010.
Schlitzer, R.: Ocean Data View, GEO/ODV, available at: http://www.awi-bremerhaven.de/ (last access: 2 March 2021), 2002.
Shopova, D., Dehairs, F., and Baeyens, W.: A simple model of biogeochemical element distribution in the oceanic water column, J. Mar. Syst., 6, 331–344, https://doi.org/10.1016/0924-7963(94)00032-7, 1995.
Siegel, D. A., Buesseler, K. O., Behrenfeld, M. J., Benitez-Nelson, C. R., Boss, E., Brzezinski, M. A., Burd, A., Carlson, C. A., D'Asaro, E. A., Doney, S. C., Perry, M. J., Stanley, R. H. R., and Steinberg, D. K.: Prediction of the Export and Fate of Global Ocean Net Primary Production: The EXPORTS Science Plan, Front. Mar. Sci., 3, 22, https://doi.org/10.3389/fmars.2016.00022, 2016.
Simon, M. and Azam, F.: Protein content and protein synthesis rates of planktonic marine bacteria, Mar. Ecol. Prog. Ser., 51, 201–213, 1989.
Speicher, E. A., Moran, S. B., Burd, A. B., Delfanti, R., Kaberi, H., Kelly, R. P., Papucci, C., Smith, J. N., Stavrakakis, S., Torricelli, L., and Zervakis, V.: Particulate organic carbon export fluxes and size-fractionated POC 234Th ratios in the Ligurian, Tyrrhenian and Aegean Seas, Deep-Sea Res. Pt. I, 53, 1810–1830, https://doi.org/10.1016/j.dsr.2006.08.005, 2006.
Sternberg, E., Jeandel, C., Miquel, J.-C., Gasser, B., Souhaut, M., Arraes-Mescoff, R., and Francois, R.: Particulate barium fluxes and export production in the northwestern Mediterranean, Mar. Chem., 105, 281–295, https://doi.org/10.1016/j.marchem.2007.03.003, 2007.
Sternberg, E., Jeandel, C., Robin, E., and Souhaut, M.: Seasonal cycle of suspended barite in the mediterranean sea, Geochim. Cosmochim. Ac., 72, 4020–4034, https://doi.org/10.1016/j.gca.2008.05.043, 2008.
Tamburini, C., Garcin, J., Ragot, M., and Bianchi, A.: Biopolymer hydrolysis and bacterial production under ambient hydrostatic pressure through a 2000 m water column in the NW Mediterranean, Deep-Sea Res. Pt. I, 49, 2109–2123, https://doi.org/10.1016/S0967-0645(02)00030-9, 2002.
Tamburini, C., Boutrif, M., Garel, M., Colwell, R. R., and Deming, J. W.: Prokaryotic responses to hydrostatic pressure in the ocean – a review, Environ. Microbiol., 15, 1262–1274, https://doi.org/10.1111/1462-2920.12084, 2013.
Taylor, S. R. and McLennan, S. M.: The continental crust: its composition and evolution, Blackwell Scientific Publications, USA, 312 pp., 1985.