the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 06 Jun 2025
| 06 Jun 2025
Reconstructing central African hydroclimate over the past century using freshwater bivalve shell geochemistry
Zita Kelemen
David P. Gillikin
Steven Bouillon
Centennial records of climatic and hydrological data are scarce on the African continent. Freshwater bivalve shells have the potential to record climate-related proxies, from which reconstructions of river discharge (Q) or water isotope variations over long time periods would be possible. The Oubangui River, the largest right-bank tributary of the Congo River, is one of the few African rivers for which long-term Q records are available. This, together with the availability of museum-archived shells, makes it an ideal location to study changes in hydroclimate in central Africa over the past century and to validate the potential of proxies recorded in freshwater shells. We analysed the carbon and oxygen isotope composition (δ13C, δ18O) across the growth axis of museum-archived (collected between 1891 and ∼1952) and contemporary (live-collected in 2011 and 2013) Chambardia wissmanni shells from the Oubangui River, covering sections of the past ∼130 years. Shell isotopes show a clear baseline shift. Both δ13C and δ18O exhibit pronounced cyclicity in recent and historical shells, but recent shells showed a much wider range in δ18O values and a narrower range in δ13C values compared to historical specimens. In the Oubangui River, high Q is represented as low water δ18O (δ18Ow) values, while low Q is represented as high δ18Ow values, and these correlations are expected to be expressed in shell δ18O (δ18Oshell) values similarly. The historical δ18Oshell values covered only the lower part of the range measured in recent shells, which suggests a major change in the low-flow conditions of the Oubangui River between the 1950s and 2010s. Reconstructed Q values, based on the logarithmic relationship established between recent water δ18O values and measured Q, tended to overestimate the low flow values in the past, suggesting a different δ18Ow–Q relationship in the earlier part of the 20th century. Thus, while the freshwater shell δ18O patterns are consistent with the documented long-term Q changes in the Oubangui, the shells show that the most pronounced changes in hydroclimate over the past century are expressed in low-flow sections of the hydrograph and likely result from a combination of changes in the rainfall regime, aquifer recharge, and/or vegetation cover in the upper catchment. These results are consistent with the suggestion that dry periods in the upper Congo basin have become more extreme in recent times and highlight that freshwater shells could offer a valuable archive to study recent changes in catchments where no long-term empirical hydrological or climatological data are available.
- Article
(5655 KB) - Full-text XML
-
Supplement
(362 KB) - BibTeX
- EndNote




The Oubangui River is the largest right-bank tributary of the Congo River and represents one of the few sites within the Congo basin where discharge (Q) has been monitored since the early 20th century. Several studies have demonstrated that over the past century, the annual average Q at Bangui has decreased by 40 %, while the dry-season flow has decreased more drastically, by more than 60 % (Wesselink et al., 1996; Orange et al., 1997; Laraque et al., 2001; Runge and Nguimalet, 2005). Given the importance of river navigability for a landlocked country such as the Central African Republic, this decrease in Q has important ramifications for transport: while the river was navigable for larger boats for 8 months of the year before the 1970s, this is currently limited to 4–5 months (Van Pul, 2023). Despite these substantial and well-documented changes, rainfall in the watershed of the Oubangui River has decreased by only 5 % since 1970 (based on 11 stations in the central Oubangui catchment; Nguimalet and Orange, 2015; Laraque et al., 2020).
Anthropogenic changes within a catchment, such as forest clearing, land-use change, or water abstraction for irrigation, have often been suggested as driving forces of hydrological changes (Descroix et al., 2012; Recha et al., 2012; Tshimanga and Hughes, 2012; Amogu et al., 2014; Essaid and Caldwell, 2017). There are signs that the entire Congo River basin has been experiencing a drying trend over the past decades, especially the northern part of the forest (Asefi-Najafabady and Saatchi, 2013; Zhou et al., 2014; Malhi and Wright, 2004). However, land use in the Oubangui watershed upstream of Bangui has changed relatively little over the past century according to Nguimalet and Orange (2013), and it is therefore currently not considered to play a major role in explaining the decreased Q.
Longer-term records of climatic and hydrological data are relatively scarce for the African continent: the data collection network in the Congo basin is very sparse and few hydrological stations are still operational, with only a handful of sites for which long-term data (i.e. spanning the past century) are available (Laraque et al., 2001; Alsdorf et al., 2016). The application of proxies in biological archives therefore has high potential to reconstruct environmental records over the last few decades and centuries. There is an increasing set of studies from various lacustrine environments (Barker et al., 2011; Berke et al., 2012; Mologni et al., 2024) and tree cores (Battipaglia et al., 2015; Van Der Sleen et al., 2015), but freshwater bivalve shells have so far been rarely used on this continent (Abell et al., 1996; Kelemen et al., 2017, 2021). However, a number of studies in temperate regions have successfully used freshwater bivalve shells to shed light on past changes in freshwater ecosystems (Dettman and Lohmann, 2000; Dettman et al., 1999, 2001, 2004; Pfister et al., 2018, 2019; Schöne et al., 2020; Versteegh et al., 2010, 2011).
Bivalve shells are considered to be useful biological archives to study sub-seasonal changes in aquatic systems (reviewed in Stringer and Prendergast, 2023). Their abundance, filter-feeding lifestyle, and sequentially secreted carbonate shell make them ideal archives for reconstructing water isotope ratios, as confirmed by Pfister et al. (2019), although the latter review focused on average values and did not focus on higher-resolution (temporal) data. However, over different timescales, freshwater bivalves have been shown to record changes in Q and precipitation (Ricken et al., 2003; Dettman et al., 2004; Kaandorp et al., 2005; Versteegh et al., 2011; Kelemen et al., 2017, 2021), as they precipitate their shell carbonate in oxygen isotopic equilibrium with the host water (Grossman and Ku, 1986; Dettman et al., 1999; Goodwin et al., 2019).
With the exception of Margaritifera species with a reported maximum lifespan of 200 years or more (see Stringer and Prendergast, 2023), most freshwater bivalve species have a relatively short lifetime (∼1 decade), which limits the period of environmental data recorded by individual specimens. Sampling the sequentially secreted layers at high spatial resolution can reveal detailed information across several hydrological cycles. We have demonstrated earlier that the bivalve Chambardia wissmanni precipitates its aragonitic shell in oxygen isotope equilibrium with the host water in the Oubangui and Niger rivers (Kelemen et al., 2017). Seasonal temperature variations in such tropical rivers are relatively limited, and thus, oxygen isotope ratios in bivalve shells (δ18Oshell) are predominantly influenced by the oxygen isotope ratios of water (δ18Ow) (Kelemen et al., 2017), which in turn are predominantly controlled by rainfall patterns and evaporation (Gat, 1996). Likewise, shell δ13C values may track the δ13C of the dissolved inorganic carbon (DIC) in the host water (e.g. Graniero et al., 2017), and a relationship between Q and δ13CDIC values has been noted in tropical rivers (Bouillon et al., 2012, 2014).
In this study, we aimed at reconstructing aspects of the hydroclimate of central Africa using high-resolution stable isotope analyses (δ13C and δ18O) on both recent and archived freshwater shells from the Oubangui River. The choice of the Oubangui River stems from the fact that (i) we have previously calibrated isotope proxies in bivalve shells from this river by comparing shell data with in situ water data collected during the shell growth period (Kelemen et al., 2017); (ii) shell specimens were available from museum collections that spanned a wide period from the late 19th century to mid-20th century; and (iii) clear changes in Q of the Oubangui have been documented over part of the period covered by our shell collection, providing an independent verification of the applicability of our approach.
Moreover, regular sampling of river water over several years has shown that a clear inverse relationship exists between Q and δ18Ow values (Kelemen et al., 2017). Thus, we predicted that historical specimens (pre-1960) would (i) record the documented long-term decrease in dry-season and annual Q, (ii) show a less pronounced amplitude in δ18Oshell values, and (iii) more specifically lack the high δ18Oshell values associated with low-Q conditions characteristic of the most recent decades (as shown by Kelemen et al., 2017).
2.1 Oubangui basin
The Oubangui River (Fig. 1) is the largest right-bank tributary of the Congo River, with an average annual Q measured at Bangui (Central African Republic) of 3700 m3 s−1 (for the period of 1935–2015; Nguimalet and Orange, 2015) and a length of 2400 km including its upper reaches, the Uele River (Boulvert, 1987). Along its path to the Congo River, the Oubangui mainly passes through peneplain landscapes, relatively flat open landscapes in the north, and mosaics of forest–savannah in its central basin, and the landscape transforms into dense humid rainforest along its southern reaches (Mayaux et al., 1999). Influenced by the seasonal migration of the equatorial (tropical) rain belt, the highest rainfall is during the July–October period in the central part of the basin, while closer to the Equator, the rainfall peak is bimodal, with the highest peak during September to December and March to May (Mahé, 1993). The central Oubangui River watershed is mostly within the Central African Republic (CAR), as in this part of the basin tributaries join exclusively from the right bank. The general water recharge in the central Oubangui is supplied by abundant rainfall north of the main stem during the wet season. In the CAR, numerous rain-gauging stations have been established in the past century; however few stations have continuous (multi-decadal) measurements available, and even fewer have data after the 1980s (data can be found in the SIEREM database, provided by the HydroSciences Montpellier Laboratory, at http://www.hydrosciences.fr/sierem/index_en.htm, last access: 1 July 2024). Continuous precipitation isotope measurements are only available for two stations at Bangui, for the period 2009–2021, provided by IAEA (Global Network of Isotopes in Precipitation (GNIP; available via https://nucleus.iaea.org/sites/ihn/Pages/GNIP.aspx, last access: 1 July 2024)). The single-peak discharge period at Bangui results from a transitional tropical regime of the basin, with one wet and one dry season, and annual precipitation of 1475 mm (average over the period between 1935 and 2015 at Bangui by Nguimalet and Orange, 2019). The highest Q is usually measured between September and November, when the rainy season peak arrives from the upper catchment, and water levels are the lowest after the dry season between February and April. The gauging station at Bangui is one of the few sites within the Congo basin for which a long-term Q record is available (Orange et al., 1997; Wesselink et al., 1996; Laraque et al., 2001; Runge and Nguimalet, 2005), and this station provides Q data from a catchment area of about 490 000 km2 upstream of Bangui. The discharge monitoring station was established in 1911 at Bangui, but a few years of intermittent records follow, leaving a relatively long data gap between 1920 and 1935, after which continuous daily measurements are available. Discharge data were obtained from the Direction de la Météorologie Nationale (CAR).
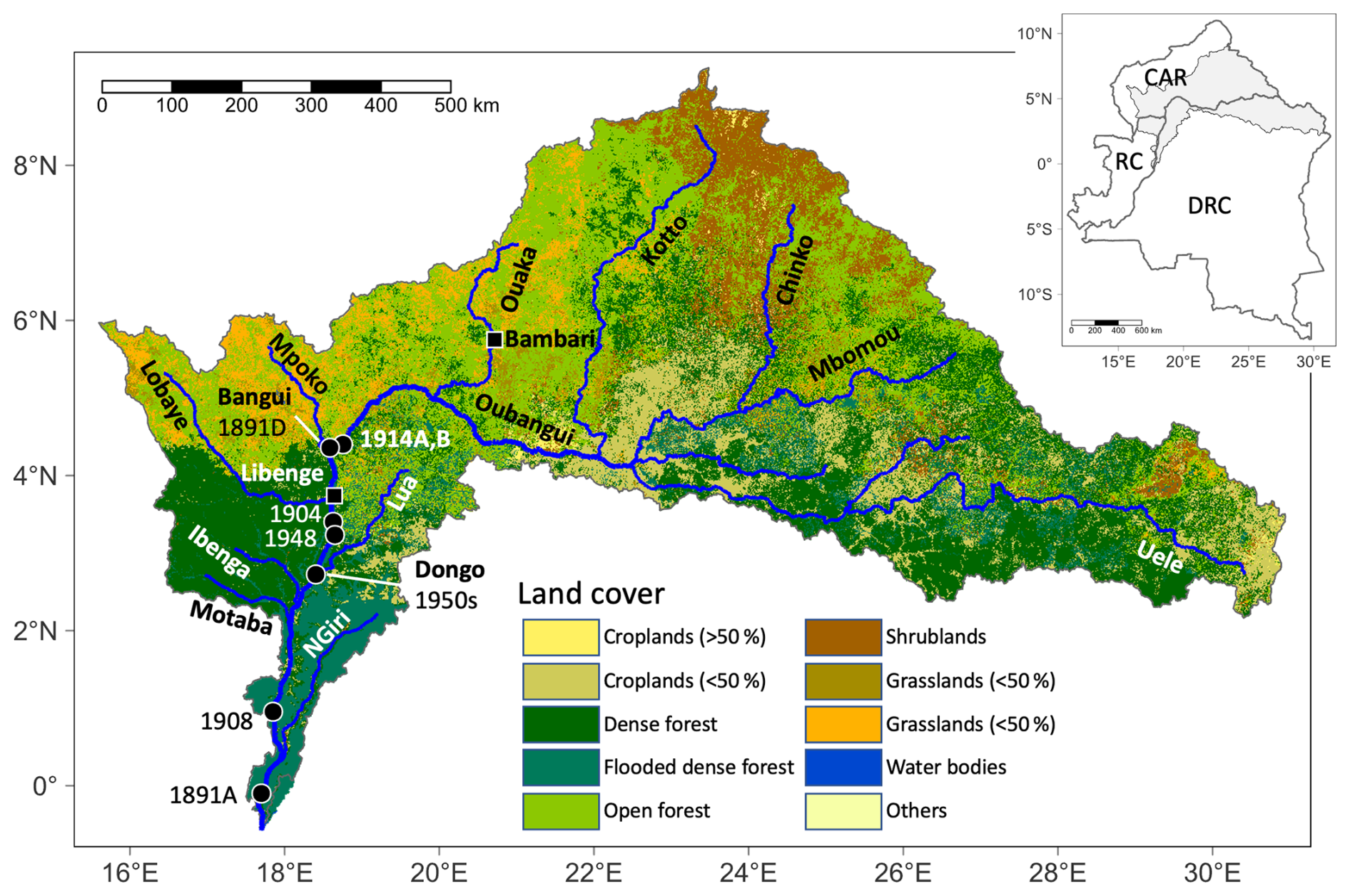
Figure 1Land cover of the Oubangui basin and the larger tributaries upstream of most downstream shell collection. Black circles represent the collection sites of the museum-archived specimens with the collection year labelled. Black squares represent cities (unless also a collection site). The inset map shows the position of the Oubangui watershed in the three countries it covers (CAR denotes the Central African Republic, DRC denotes the Democratic Republic of the Congo, and RC denotes the Republic of the Congo).
2.2 Shell collection and analyses
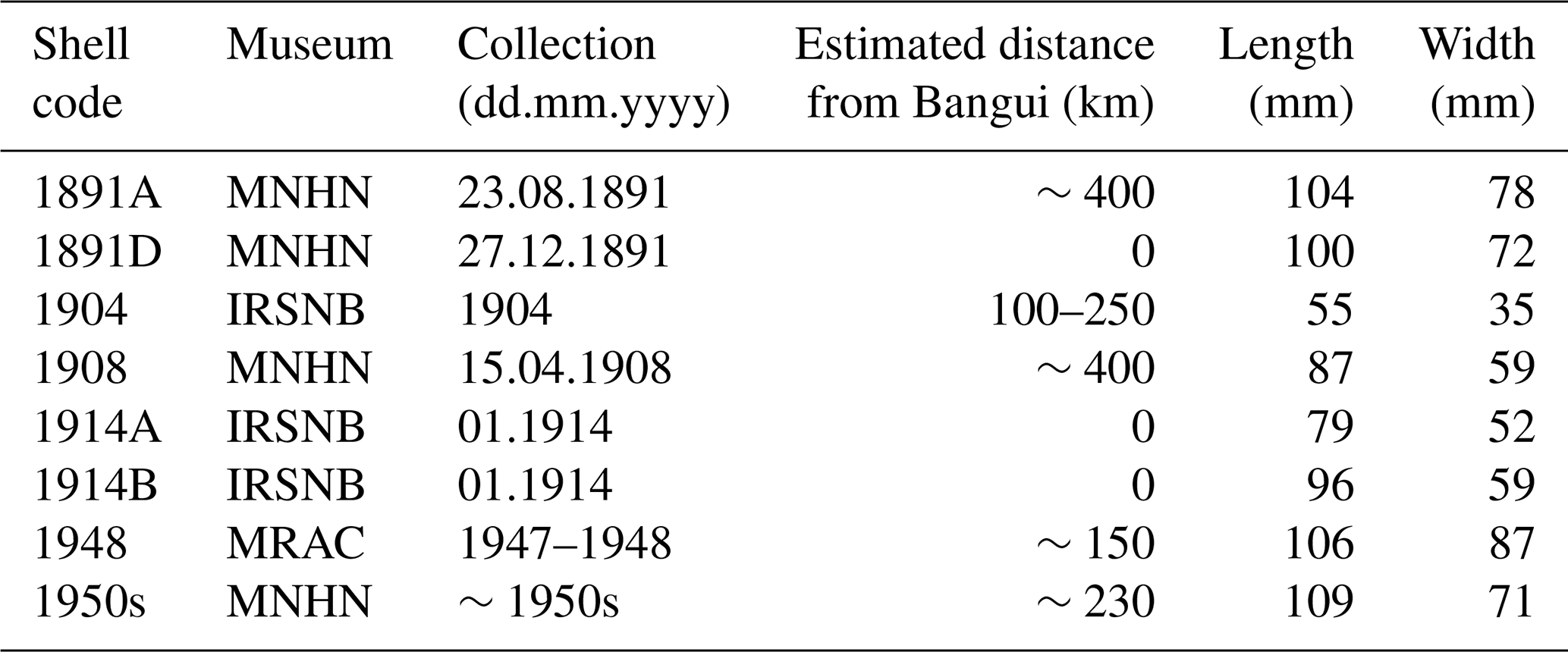
Recent specimens of C. wissmanni were collected in the Oubangui River in Bangui between 2011 and 2014 (see Kelemen et al., 2017). We searched museum collections (initially from the MusselP database; http://mussel-project.uwsp.edu, last access: 15 February 2020) for specimens of C. wissmanni that could unambiguously be assigned as originating from the Oubangui River and that included sufficient information on the collection date or period. A total of eight shells (see Table 1 for details) could be sourced from various museums (MRAC – Royal Museum for Central Africa, Tervuren, Belgium; IRSNB – Royal Belgian Institute of Natural Sciences, Brussels, Belgium; and MNHN – Muséum National d'Histoire Naturelle, Paris, France). Two C. wissmanni specimens (labelled as Spatha rubens) were collected in 1891 by Jean Dybowski, based on the mission reports one of these shells (1891A – August 1891) was collected a few kilometres above the confluence of Oubangui and Congo rivers, while the specimen from December 1891 (1891D) was collected at Bangui. One C. wissmanni shell from 1904 mentions that it was collected between 100 and 250 km downstream of Bangui, somewhere between Libenge and Dongo (Fig. 1). The 1908 specimen (labelled as Spathopsis wissmanni) was collected ∼500 km downstream of Bangui at Longo during the Robert Hottot mission. Two shells sampled in 1914 (labelled as Spatha oubanghi) are from Bangui (1914A, 1914B). Two bivalves were collected downstream of Bangui, near Mawuya (Aspatharia) and Dongo (C. wissmanni). The specimen from Mawuya is part of a larger, carefully preserved collection (different species, all valves paired and in excellent condition, each pair bound together with string); thus even though we could not find the exact date of sampling, we strongly expect late 1947 or early 1948 to be the collection period, as two large fauna collection campaigns were held in Mawuya by the Royal Belgian Institute for Natural History – in October 1947 and in January–February 1948. C. wissmanni from Dongo was a part of a collection of a known collector, R. H. Raemackers; this specimen entered the collection on 9 November 1957, yet the exact date of collection is not provided – we therefore considered the shell likely to have been from the 1950s. The shell collection was reviewed by Graf and Cummings (2011), and all specimens we used were confirmed to be C. wissmanni.
Table 1Data from all analysed C. wissmanni valves including sample code, estimated downstream distance from Bangui, collection date and collection holder, and shell size.
Shells were sectioned along the maximal growth axis, and sections of a few millimetres in thickness were mounted on glass slides. The prismatic layer was sampled from the tip towards the umbo using a New Wave MicroMill equipped with 300 µm diameter drill bit. The sampling resolution was between 350–750 and 300–450 µm depth, resulting in 50–80 µg powder for analysis. Samples were collected in 12 mL Labco Exetainers, flushed with helium and reacted with>100 % phosphoric acid. The evolved carbon dioxide was then analysed on a Thermo Delta V Advantage isotope ratio mass spectrometer coupled to a GasBench II, at either Union College (NY, USA) or KU Leuven (Belgium). Along with the samples, certified reference materials (LSVEC, NBS-18, NBS-19) and in-house CaCO3 standards were analysed, and the long-term standard deviations were better than 0.1 ‰ for both δ13C and δ18O.
To calculate reconstructed δ18Ow values, the 1000ln (α) equation of Dettman et al. (1999) (based on data from Grossman and Ku, 1986) was used (Eqs. 1 and 2). First the aragonite–water fractionation factor (α) was determined (Eq. 1):
where T is the water temperature in kelvins. Subsequently, based on the fractionation between water and aragonite described by
δ18Ow values were calculated including the conversion factor from the VSMOW to the VPDB scale of 1.0309 (Gonfiantini et al., 1995).
For further calculations, an average temperature of 28.6 °C (derived from in situ measurements over 3 years of monitoring; see Kelemen et al., 2017) was used, assuming only a minimal water temperature change over the past century and based on the measured centennial air temperature increase in the 20th century of about 0.5 °C on the African continent (Hulme et al., 2001).
Chambardia wissmanni is known to precipitate aragonite shells (all unionids make aragonite shells; Strayer and Malcom 2007). Experimental studies (e.g. Casella et al., 2017) have shown that metastable aragonite does not transform into a more stable structure of calcite below 175 °C, especially over the time frames considered here. Fossil freshwater bivalves (Diplodon longulus) collected from the Miocene Pebas Formation in Amazonia showed no evidence of diagenetic alteration (Kaandorp et al., 2005; Vonhof et al., 1998, 2003) and nor did 66 Ma (Cretaceous) freshwater shells from the Hell Creek formation (Dettman and Lohmann, 2000; Tobin et al., 2014); thus we can be confident that the aragonite structure of our collection stored for ∼100 years at ambient conditions is unaltered.
The full set of δ13C and δ18O data on historical shells is provided in the Supplement. Data on recent shells (Kelemen et al., 2017) are publicly accessible via ResearchGate (https://www.researchgate.net/publication/331716269, last access: 3 June 2025).
The δ18Oshell cyclicity in the historical specimens was well defined, with values ranging between −5.0 and −1.4 ‰ and individual shell amplitudes (difference between maximum and minimum values) ranging between 1.6‰ and 2.7‰ (Figs. 2 and 3). In recent shells, δ18Oshell values ranged between −5.1 ‰ and +0.2 ‰, with amplitudes up to 5.0 ‰, considerably higher than the historical specimens. With only a few exceptions, the δ18Oshell data in historical specimens remained below −2 ‰, whereas the modern shells were often above −1 ‰ (Fig. 3). In the collection of historical specimens, there was no considerable difference in the δ18Oshell record between the bivalves collected at Bangui and those collected somewhat more downstream (Fig. 2).
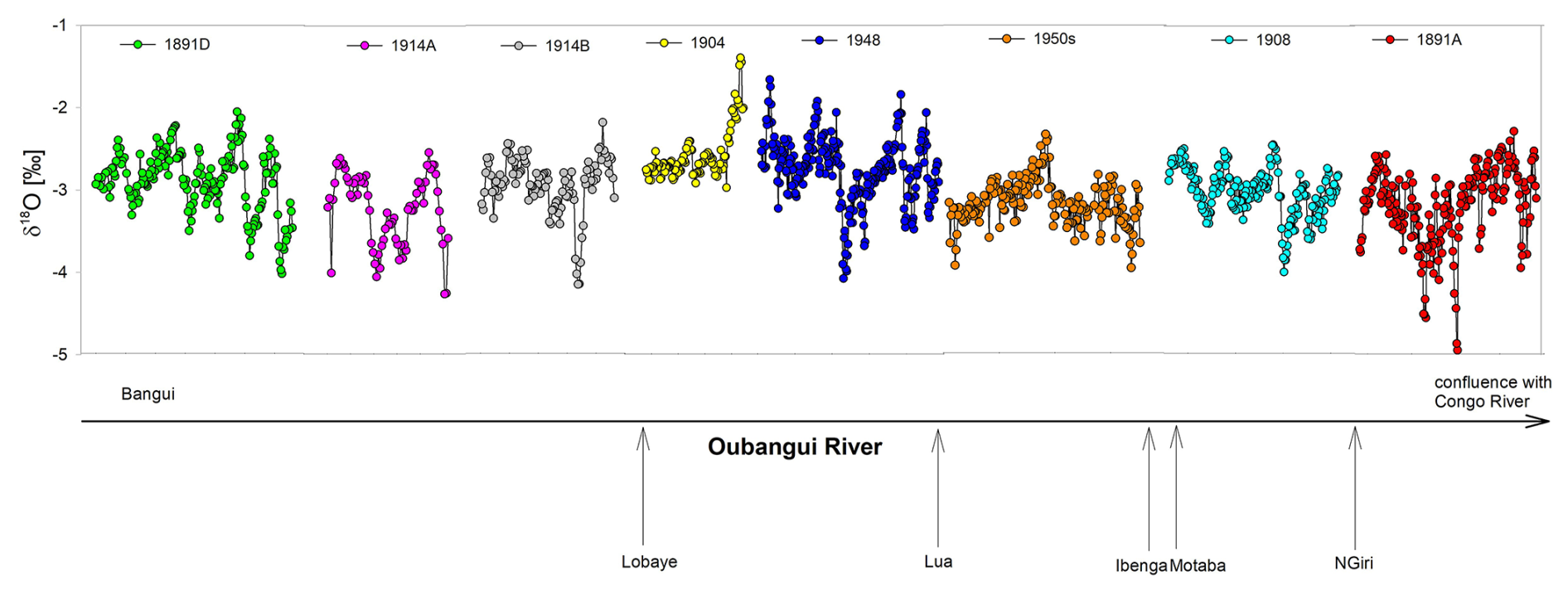
Figure 2δ18O values of all analysed historical specimens in order of collection site, from Bangui towards the confluence with the Congo River. Arrows on the x axis show the locations where important tributaries join the Oubangui River. The year of shell collection is mentioned for each specimen in the symbol legend. See Table 1 for the actual distance downstream of Bangui.
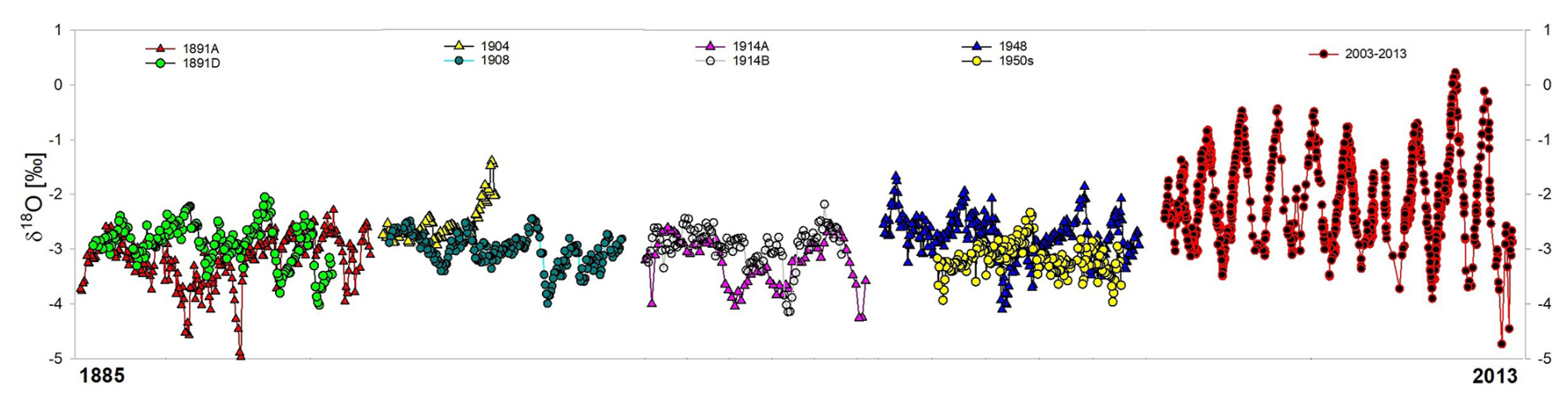
Figure 3δ18Oshell values of all analysed archived and recently collected specimens, starting from the oldest archived specimens (collected in 1891) and going towards the most recently collected shells, where nine specimens are combined into one composite record (recent shell data from Kelemen et al., 2017).
δ13Cshell values in historical specimens ranged between −15.7 ‰ and −7.0 ‰ and show clear cyclicity (Fig. 4). They were non-synchronous with δ18Oshell cycles, but the patterns observed in the older shells were analogous with the recently collected specimens (Kelemen et al., 2017). The δ13Cshell range was slightly wider than that in modern specimens (from −14.6 ‰ to −8.2 ‰; Kelemen et al., 2017). With one exception (a young specimen from 1904), all shells expressed an ontogenetic decrease in δ13Cshell values (Fig. 4; but note that the modern shells are a compilation of many shells, while the older shells represent individuals).
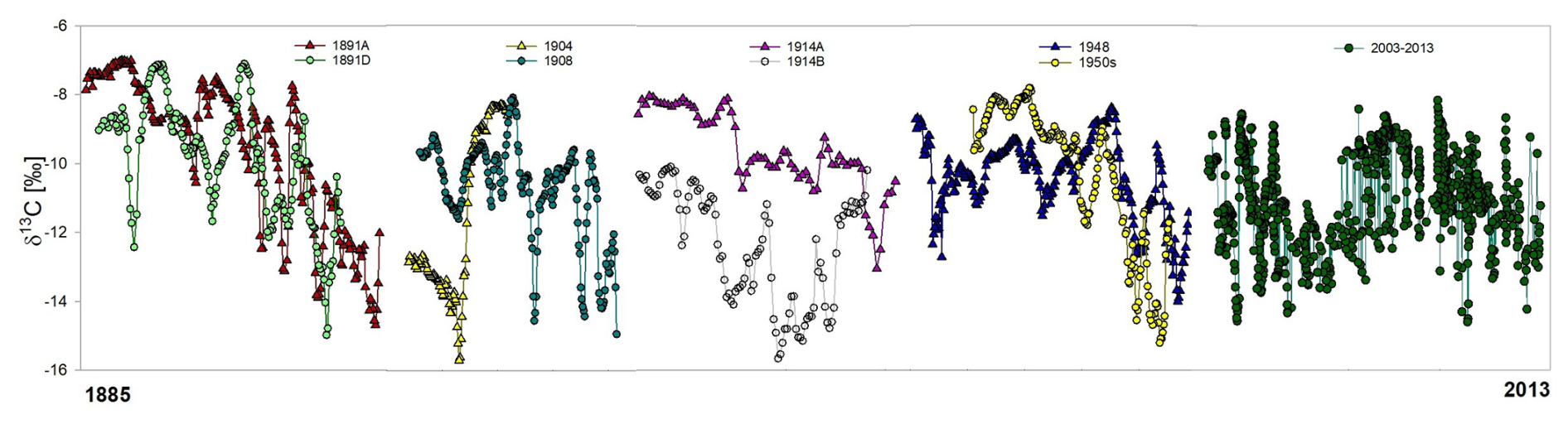
Figure 4δ13Cshell values of bivalve shells, starting from the oldest archived specimens (collected in 1891) and going towards the most recently collected shells; recent shells are sorted chronologically into one composite record (recent shell data from Kelemen et al., 2017).
Freshwater bivalve shells spanning the past ∼130 years show a major hydrological shift in the northern Congo basin over this time frame. From approximately the 1880s to the 1950s, shells show a fairly uniform range of δ18O values, but shells collected after 2003 show a marked increase in the upper range of δ18O values, suggesting a major shift in the hydroclimate to dry seasons with less precipitation. Unionid bivalves precipitate shell material in oxygen isotope equilibrium with the surrounding water, thus reflecting δ18O values of the water and temperature (Dettman et al., 1999; Versteegh et al., 2011; Kelemen et al., 2017; Goodwin et al., 2019), but in these tropical specimens, temperature plays a minor role, as the temperature variations throughout the year are relatively low (28.6±1.2 °C for continuous measurements between 2010–2013; discussed in Kelemen et al., 2017). Thus, the difference between the historical and recent bivalves clearly reflects a difference in δ18Ow between the early 20th–mid-20th century and those of the past decade (i.e. 2005–2013; see Fig. 3). This matches the expectation that shells recorded the different discharge conditions from the early 20th century, but a robust interpretation requires a more detailed look at the data and assumptions. Therefore, in the following discussion we will first briefly describe the most important characteristics of the Oubangui River, then subsequently attempt to reconstruct Q using the shell δ18O record and discuss whether a change in the δ18O–Q relationship might have occurred over time. We then explore the possible cause(s) of the lower δ18O values of historical shells in comparison with the recently collected specimens in the context of shell collection sites, rainfall anomalies, and land-use and vegetation changes.
4.1 The Oubangui River discharge (Q)
The Oubangui River has a single-peak discharge regime, and the century-long Q record showed a stable phase from the beginning of the record until the early 1960s, when a decade of wetter conditions was experienced in central and west Africa, which was followed by a hydrological deficit over the entire basin (Orange et al., 1997; Laraque et al., 2001, 2013, 2020). A 100-year flood was measured in October 1916, when a Q of 14 000 m3 s−1 was recorded, and the lowest Q was measured in April 1990 (266 m3 s−1) (Fig. 5). In the centennial Q record, a difference can be seen between the flow in the early 20th–mid-20th century, when the lowest Q was generally not below 700 m3 s−1 (with three exceptional years: 1914, 1945, and 1946), while after the 1970s the low Q gradually decreased, regularly reaching values between 250–500 m3 s−1 during the dry season. However, the decrease in peak Q is also substantial over this time period (pre-1970 , recent ) (Fig. 5).
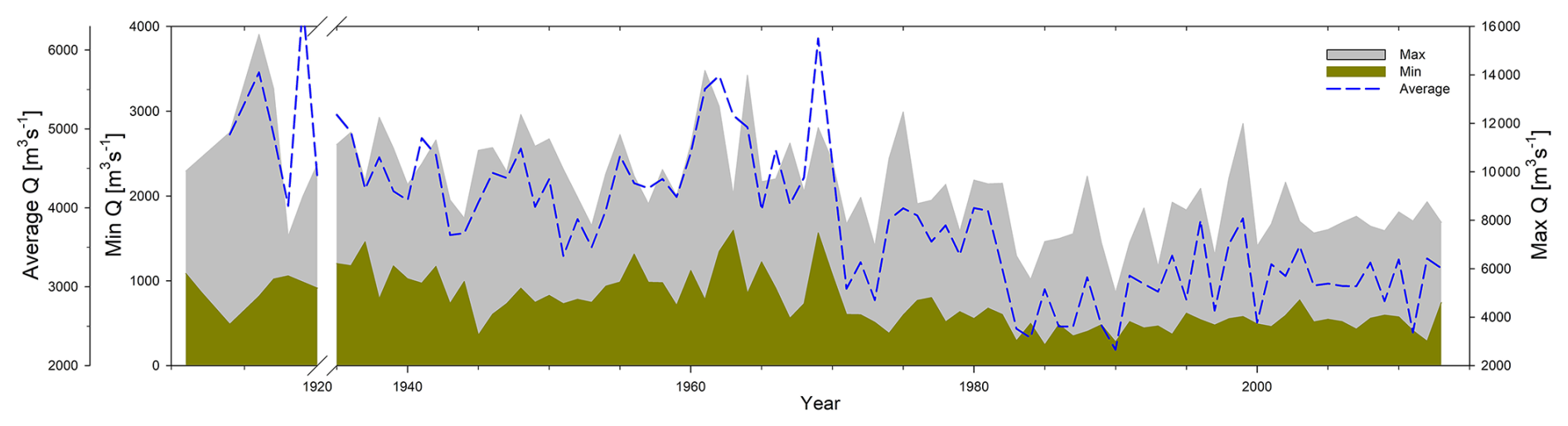
Figure 5Yearly maxima, minima, and average values of the centennial discharge record. Yearly maxima and minima were calculated as the average of the 10 consecutive highest and lowest measured values each calendar year.
4.2 Discharge reconstruction based on shell δ18O values
As determined by Kelemen et al. (2017), the Oubangui River at Bangui exhibits a logarithmic relationship between δ18Ow and Q (; R2=0.75). Due to the logarithmic nature of this relationship, the most robust Q reconstructions based on δ18Ow are obtained for low Q values (when the correlation is nearly linear), while for higher Q the data are more scattered, making the reconstruction less accurate. Using δ18Oshell values from archived bivalves and measured river Q, we investigated if the δ18Ow–Q relationship changed over time, thereby indicating changes in hydroclimate. The availability of bivalve shells collected at Bangui that cover suitable time frames was limited; only the very beginning of the continuous discharge data series is covered by shells collected at Bangui, as bivalves from Bangui were mainly collected before 1911. The other shells covering most of the record were collected downstream of Bangui. Although not all shells were collected at Bangui, they show similar δ18Oshell profiles, suggesting limited influence of downstream inputs on river water isotope values (Fig. 2).
The shells record a large shift in dry-season river water δ18O values and thereby suggest a major change in watershed hydrology and/or hydroclimate. Dry conditions and low flow are reflected in shells as higher δ18O values. Thus, if we assume as a first approach that the δ18Ow–Q relationship measured between 2009–2013 (Kelemen et al., 2017) can be considered representative of the full time frame of our shell collection, substantially lower peak δ18Oshell values in historical specimens suggest that Q was substantially higher during low-flow conditions at Bangui in the past. The δ18Oshell record also suggests that the δ18Ow values characterising the high Q remained similar over the past 100 years (based on δ18Oshell minima being similar through time (Fig. 3)). Constant δ18Ow values over the past century would suggest that the hydroclimatology of the high-flow period has not changed considerably.
A large decrease was observed in the Q record during the 1970s (Fig. 5); however, high and low flows were not evenly affected: peak flow decreased by 35 %, while the flow minima decreased by 60 % (Orange et al., 1997). Flow minima are indeed thought to be more sensitive to hydroclimatological changes than high (or annual) flows (Wesselink et al., 1996; Bricquet et al., 1997; Orange et al., 1997). This might be explained by the different origin of water during high- and low-flow periods: while peak flow is fed by surface runoff directly provided by precipitation, baseflow (low flow during the dry season) is maintained by riparian aquifers and water stored in upstream channels (Delleur, 1999; Brutsaert, 2005). Therefore, we hypothesise that this different response of discharge phases during the 1970s was when changes in the sources of water feeding the river (precipitation, groundwater) likely also occurred.
The lower amplitude and generally lower δ18Oshell values in historical shell specimens are qualitatively consistent with the trends in long-term Q data, and this trend can be extended into the late 19th century when Q data are not available. However, it must be kept in mind that the Q–δ18Ow relationship might vary over time and that the one established based on recent data might not be applicable to the historical data. To explore this possibility, we compared the reconstructed Q data (using the recent Q–δ18Ow relationship) with the empirical Q record, which indeed indicates that discharge during low flow is overestimated in the older shells (Fig. 6). In addition, the reconstructed high-discharge record exceeds the Q above which C. wissmanni seems to temporarily stop precipitating carbonate (Kelemen et al., 2017). It should be kept in mind, however, that the shell from 1948 was collected downstream of Bangui, and this may affect the Q–δ18Ow relationship locally (as discussed further on). Nevertheless, the 1914 shell, collected at Bangui, appears to overestimate dry-season Q, and thus our data indicate that the Q–δ18Ow relationship might indeed have changed through time.
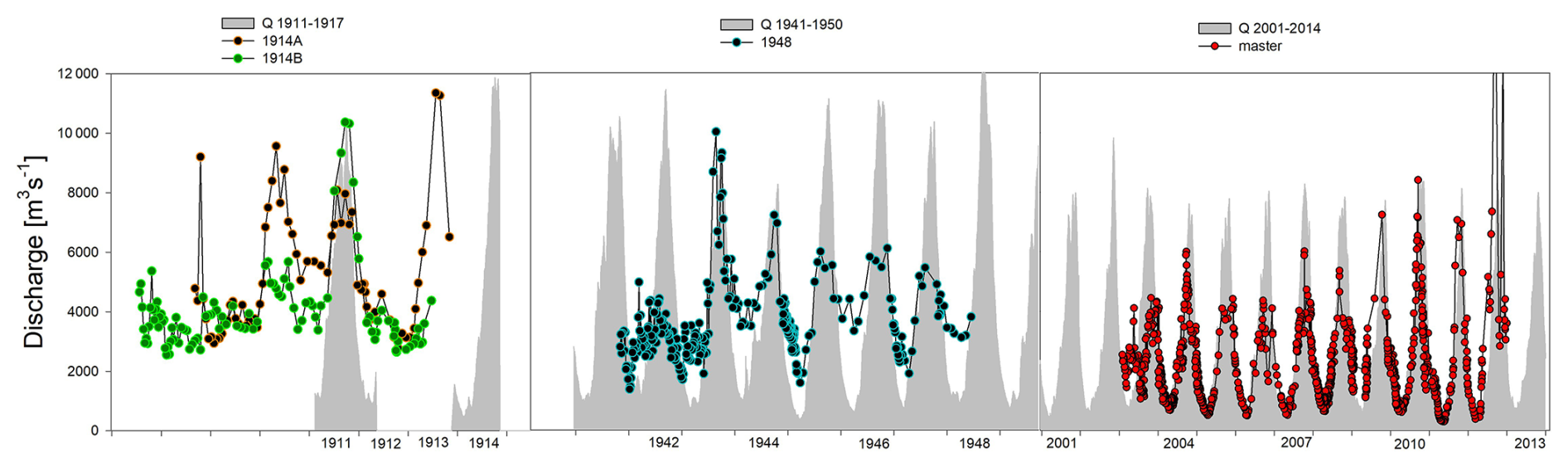
Figure 6Reconstructed discharge of the Oubangui River based on shell δ18O values and the logarithmic δ18Ow–Q relationship measured for the period 2009–2013 (coloured circles). The graph includes shell data that coincide with the available discharge measurements (grey areas), where δ18Oshell values were aligned with the best-matching discharge data after aligning the collection date.
4.3 Influence of vegetation cover, precipitation, and groundwater on δ18Ow values
In order to explore possible mechanisms leading to a proposed difference in the δ18Ow–Q relationship between recent direct observations and that based on pre-1960 bivalve shell data, several factors should be considered. We first examine whether a change in precipitation could influence the relationship and whether a shift in dry-season δ18Ow values is more likely to be caused by large-scale or by more regional-scale changes in precipitation and evapotranspiration. Other possibilities are also explored, such as the possible influence of the shell collection site (distance from Bangui along the Oubangui River) or possible impacts of changing vegetation cover and of groundwater fluctuations.
Influence of precipitation
Specimens collected before 1960 showed a generally smaller δ18Oshell range and recorded only the lower range of δ18Oshell values in comparison with the shells collected after 2010 (Fig. 3), which suggests a substantially higher dry-season river flow (with correspondingly lower δ18Ow values) during the first part of the 20th century. However, it is also possible that substantial changes in the precipitation regime, and thus in δ18O values, have occurred in the region during this period.
In the centennial rainfall record from Bangui, a large decrease in yearly rainfall amount was observed following the wet decade in the 1960s (Orange et al., 1997; Nguimalet and Orange, 2013, 2019). This precipitation regime shift was noticed throughout central Africa and over a major part of west Africa as well. The year 1970 is most commonly accepted as being the beginning of a major drought in sub-Saharan Africa, with a precipitation decrease of up to 40 % (Mahé and Olivry, 1999; Mahé et al., 2001; Hulme et al., 2001; Dai et al., 2004). According to Nguimalet and Orange (2019), statistical calculations found only a single break in the rainfall record over the Oubangui basin for the period 1935–2015, a break that occurred in 1970, after which the rainfall decreased by about 5 %, which cannot directly explain the annual average Q decrease of 22 % for the 1982–2013 period at Bangui (Nguimalet and Orange, 2019). However, these studies focused on yearly averaged rainfall data – we refined this analysis for two stations with a continuous multi-decadal monthly rainfall record in the Oubangui basin upstream of Bangui: Bangui (1907–2007) and Bambari (1931–2007). When monthly data were separated by dry season (December, January, February) and wet season (remaining months), it became apparent that the amount of precipitation during wet months did not change notably at either of the stations (Fig. 7). Conversely, precipitation during dry months (in particular December and January) gradually decreased at Bambari starting after 1970. In comparison to the period of 1931–2007 (the period for which we have data coverage for both stations), between 1982 and 2007 the dry-season precipitation decreased by 34 % at Bambari and by 18 % at Bangui, while the wet-season precipitation decreased by less than 2 % at Bambari and 3 % at Bangui. Moreover, rainfall in March at Bambari shows a decreasing trend since 1982, which suggests that the dry season may have increased in length in this area.
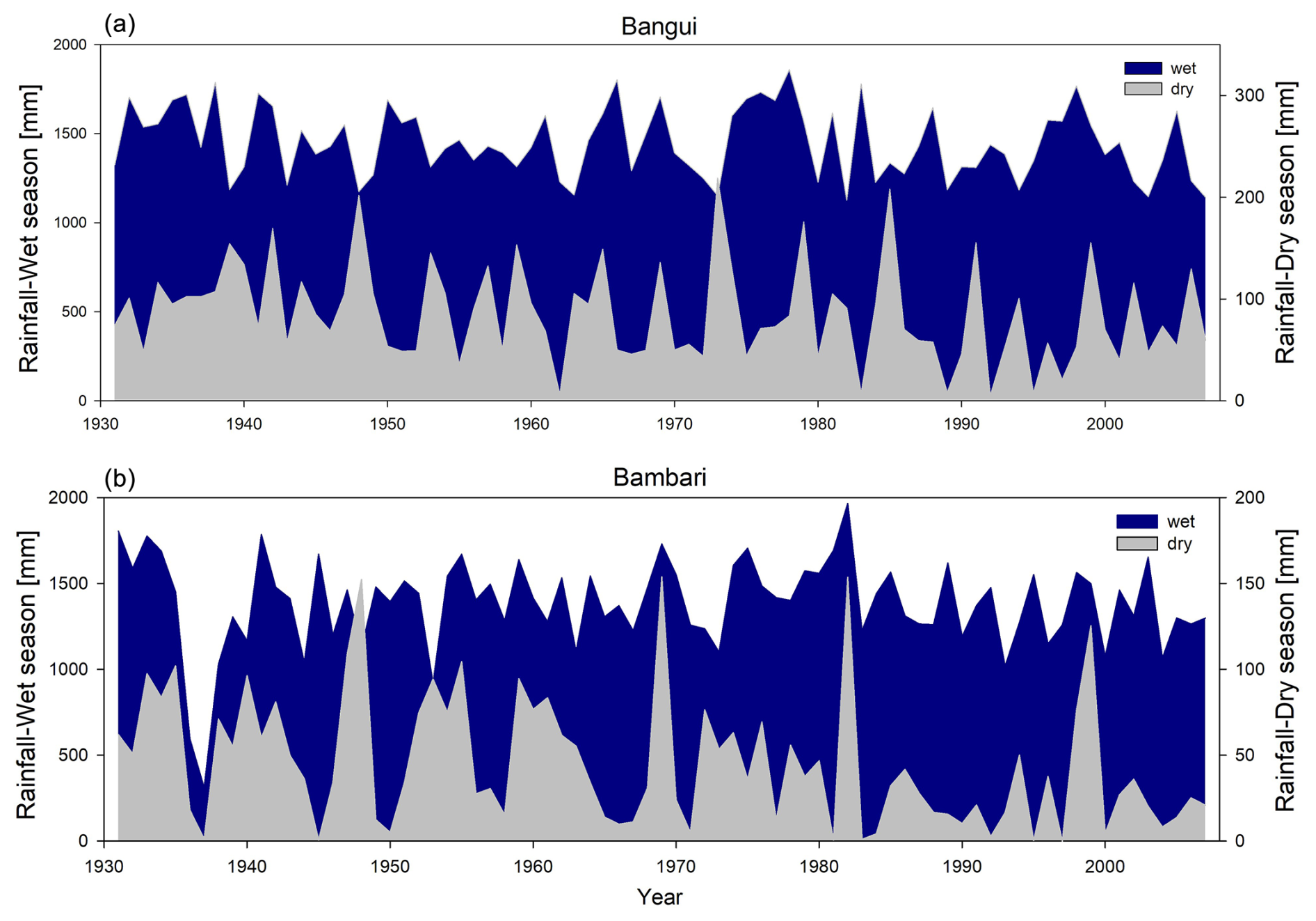
Figure 7Area plot of the dry-season (light area) and wet-season (dark area) rainfall amount at Bangui (a) and Bambari (b) station. December, January, and February are included in the dry season, while March to November represents the wet season.
Changes in precipitation amount, air mass trajectory, and temperature lead to changes in δ18Oprecip values (Gat, 1996). If during the dry season less rainfall occurs, the precipitation–evaporation balance shifts towards evaporation and river water δ18O values are expected to increase. Indeed, rainfall is usually more depleted in 18O in comparison with the water source because the water molecules with lighter isotopes evaporate faster; thus the residual evaporating source becomes more enriched. At times of heavy rainfall, rain is usually very depleted in 18O as a result of more complete rainout of water vapour, a process referred to as the “amount effect” (Dansgaard, 1964). In contrast, dry-season precipitation is typically 18O-enriched in comparison to that during the wet season (Dansgaard, 1964; Gat, 1996; Balagazi and Liotta, 2019). The precipitation δ18O record at Bangui indeed shows the amount effect: during abundant rainfall the δ18O values are lower than δ18O values of low rain amounts (Fig. 8). However, wet- and dry-season precipitation data are not perfectly separated in terms of isotope composition. All these factors could lead to an increase in δ18Ow values during the dry season, which was clearly recorded in the recently collected shells.
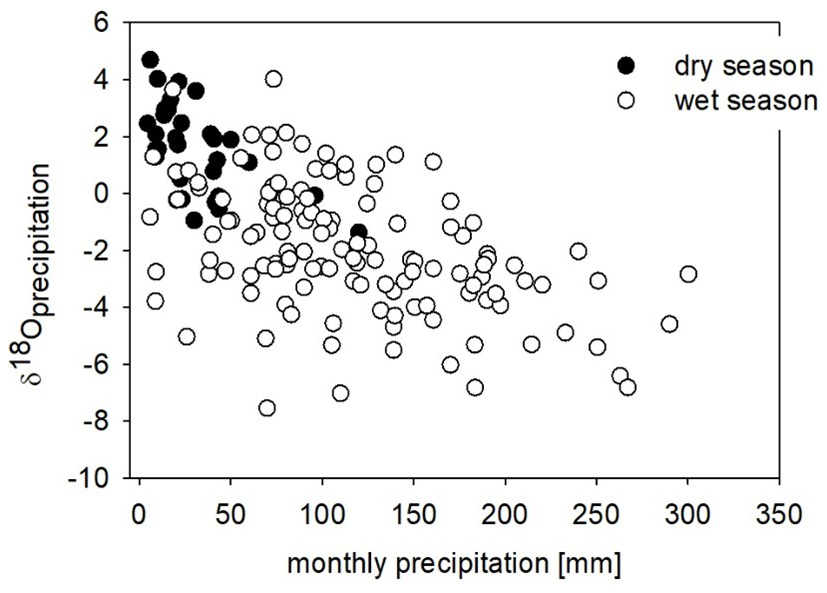
Figure 8Precipitation amount and precipitation δ18O values at Bangui for the period 2009–2018 (two collection sites, data from the IAEA-GNIP database; two outliers were removed). Filled circles represent precipitation during the dry season (December, January, February) and open circles that during the wet season (from March–November). The weighted average δ18O value is −2.3 ‰.
Influence of the collection site and vegetation cover
The Oubangui basin has a diverse landscape; the central region where the river receives only right-bank tributaries is covered by tree savannahs, while the lower section (downstream of Bangui) flows through rainforests. Not all museum-archived shells were collected near Bangui, and the river location is important to consider in interpreting environmental signals, as it is possible that the specimens collected downstream of the confluence with the Lobaye (shells 1904, 1948, 1950s, 1908, and 1891A; see Figs. 1 and 2) may be influenced by inputs from tributaries that drain rainforests. The Lobaye River represents the boundary between savannah and rainforest in the basin (see Bouillon et al., 2014; Kelemen, 2019). The Lobaye and its right-bank tributaries flow through the humid Ngotto rainforest, which is considerably different from most of the upper Oubangui watershed. The Ngotto rainforest likely has dry-season precipitation with lower δ18Ow values compared to the Oubangui watershed. The Lobaye River has a relatively stable discharge throughout the year, with a range on average between 240 and 500 m3 s−1 and a of about 2 for the available record (1950–1987) at the Mbata gauge station. Given the low variation in Q, δ18Ow values are expected to be similarly uniform; thus the very few available measured river δ18Ow values, which ranged between −3.0 ‰ and −3.3 ‰ (n=13, November 2012; Kelemen, 2019), are most likely representative (similar to δ18O measured in a single rain event on 20 November 2012: −1.9 ‰). During the low-Q period, the Lobaye River provides nearly 50 % of the flow of the lower Oubangui River, which isotopically should reflect Ngotto rainforest precipitation. While in the two bivalves collected just below the confluence with Lobaye (shell 1904 and 1948), δ18O values occasionally reached values somewhat higher than in historical bivalves collected more upstream, most of their δ18O record fell in the same range.
The two shells collected near the confluence with the Congo River (shell 1908 and 1891A) showed a very narrow range of δ18Oshell values, covering only the lowest δ18Oshell values recorded in the other shells. This corroborates the hypothesis that the three rainforest rivers (Ibenga, Motaba, and Ngiri) contribute to steadier and lower δ18Ow values in the lower Oubangui (Fig. 2). Nevertheless, the older shells collected at, or upstream of, Bangui (shells 1914A, 1914B, and 1891D) all have similar isotope profiles to shells collected downstream of the Lobaye. This is particularly evident by direct comparison of shells 1891D and 1891A, which were both collected in the same year but grew in different sections of the river: 1891D close to Bangui and 1891A ∼500 km downstream (Fig. 2). This suggests that the archived shells do indeed reflect dry-season precipitation with lower δ18O values in the Oubangui and that the long-term trend in the δ18Oshell record is not skewed due to some shells having been collected downstream of rainforest watersheds.
Influence of groundwater
Orange et al. (1997) noted the sponge-like functioning of the majority of the Oubangui basin, based on the 3-year time lag between the decrease in rainfall and the change in the Q record. This behaviour of the basin might be the main reason why the Q decline at Bangui was so intense and rapid after 1970, while the rainfall record showed only minor changes with a sudden reduction in precipitation in 1968 (Nguimalet and Orange, 2013, 2019). Moreover, a study of the Oubangui at Mobaye (upstream of Bangui) showed that at the northern tributary (Kotto), which exclusively drains savannah, it had its infiltration capacity to recharge the aquifer altered by the long drought (Nguimalet et al., 2022). This would result in the abundant precipitation being diverted from recharge to runoff, especially after the drought of the 1970s (Wesselink et al., 1996; Bricquet et al., 1997). The drier period starting around 1970 would thus have led to a decrease in groundwater storage, which is currently not high enough to sustain the hydrological regime as it did before during the dry season (Orange et al., 1997). In contrast, the southwest of the basin (downstream of Bangui) lies above a semi-continuous aquifer which contributes to river Q during low flow (Laraque et al., 2001), and this might explain why the Oubangui at Bangui remained in a drought phase while the Congo River at Brazzaville returned to its normal flow conditions after the 1990s (Laraque et al., 2013).
Jasechko and Taylor (2015) found that groundwater stable isotope values across different tropical catchments typically followed those of annual precipitation, weighted towards more rainfall-intense months. As the aquifer is recharged by rainwater (in particular during wet periods), it preserves the δ18O values of the precipitation. Thus during low flow, when the groundwater is the main supply of the river, δ18Ow values are expected to reflect the δ18O values of the annual precipitation. If that principle is applicable to our study basin, this would suggest that groundwater δ18O values would be lower than −2.3 ‰ (i.e. the amount-weighted average precipitation value at Bangui; see Fig. 8). Indeed, groundwater δ18O values in shallow (3–20 m deep) wells reported for Bangui range between −3.3 ‰ and −2 ‰ (Djebebe-Ndjiguim et al., 2019), while in the region of Mambéré-Kadéi (the region drained by the Lobaye River) west of Bangui, measured borehole water δ18O values range between −3.5 ‰ and −2.4 ‰ (Foto et al., 2019).
These groundwater δ18Ow values are in good agreement with the highest δ18Oshell values of the historical specimens, as all showed low peak δ18O values during times of low Q in comparison with the δ18Oshell values in recent specimens. This favours the hypothesis that a (partial) change in the source and composition of the low-flow water occurred. According to this hypothesis, the Oubangui has received a decreased groundwater contribution (low δ18O values) during times of low Q since the drought in the 1970s, with the majority of the water sourced from the upper reaches of the river, where the water travels hundreds of kilometres through savannah, and from 18O-enriched precipitation. According to Nguimalet and Orange (2019), the volume mobilised by the Oubangui aquifer has been gradually decreasing since 1970; in some years the water provided by the aquifer has been less than 50 % of the amount before the drought. Dry-season precipitation with higher δ18O values and without the dilution of groundwater would then be consistent with shells recording increased δ18O values.
4.4 Shell δ13C values
Some reports postulate that shell δ13C values track the δ13C of the dissolved inorganic carbon (DIC) in the host water (Fritz and Poplawski, 1974; Poulain et al., 2010; Graniero et al., 2017). However, several authors have found disequilibrium between DIC and freshwater shell δ13C values (Dettman et al., 1999; Gillikin et al., 2009; Kelemen et al., 2017; Graniero et al., 2021), while some studies have reported constant fractionation (Abell and Hoelzmann, 2000; Kaandorp et al., 2003). There is a strong inverse relationship between Q and δ13CDIC values in the Oubangui today (Bouillon et al., 2012, 2014). Bivalves collected in the 1950s and earlier exhibited pronounced cyclicity, with an amplitude between 5‰ and 7.9‰ (Fig. 4), which is higher than the amplitude (between 2.9‰ and 6.4‰) found in recently collected bivalves from Oubangui River (Kelemen et al., 2017). At the time of shell precipitation, δ13Cshell values are derived from dissolved inorganic carbon (DIC) and the bivalve's own metabolic (respired) carbon (McConnaughey et al., 1997; McConnaughey and Gillikin, 2008); however the proportion is highly variable among species (McConnaughey et al., 1997; Lorrain et al., 2004; Gillikin et al., 2006, 2007, 2009; Poulain et al., 2010; Graniero et al., 2021). While young specimens during fast growth mainly use carbon from the DIC pool, older bivalves incorporate a relatively high amount of metabolically derived carbon which is more 13C-depleted than the DIC pool (Lorrain et al., 2004). In all analysed specimens, an ontogenetic decrease in δ13C values was observed, which is in agreement with other studies suggesting an inclination towards metabolic carbon incorporation in older age (Lorrain et al., 2004; Gillikin et al., 2006, 2007, 2009; McConnaughey and Gillikin, 2008). In the Oubangui basin, the weathering regime (silicate versus carbonate weathering) exerts strong control on δ13CDIC values, but there is also strong seasonality with higher values during low-flow conditions, likely due to a combination of increased aquatic primary production and CO2 outgassing during low flow (Bouillon et al., 2012). A reduction in precipitation could have an indirect effect on δ13Cshell values by increasing δ13CDIC values of the water during low flow, but this is not evident in the shell data. δ13Cshell values did not closely match δ13CDIC in a previous study on C. wissmanni shells, and thus they are not quantitative environmental proxies (Kelemen et al., 2017); however, they might qualitatively indicate large-scale changes in watershed characteristics (e.g. Goewert et al., 2007).
The analysis of δ18O values in historical and contemporary freshwater bivalve shells from the Oubangui River demonstrated a clear baseline shift in river δ18O values over time. Shells collected between the late 19th and mid-20th century exhibited a much narrower range of δ18O values than recently collected shells. The seasonal cyclicity was nevertheless pronounced, and our data indicate a higher intra-annual variation in river water δ18O values in recent times, indicating a major shift in the hydroclimate baseline. The long-term Q record shows a decline in both high- and low-Q conditions, and when Q is reconstructed based on δ18O data from historical shells and a recently established δ18Ow–Q relationship, a divergence in low-flow periods suggests that a change in the δ18Ow–Q relationship might have occurred. The mechanisms underlying such a shift are, however, not straightforward to identify. According to Nguimalet and Orange (2013, 2019), there have only been minor changes in precipitation patterns since 1935, and the δ18Oshell values during times of high Q (wet season) appeared to be similar in historical and recent shells, which suggests no substantial shift in (wet-season) precipitation δ18O values over the past century. However, the monthly precipitation data from Bangui and Bambari revealed a decrease in dry-season rainfall amounts, which should directly or indirectly influence the river water δ18O. The aquifer recharging capacity above Bangui might have become more limited since the severe drought in the 1970s. As the aquifer is mainly expected to recharge during abundant rainfall (with low δ18O values), the increased δ18O values during low-flow conditions in recent times coupled with significantly decreased low Q would suggest that aquifers upstream of the rainforest belt have not yet completely recovered their capacity to support the river baseflow.
Further studies of contemporary and past δ18Ow patterns in precipitation across the basin (e.g. from direct precipitation sampling and tree core δ18O studies) should be able to provide more insight into changes in the hydroclimate of the Oubangui basin and other such river basins for which no historical Q data are available. While we have previously shown that tropical bivalves living in rivers with near-constant annual water temperatures faithfully record water δ18O values in their shells, this study shows that interpreting the δ18Ow values using the δ18Ow–Q relationship is more complex as several parameters can influence this relationship. Nevertheless, large changes in river discharge over the past century were clearly reflected in shell δ18O values. We posit that archived collections of freshwater shells can provide a wealth of hydroclimate proxy data covering the past century.
The full set of stable isotope data on historical shells is provided in the Supplement.
The supplement related to this article is available online at https://doi.org/10.5194/bg-22-2621-2025-supplement.
This study was conceived by DPG and SB. All authors contributed to sample collection (recent and/or historical), sample analyses, and data interpretation. ZK drafted the initial version of the manuscript, which was revised by DPG and SB.
At least one of the (co-)authors is a member of the editorial board of Biogeosciences. The peer-review process was guided by an independent editor, and the authors also have no other competing interests to declare.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
We thank Didier van der Spiegel at the Royal Museum for Central Africa (MRAC), Yves Samyn at the Royal Belgian Institute of Natural Sciences (KBIN), and Philippe Bouchet and Virginie Héros at the MNHN Paris for providing access to museum collections. We thank Dan Graf for help in identifying the shell specimens and for his work on the MusselP database (http://mussel-project.uwsp.edu, last access: 15 February 2020), Mason Stahl (Union College) and Thibault Lambert for producing Fig. 1, and Anouk Verheyden (Union College) for lab assistance. We also thank Athanase Yambélé (Direction de la Météorologie Nationale, Bangui), who was instrumental in the collection of contemporary shells and data that made this follow-up study possible, Gil Mahé (IRD, Montpellier, France) and Julien Thébault (Université de Bretagne Occidentale, Brest, France) are acknowledged for their constructive discussions on the data, and the two anonymous reviewers are acknowledged for their insightful and constructive feedback during the review process.
Funding for this study was provided by the European Research Council (ERC Starting Grant StG 240002, AFRIVAL) to Steven Bouillon, a National Geographic Society Research and Exploration Grant (grant no. 8885-11) to David P. Gillikin and Steven Bouillon, the KU Leuven Special Research Fund (DBOF PhD scholarship to Zita Kelemen), the Research Foundation Flanders (FWO-Vlaanderen; research project G.0D87.14N and travel grants) to Steven Bouillon and David P. Gillikin, and a Research Corporation for Science Advancement Single-Investigator Cottrell College Science Award (no. 20169) to David P. Gillikin. The US National Science Foundation funded Union College's isotope ratio mass spectrometer and peripherals (NSF-MRI no. 1229258).
This paper was edited by Sebastian Naeher and reviewed by two anonymous referees.
Abell, P. I. and Hoelzmann, P.: Holocene palaeoclimates in northwestern Sudan: stable isotope studies on molluscs, Global Planet. Change, 26, 1–12, https://doi.org/10.1016/S0921-8181(00)00030-8, 2000.
Abell, P. I., Amegashitsi, L., and Ochumba, P. B.: The shells of Etheria elliptica as recorders of seasonality at Lake Victoria, Palaeogeogr. Palaeocl., 119, 215–219, https://doi.org/10.1016/0031-0182(95)00019-4, 1996.
Alsdorf, D., Beighley, E., Laraque, A., Lee, H., Tshimanga, R., O'Loughlin, F., Mahé, G., Dinga, B., Moukandi, G., and Spencer, R. G.: Opportunities for hydrologic research in the Congo Basin, Rev. Geophys., 54, 378–409, https://doi.org/10.1002/2016RG000517, 2016.
Amogu, O., Esteves, M., Vandervaere, J.-P., Malam Abdou, M., Panthou, G., Rajot, J.-L., Souley Yéro, K., Boubkraoui, S., Lapetite, J.-M., Dessay, N., Zin, I., Bachi, R. A., Bouzou Moussa, I., Faran Maïga, O., Gautier, E., Mamadou, I., and Descroix, L.: Runoff evolution due to land-use change in a small Sahelian catchment, Hydrolog. Sci. J., 60, 78–95, https://doi.org/10.1080/02626667.2014.885654, 2014.
Asefi-Najafabady, S. and Saatchi, S.: Response of African humid tropical forests to recent rainfall anomalies, Philos. T. R. Soc. Lon. B., 368, 20120306, https://doi.org/10.1098/rstb.2012.0306, 2013.
Balagazi, C. M. and Liotta, M.: Key factors of precipitation stable isotope fractionation in Central-Eastern Africa and Central Mediterranean, Geosciences, 9, 337, https://doi.org/10.3390/geosciences9080337, 2019.
Barker, P. A., Hurrell, E. R., Leng, M. J., Wolff, C., Cocquyt, C., Sloane, H. J., and Verschuren, D.: Seasonality in equatorial climate over the past 25 k.y. revealed by oxygen isotope records from Kilimanjaro, Geology, 39, 1111–1114, https://doi.org/10.1130/G32419.1, 2011.
Battipaglia, G., Zalloni, E., Castaldi, S., Marzaioli, F., Cazzolla-Gatti, R., Lasserre, B., Tognetti, R., Marchetti, M., and Valentini, R.: Long tree-ring chronologies provide evidence of recent tree growth decrease in a central african tropical forest, PLoS One, 10, 1–21, https://doi.org/10.1371/journal.pone.0120962, 2015.
Berke, M. A., Johnson, T. C., Werne, J. P., Schouten, S., and Sinninghe Damsté, J. S.: A mid-Holocene thermal maximum at the end of the African Humid Period, Earth Planet. Sc. Lett., 351–352, 95–104, https://doi.org/10.1016/j.epsl.2012.07.008, 2012.
Bouillon, S., Yambélé, A., Spencer, R. G. M., Gillikin, D. P., Hernes, P. J., Six, J., Merckx, R., and Borges, A. V.: Organic matter sources, fluxes and greenhouse gas exchange in the Oubangui River (Congo River basin), Biogeosciences, 9, 2045–2062, https://doi.org/10.5194/bg-9-2045-2012, 2012.
Bouillon, S., Yambélé, A., Gillikin, D. P., Teodoru, C., Darchambeau, F., Lambert, T., and Borges, A. V.: Contrasting biogeochemical characteristics of the Oubangui River and tributaries (Congo River basin), Sci. Rep.-UK, 4, 5402, https://doi.org/10.1038/srep05402, 2014.
Boulvert, Y. : République Centrafricaine, carte Orohydrographiqueu 11 000 000, O. R. S. T. O.M, Paris, 1987.
Bricquet, J. P., F. Bamba, G. Mahé, M. Touré, and Olivry, J. C.: Water resource variations of the Atlantic river basins of Africa: The long term effect of rain shortage, J. Water Sci. (Rev. Sci. Eau), 10, 321–337, https://doi.org/10.7202/705282ar, 1997.
Brutsaert, W.: Hydrology: an introduction, Cambridge University Press, https://doi.org/10.1017/CBO9780511808470, 2005.
Casella, L. A., Griesshaber, E., Yin, X., Ziegler, A., Mavromatis, V., Müller, D., Ritter, A.-C., Hippler, D., Harper, E. M., Dietzel, M., Immenhauser, A., Schöne, B. R., Angiolini, L., and Schmahl, W. W.: Experimental diagenesis: insights into aragonite to calcite transformation of Arctica islandica shells by hydrothermal treatment, Biogeosciences, 14, 1461–1492, https://doi.org/10.5194/bg-14-1461-2017, 2017.
Dai, A., Trenberth, K. E., and Qian, T.: A global dataset of Palmer Drought Severity Index for 1870–2002: Relationship with soil moisture and effects of surface warming, J. Hydrometeorol., 5, 1117–1130, https://doi.org/10.1175/JHM-386.1, 2004.
Dansgaard, W.: Stable isotopes in precipitation, Tellus, 16, 438–468, https://doi.org/10.1111/j.2153-3490.1964.tb00181.x, 1964.
Delleur, J. W.: The handbook of groundwater engineering, CRC Press, Boca Raton, FL, https://doi.org/10.1201/9781420006001, 1999.
Descroix, L., Genthon, P., Amogu, O., Rajot, J. L., Sighomnou, D., and Vauclin, M.: Change in Sahelian Rivers hydrograph: the case of recent red floods of the Niger River in the Niamey region, Global Planet. Change, 98, 18–30, https://doi.org/10.1016/j.gloplacha.2012.07.009, 2012.
Dettman, D. L. and Lohmann, K.: Oxygen isotope evidence for high-altitude snow in the Laramide Rocky Mountains of North America during the Late Cretaceous and Paleogene, Geology, 28, 243–246, https://doi.org/10.1130/0091-7613(2000)28<243:OIEFHS>2.0.CO;2, 2000.
Dettman, D. L., Flessa, K. W., Roopnarine, P. D., Schöne, B. R., and Goodwin, D. H.: The use of oxygen isotope variation in shells of estuarine mollusks as a quantitative record of seasonal and annual Colorado River discharge, Geochim. Cosmochim. Ac., 68, 1253–1263, https://doi.org/10.1016/j.gca.2003.09.008, 2004.
Dettman, D. L., Kohn, M. J., Quade, J., Ryerson, F. J., Ojha, T. P., and Hamidullah, S.: Seasonal stable isotope evidence for a strong Asian monsoon throughout the past 10.7 my, Geology, 29, 31–34, https://doi.org/10.1130/0091-7613(2001)029<0031:SSIEFA>2.0.CO;2, 2001.
Dettman, D. L., Reische, A. K., and Lohmann, K. C.: Controls on the stable isotope composition of seasonal growth bands in aragonitic fresh-water bivalves (Unionidae), Geochim. Cosmochim. Ac., 63, 1049–1057, https://doi.org/10.1016/S0016-7037(99)00020-4, 1999.
Djebebe-Ndjiguim, C. L., Mbane, J. S., Foto, E., Kongandembou, M. L., and Mabingui, J.: Impacts of cracks on underground flows of the superficial aquifer: case of the northern sector of the city of Bangui, Int. J. Adv. Res. Publ., 3, 80–89, 2019.
Essaid, H. I. and Caldwell, R. R.: Evaluating the impact of irrigation on surface water – groundwater interaction and stream temperature in an agricultural watershed, Sci. Total Environ., 599–600, 581–596, https://doi.org/10.1016/j.scitotenv.2017.04.205, 2017.
Foto, E., Djebebe Ndjinguim, C. L., Zoudamba, N., Basse Keke, E., and Mabingui, J.: Etude hydrogéochimique des eaux souterraines dans la formation Gréseuse de la région de Berberati en République Centrafricaine, Eur. Sci. J., 15, 487–507, https://doi.org/10.19044/esj.2019.v15n6p487, 2019.
Fritz, P. and Poplawski, S.: 18O and 13C in the shells of freshwater molluscs and their environments, Earth Planet. Sc. Lett., 24, 91–98, https://doi.org/10.1016/0012-821X(74)90012-0, 1974.
Gat, J. R.: Oxygen and hydrogen isotopes in the hydrologic cycle, Annu. Rev. Earth Pl. Sc. 24, 225–262, https://doi.org/10.1146/annurev.earth.24.1.225, 1996.
Gillikin, D. P., Lorrain, A., Bouillon, S., Willenz, P., and Dehairs, F.: Stable carbon isotopic composition of Mytilus edulis shells: relation to metabolism, salinity δ13CDIC and phytoplankton, Org. Geochem., 37, 1371–1382, https://doi.org/10.1016/j.orggeochem.2006.03.008, 2006.
Gillikin, D. P., Lorrain, A., Meng, L., and Dehairs, F.: A large metabolic carbon contribution to the δ13C record in marine aragonitic bivalve shells, Geochim. Cosmochim. Ac., 71, 2936–2946, https://doi.org/10.1016/j.gca.2007.04.003, 2007.
Gillikin, D. P., Hutchinson, K. A., and Kumai, Y.: Ontogenic increase of metabolic carbon in freshwater mussel shells (Pyganodon cataracta), J. Geophys. Res.-Biogeo., 114, 1–6, https://doi.org/10.1029/2008JG000829, 2009.
Goewert, A., Surge, D., Carpenter, S. J., and Downing, J.: Oxygen and carbon isotope ratios of Lampsilis cardium (Unionidae) from two streams in agricultural watersheds of Iowa, USA, Palaeogeogr. Palaeocl., 252, 637–648, https://doi.org/10.1016/j.palaeo.2007.06.002, 2007.
Gonfiantini, R., Stichler, W., and Rozanski, K.: Standards and intercomparison materials distributed by the International Atomic Energy Agency for stable isotope measurements, IAEA-Techdoc-825, ISSN 1011-4289, 13–29, 1995.
Goodwin, D. H., Gillikin, D. P., Banker, R., Watters, G. T., Dettman, D. L., and Romanek, C. S.: Reconstructing intra-annual growth of freshwater mussels using oxygen isotopes, Chem. Geol., 526, 7–22, https://doi.org/10.1016/j.chemgeo.2018.07.030, 2019.
Graf, D. L. and Cummings, K. S.: Freshwater mussel (Mollusca: Bivalvia: Unionoida) richness and endemism in the ecoregions of Africa and Madagascar based on comprehensive museum sampling, Hydrobiologia, 678, 17–36, https://doi.org/10.1007/s10750-011-0810-5, 2011.
Graniero, L. E., Grossman, E. L., Robbins, J., Morales, J., Thompson, R., and O'Dea, A.: Conus shell δ13C values as proxies for δ13CDIC in tropical waters, Palaeogeogr. Palaeocl., 472, 119–127, https://doi.org/10.1016/j.palaeo.2017.02.007, 2017.
Graniero, L. E., Gillikin, D. P., and Surge, D.: Evaluating freshwater mussel shell δ13C values as a proxy for dissolved inorganic carbon δ13C values in a temperate river, J. Geophys. Res.-Biogeo., 126, e2020JG006003, https://doi.org/10.1029/2020JG006003, 2021.
Grossman, E. L. and Ku, T.-L.: Oxygen and carbon isotope fractionation in biogenic aragonite: temperature effects, Chem. Geol. Isot. Geosci. Sect., 59, 59–74, https://doi.org/10.1016/0168-9622(86)90057-6, 1986.
Hulme, M., Doherty, R., Ngara, T., New, M., and Lister, D.: African climate change: 1900–2100, Clim. Res., 17, 145–168, https://doi.org/10.3354/cr017145, 2001.
Jasechko, S. and Taylor, R. G.: Intensive rainfall recharges tropical groundwaters, Environ. Res. Lett., 10, 124015, https://doi.org/10.1088/1748-9326/10/12/124015, 2015.
Kaandorp, R. J. G., Vonhof, H. B., Del Busto, C., Wesselingh, F. P., Ganssen, G. M., Marmól, A. E., Romero Pittman, L., and Van Hinte, J. E.: Seasonal stable isotope variations of the modern Amazonian freshwater bivalve Anodontites trapesialis, Palaeogeogr. Palaeocl., 194, 339–354, https://doi.org/10.1016/S0031-0182(03)00332-8, 2003.
Kaandorp, R. J. G., Vonhof, H. B., Wesselingh, F. P., Pittman, L. R., Kroon, D., and Van Hinte, J. E.: Seasonal Amazonian rainfall variation in the Miocene climate optimum, Palaeogeogr. Palaeocl., 221, 1–6, https://doi.org/10.1016/j.palaeo.2004.12.024, 2005.
Kelemen, Z.: Reconstructing the biogeochemistry of tropical aquatic ecosystems using elemental and stable isotope tracers in freshwater bivalve shells, Ph. D. dissertation, KU Leuven, Belgium, 157 pp., 2019.
Kelemen, Z., Gillikin, D. P., Graniero, L. E., Havel, H., Darchambeau, F., Borges, A. V., Yambélé, A., Bassirou, A., and Bouillon, S.: Calibration of hydroclimate proxies in freshwater bivalve shells from Central and West Africa, Geochim. Cosmochim. Ac., 208, 41–62, https://doi.org/10.1016/j.gca.2017.03.025, 2017.
Kelemen, Z., Gillikin, D. P., Borges, A. V., Tambwe, E., Sembaito, A. T., Mambo, T., Wabakhangazi, J. N., Yambélé, A., Stroobandt, Y., and Bouillon, S.: Freshwater bivalve shells as hydrologic archives in the Congo Basin, Geochim. Cosmochim. Ac., 308, 101–117, https://doi.org/10.1016/j.gca.2021.05.023, 2021.
Laraque, A., Mahé, G., Orange, D., and Marieu, B.: Spatiotemporal variations in hydrological regimes within Central Africa during the XXth century, J. Hydrol., 245, 104–117, https://doi.org/10.1016/S0022-1694(01)00340-7, 2001.
Laraque, A., Bellanger, M., Adele, G., Guebanda, S., Gulemvuga, G., Pandi, A., Paturel, J. E., Robert, A., Tathy, J. P., and Yambélé, A.: Evolutions récentes des débits du Congo, de l'Oubangui et de la Sangha, Geo. Eco. Trop., 37, 93–100, 2013.
Laraque, A., Moukandi N'kaya, G. D., Orange, D., Tshimanga, R., Tshitenge, J. M., Mahé, G. Nguimalet, C. R., Trigg, M. A., Yepez, S, and Gulemvuga, G.: Recent budget of hydroclimatology and hydrosedimentology of the Congo River in Central Africa, Water, 12, 2613, https://doi.org/10.3390/w12092613, 2020.
Lorrain, A., Paulet, Y. M., Chauvaud, L., Dunbar, R., Mucciarone, D., and Fontugne, M.: δ13C variation in scallop shells: Increasing metabolic carbon contribution with body size?, Geochim. Cosmochim. Ac., 68, 3509–3519, https://doi.org/10.1016/j.gca.2004.01.025, 2004.
Mahé, G.: Modulation annuelle et fluctuations interannuelles des précipitations sur le bassin versant du Congo, Coll. PEGI/INSU/ORSTOM, Paris, ISBN 2-7099-1245-7, 22–24, 13–26, Novembre 1993.
Mahé, G. and Olivry, J. C.: Assessment of freshwater yields to the ocean along the intertropical Atlantic coast of Africa (1951–1989), CR. Acad. Sci. II A, 328, 621–626, https://doi.org/10.1016/S1251-8050(99)80159-1, 1999.
Mahé, G., L'hote, Y., Olivry, J. C., and Wotling, G.: Trends and discontinuities in regional rainfall of West and Central Africa: 1951–1989, Hydrolog. Sci. J., 46, 211–226, https://doi.org/10.1080/02626660109492817, 2001.
Malhi, Y. and Wright, J.: Spatial patterns and recent trends in the climate of tropical rainforest regions, Philos. T. R. Soc. B, 359, 311–329, https://doi.org/10.1098/rstb.2003.1433, 2004.
Mayaux, P., Richards, T., and Janodet, E.: A vegetation map of Central Africa derived from satellite imagery, J. Biogeogr., 26, 353–366, https://doi.org/10.1046/j.1365-2699.1999.00270.x, 1999.
McConnaughey, T. A., Burdett, J., Whelan, J. F., and Paull, C. K.: Carbon isotopes in biological carbonates: respiration and photosynthesis, Geochim. Cosmochim. Ac., 61, 611–622, https://doi.org/10.1016/S0016-7037(96)00361-4, 1997.
McConnaughey, T. A. and Gillikin, D. P.: Carbon isotopes in mollusk shell carbonates, Geo-Mar. Lett., 28, 287–299, https://doi.org/10.1007/s00367-008-0116-4, 2008.
Mologni, C., Revel, M., Chaumillon, E., Malet, E., Coulombier, T., Sabatier, P., Brigode, P., Hervé, G., Develle, A.-L., Schenini, L., Messous, M., Davtian, G., Carré, A., Bosch, D., Volto, N., Ménard, C., Khalidi, L., and Arnaud, F.: 50-year seasonal variability in East African droughts and floods recorded in central Afar lake sediments (Ethiopia) and their connections with the El Niño–Southern Oscillation, Clim. Past, 20, 1837–1860, https://doi.org/10.5194/cp-20-1837-2024, 2024.
Nguimalet, C. R. and Orange, D.: Dynamique hydrologique récente de l'Oubangui à Bangui (Centrafrique): impacts anthropiques ou climatiques?, Geo. Eco. Trop., 37, 101–112, 2013.
Nguimalet, C. R. and Orange, D. : Caractérisation de la baisse hydrologique de l'Oubangui à Bangui, République Centrafricaine. Colloque international Hydrologie des grands bassins fluviaux de l'Afrique, 26–30 October, Hammamet, Tunisie, 8 p., 2015.
Nguimalet, C. R., Orange, D. : Caractérisation de la baisse hydrologique actuelle de la rivière Oubangui à Bangui, République Centrafricaine, La Houille Blanche, 1, 78–84, 2019.
Nguimalet, C. R., Orange, D., Waterendji, J. P., and Yambélé, A.: Hydroclimatic dynamics of upstream Ubangui river at Mobaye, Central African Republic: comparative study of the role of savannah and equatorial forest. in: Congo basin hydrology, climate and biogeochemistry: a foundation for the future, edited by: Tshimanga, R. M., Moukandi N'kaya, G. D., and Alsdorf, D., Geophysical Monograph, American Geophysical Union, Wiley & Sons, https://doi.org/10.1002/9781119657002.ch6, 2022.
Orange, D., Wesselink, A. J., Mahé, G., and Feizoure, C. T.: The effects of climate changes on river baseflow and aquifer storage in Central Africa, Sustainability of Water Resources under Increasing Uncertainty, Proceedings of the Rabat Symposium, IAHS Publ. no. 240, 19, ISBN 1-801502-05-8, 1997.
Pfister, L., Thielen, F., Deloule, E., Valle, N., Lentzen, E., Grave, C., Beisel, J.-N., and McDonnell, J. J.: Freshwater pearl mussels as a stream water stable isotope recorder, Ecohydroclimate, 11, e2007, https://doi.org/10.1002/eco.2007, 2018.
Pfister, L., Grave, C., Beisel, J. N., and McDonnell, J. J.: A global assessment of freshwater mollusk shell oxygen isotope signatures and their relation to precipitation and stream water, Sci. Rep.-UK, 9, 1–6, https://doi.org/10.1038/s41598-019-40369-0, 2019.
Poulain, C., Lorrain, A., Mas, R., Gillikin, D. P., Dehairs, F., Robert, R., Paulet, Y.-M.: Experimental shift of diet and DIC stable carbon isotopes: influence on shell δ13C values in the Manila clam Ruditapes philippinarum, Chem. Geol., 272, 75–82, https://doi.org/10.1016/j.chemgeo.2010.02.006, 2010.
Recha, J. W., Lehmann, J., Todd Walter, M., Pell, A., Verchot, L., and Johnson, M.: Stream discharge in tropical headwater catchments as a result of forest clearing and soil degradation, Earth Interact., 16, 1–18, https://doi.org/10.1175/2012EI000439.1, 2012.
Ricken, W., Steuber, T., Freitag, H., Hirschfeld, M., and Niedenzu, B.: Recent and historical discharge of a large European river system – Oxygen isotopic composition of river water and skeletal aragonite of Unionidae in the Rhine, Palaeogeogr. Palaeocl., 193, 73–86, https://doi.org/10.1016/S0031-0182(02)00713-7, 2003.
Runge, J. and Nguimalet, C. R.: Physiogeographic features of the Oubangui catchment and environmental trends reflected in discharge and floods at Bangui 1911–1999, Central African Republic, Geomorphology, 70, 311–324, https://doi.org/10.1016/j.geomorph.2005.02.010, 2005.
Schöne, B. R., Meret, A. E., Baier, S. M., Fiebig, J., Esper, J., McDonnell, J., and Pfister, L.: Freshwater pearl mussels from northern Sweden serve as long-term, high-resolution stream water isotope recorders, Hydrol. Earth Syst. Sci., 24, 673–696, https://doi.org/10.5194/hess-24-673-2020, 2020.
Strayer, D. L. and Malcom, H. M.: Shell decay rates of native and alien freshwater bivalves and implications for habitat engineering, Freshwater Biol., 52, 1611–1617, https://doi.org/10.1111/j.1365-2427.2007.01792.x, 2007.
Stringer, C. A. and Prendergast, A. L.: Freshwater mollusc sclerochronology: trends, challenges, and future directions, Earth Sci. Rev., 247, 104621, https://doi.org/10.1016/j.earscirev.2023.104621, 2023.
Tobin, T. S., Wilson, G. P., Eiler, J. M., and Hartman, J. H.: Environmental change across a terrestrial Cretaceous-Paleogene boundary section in eastern Montana, USA, constrained by carbonate clumped isotope paleothermometry, Geology, 42, 351–354, https://doi.org/10.1130/G35262.1, 2014.
Tshimanga, R. M. and Hughes, D. A.: Climate change and impacts on the hydroclimate of the Congo Basin: The case of the northern sub-basins of the Oubangui and Sangha Rivers, Phys. Chem. Earth, 50–52, 72–83, https://doi.org/10.1016/j.pce.2012.08.002, 2012.
Van Der Sleen, P., Groenendijk, P., Vlam, M., Anten, N. P. R., Boom, A., Bongers, F., Pons, T. L., Terburg, G., and Zuidema, P. A.: No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased, Nat. Geosci., 8, 24–28, https://doi.org/10.1038/ngeo2313, 2015.
Van Pul, P.: Hydrography and navigation on the Congo River: a century of visual history, Springer Nature Switzerland, https://doi.org/10.1007/978-3-031-41065-9, 2023.
Versteegh, E. A. A., Vonhof, H. B., Troelstra, S. R., and Kroon, D.: A molluscan perspective on hydrological cycle dynamics in northwestern Europe, Geol. Mijnbouw-N. J. G., 89, 51–60, https://doi.org/10.1017/S0016774600000810, 2010.
Versteegh, E. A. A., Vonhof, H. B., Troelstra, S. R., and Kroon, D.: Can shells of freshwater mussels (Unionidae) be used to estimate low summer discharge of rivers and associated droughts?, Int. J. Earth Sci., 100, 1423–1432, https://doi.org/10.1007/s00531-010-0551-0, 2011.
Vonhof, H. B., Wesselingh, F. P., and Ganssen, G. M.: Reconstruction of the Miocene western Amazonian aquatic system using molluscan isotopic signatures, Palaeogeogr. Palaeocl., 141, 85–93, https://doi.org/10.1016/S0031-0182(98)00010-8, 1998.
Vonhof, H. B., Wesselingh, F. P., Kaandorp, R. J. G., Davies, G. R., Van Hinte, J. E., Guerrero, J., Rasanen, M., Romero-Pittman, L., and Ranzi, A.: Paleogeography of Miocene Western Amazonia: isotopic composition of molluscan shells constrains the influence of marine incursions, Geol. Soc. Am. Bull., 115, 983–993, https://doi.org/10.1130/B25058.1, 2003.
Wesselink, A. J., Orange, D., Feizoure, C. T., and Randriamiarisoa: Les régimes hydroclimatiques et hydrologiques d'un bassin versant de type tropical humide: l'Oubangui (République Centrafricaine), L'hydrologie tropicale: géoscience et outil pour le développement (Actes de la conférerice de Paris, mai 1995), IAHS Publ. 238, ISBN 0-947571-99-X, 179–194, 1996.
Zhou, L., Tian, Y., Myneni, R. B., Ciais, P., Saatchi, S., Liu, Y. Y., Piao, S., Chen, H., Vermote, E. F., Song, C., and Hwang, T.: Widespread decline of Congo rainforest greenness in the past decade, Nature, 508, 86–90, https://doi.org/10.1038/nature13265, 2014.