the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 15 Feb 2021
| 15 Feb 2021
The transformation of the forest steppe in the lower Danube Plain of southeastern Europe: 6000 years of vegetation and land use dynamics
Angelica Feurdean
Roxana Grindean
Gabriela Florescu
Ioan Tanţău
Eva M. Niedermeyer
Andrei-Cosmin Diaconu
Simon M. Hutchinson
Anne Brigitte Nielsen
Tiberiu Sava
Andrei Panait
Mihaly Braun
Thomas Hickler
Forest steppes are dynamic ecosystems, highly susceptible to changes in climate, disturbances and land use. Here we examine the Holocene history of the European forest steppe ecotone in the lower Danube Plain to better understand its sensitivity to climate fluctuations, fire and human impact, and the timing of its transition into a cultural forest steppe. We used multi-proxy analyses (pollen, n-alkanes, coprophilous fungi, charcoal and geochemistry) of a 6000-year sequence from Lake Oltina (southeastern Romania) combined with a REVEALS (Regional Estimates of Vegetation Abundance from Large Sites) model of quantitative vegetation cover. We found a greater tree cover, composed of xerothermic (Carpinus orientalis and Quercus) and temperate (Carpinus betulus, Tilia, Ulmus and Fraxinus) tree taxa, between 6000 and 2500 cal yr BP. Maximum tree cover (∼ 50 %), dominated by C. orientalis occurred between 4200 and 2500 cal yr BP at a time of wetter climatic conditions and moderate fire activity. Compared to other European forest steppe areas, the dominance of C. orientalis represents the most distinct feature of the woodland's composition at this time. Tree loss was underway by 2500 yr BP (Iron Age), with the REVEALS model indicating a fall to ∼ 20 % tree cover from the Late Holocene forest maximum, linked to clearance for agriculture, while climate conditions remained wet. Biomass burning increased markedly at 2500 cal yr BP, suggesting that fire was regularly used as a management tool until 1000 cal yr BP when woody vegetation became scarce. A sparse tree cover, with only weak signs of forest recovery, then became a permanent characteristic of the lower Danube Plain, highlighting more or less continuous anthropogenic pressure. The timing of anthropogenic ecosystem transformation here (2500 cal yr BP) falls between that in central-eastern (between 3700 and 3000 cal yr BP) and eastern (after 2000 cal yr BP) Europe. Our study is the first quantitative land cover estimate at the forest steppe ecotone in southeastern Europe spanning 6000 years. It provides critical empirical evidence that, at a broad spatial scale, the present-day forest steppe and woodlands reflect the potential natural vegetation in this region under current climate conditions. However, the extent of tree cover and its composition have been neither stable in time nor shaped solely by the climate. Consequently, vegetation change must be seen as dynamic and reflecting wider changes in environmental conditions including natural disturbances and human impact.
- Article
(5782 KB) - Full-text XML
-
Supplement
(612 KB) - BibTeX
- EndNote





Projected changes in climate and increasing human environmental impacts are generating global concern about the functioning of ecosystems as well as the provision of ecosystem services (IPCC, 2014; IPBES, 2019). Lowland ecosystems (mesic and steppic grasslands, woodlands, etc.) provide an array of provisioning (e.g. crops, grazed areas and wood) and regulating services (e.g. soil protection; European Environmental Agency, 2016). However, in comparison to the mountainous areas of central-eastern Europe, lowland ecosystems, especially steppic grasslands, have been more strongly impacted by human activities (Magyari et al., 2010; Tonkov et al., 2014; Feurdean et al., 2015; Kuneš et al., 2015; Novenko et al., 2016; Shumilovskikh et al., 2018, 2019; Jamrichová et al., 2019; Vincze et al., 2019; Cleary et al., 2019; Gumnior et al., 2020). This partly reflects the lowlands' deeper and more fertile soils and the greater accessibility of the terrain, which have promoted extensive agro-pastoral activities and human settlement. Lowlands also include more frequent ecotones, i.e. woodland and grassland borders, which are naturally more sensitive to climate change (Bohn et al., 2003).
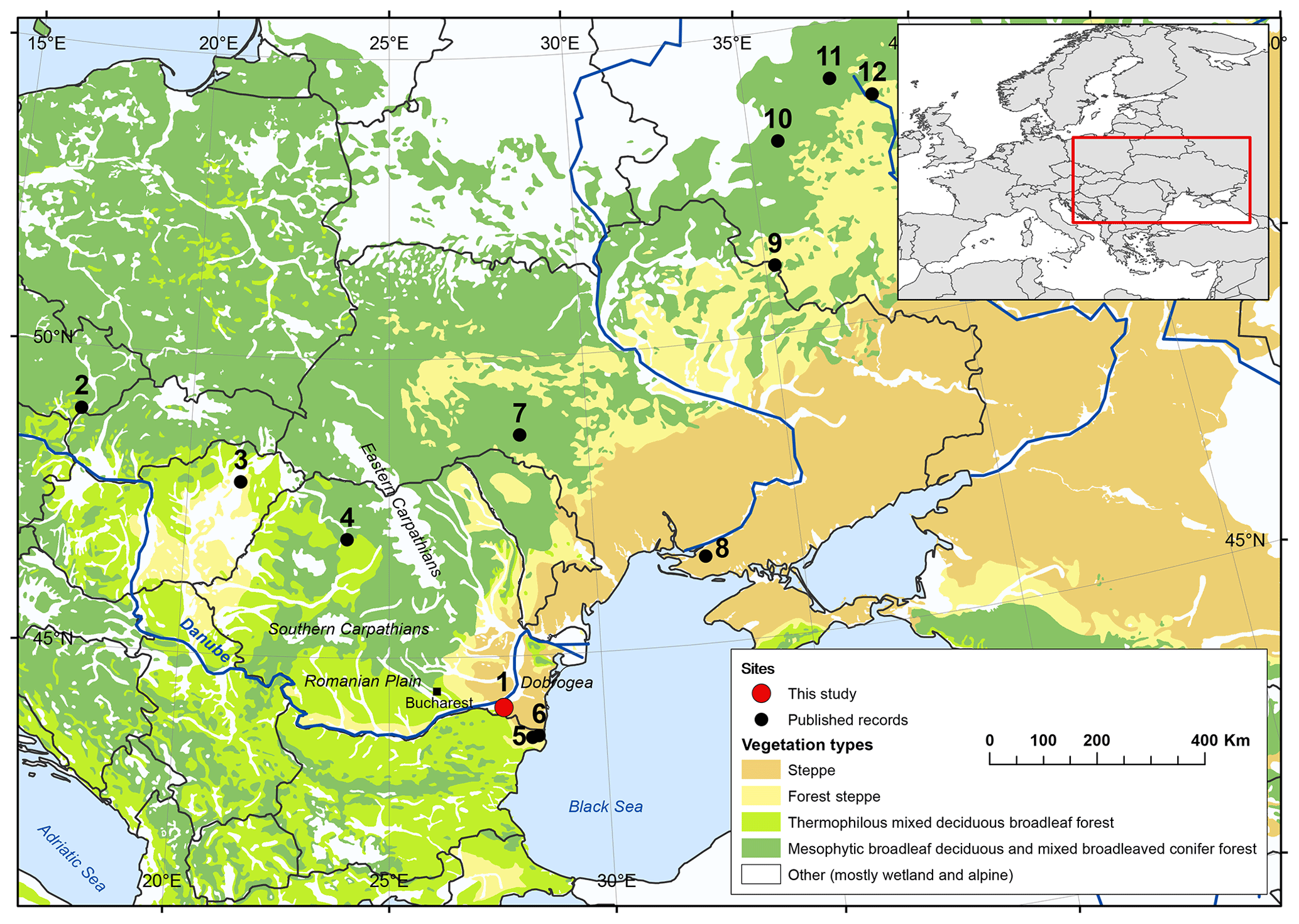
Figure 1Potential natural vegetation cover in Europe showing the extent of the European forest steppe region (after Bohn, 2003; © BfN, Bundesamt für Naturschutz), the location of the study site and the other published records used for comparisons. 1: Lake Oltina (study site); 2: Lake Vracov; 3: Sarló-hát; 4: Lake Stiucii; 5: Durankulak 2; 6: Durankulak 3; 7: Dovjok; 8: Kardashinski; 9: Sudzha; 10: Selikhovo; 11: Istochek; 12: Podkosmovo. For references see Table 3.
According to Bohn et al. (2003), the potential natural vegetation (PNV) in the study area, the easternmost part of the lower Danube Plain, also known as the southern Dobrogea Plateau, is forest steppe, i.e. woodland patches within a matrix of graminoid- and forb-dominated communities. It borders steppe grasslands i.e. treeless vegetation cover dominated by graminoids and forbs to the east. Forest steppe and steppe vegetation extend over 6000 km along an east–west gradient across Eurasia (Fig. 1) under climate conditions delimited by a ca. 2-month long late-summer drought for the forest steppe zone and a 4–6 month one for the steppe (Walter, 1974). However, according to the management plan for the region (Planul de Management, 2016), there are currently few patches of natural steppe vegetation preserved in the southern Dobrogea Plateau, as most have become ruderal steppe. Since the lower Danube Plain represents one of the oldest areas of continuous human occupation from the Neolithic onwards, i.e. 8000 cal yr BP (http://ran.cimec.ro (last access: 28 May 2019); Bălăşescu and Radu, 2004; Weininger et al., 2009; Wunderlich et al., 2012; Nowacki et al., 2019; Preoteasa et al., 2018), and one of the most important agricultural areas in Europe (European Environmental Agency, 2016), the current vegetation is likely very different from its natural state, though it is not known exactly how different it is. The use of the PNV as a baseline for natural vegetation has been found to be problematic, as it considers only climax vegetation, i.e. the final stage of an ecological succession, and assumes that vegetation remains static in space and time (Chiarucci et al., 2010; Jackson et al., 2013; Abraham et al., 2016; Rull, 2015). The PNV also fails to consider the vital role of natural disturbances such as fire and herbivores and their impacts on vegetation succession (Chiarucci et al., 2010; Feurdean et al., 2018), leading to unrealistic vegetation reconstruction even in the absence of disturbance by humans (Jackson et al., 2013). As a consequence, many areas that are currently covered by grasslands or open woodlands in central-eastern Europe are defined as naturally dominated by deciduous broadleaf forest or mixed coniferous and broadleaf forest as the PNV (Feurdean et al., 2018). Additionally, the representation of pioneer trees is much lower in the PNV than suggested by pollen-based Holocene vegetation estimates (Abraham et al., 2016). Significantly, the inaccurate identification of natural vegetation types can lead to inappropriate decision making in terms of conservation practices and policies. For example, the Global Partnership on Forest and Landscape Restoration use the PNV to identify opportunities for landscape restoration to mitigate climate change in areas where the climate can sustain forest (http://www.wri.org/applications/maps/flr-atlas, last access: 20 May 2019). However, this approach threatens grassland ecosystems, as such policies are based on the false assumption that most grasslands are man-made and ignores their richness at a smaller spatial scale (Whittaker et al. 2001), as well as their unique cultural significance (Dengler et al., 2014).
Palaeoecological records provide a way to assess the former natural vegetation of a region and the legacy of natural disturbances and anthropogenic impacts on landscapes (Willis and Birks, 2006). However, due to the dry climate of the lower Danube Plain, very few palaeoecological archives are available to document past natural vegetation types. This leaves many open questions regarding the temporal dynamics of vegetation composition and drivers of changes in this region and how its vegetation composition compares to other forest steppe areas in central-eastern Europe (Magyari et al., 2010; Feurdean et al., 2015; Kuneš et al., 2015), southeastern Europe (Tonkov et al., 2014; Marinova and Atanassova, 2006) and the Eastern European Plain (Novenko et al., 2016; Shumilovskikh et al., 2018). Most of the archaeological and loess deposits in the region are devoid of an absolute chronology, have poor lithological context and lack favourable pollen preservation (Tomescu, 2000). Nevertheless, based on these fragmentary records, it appears that steppe may have covered the landscapes of this region in the Early Holocene, while forest steppe vegetation may have expanded during a moist phase of the Mid Holocene (Feurdean et al., 2014; Tomescu, 2000; Wunderlich et al., 2012; Hansen et al., 2015). In addition, models of deforestation rates, using a scenario that accounts for population history and technological advances, suggest that the extent of deforestation in the lower Danube basin has increased continuously since 4000 cal yr BP (Kaplan et al., 2009; Giosan et al., 2012). Indirect evidence for the Holocene persistence of steppe grasslands in this region comes from the genetic investigation of steppe species including Adonis vernalis, Astragalus exscapus, Stipa capillata, and S. pulcherrima, which mostly show a unique genetic diversity reflecting the continuous occurrence and limited past migration of these steppe elements within Europe (Kropf et al., 2020; Plenk et al., 2020).
Here, we explore the long-term vegetation dynamics of the lower Danube Plain's landscape and the competing driving forces (climate, fire and anthropogenic impact). More specifically, we address the following research questions:
- i.
Is forest steppe the natural vegetation type of the lower Danube Plain under climatic conditions similar to those in the present?
- ii.
Has the tree cover been more extensive or dominated by other tree taxa in the past?
- iii.
When did this area undergo the most marked land cover and land use changes, and was this transformation continuous in time?
This study is built on a pollen-based quantitative vegetation reconstruction (REVEALS model; Regional Estimates of Vegetation Abundance from Large Sites), along with records of long-chain higher-plant wax n-alkanes, macro-charcoal, coprophilous fungi and geochemistry from a sedimentary sequence retrieved from Lake Oltina, southeastern Romania. This is the first pollen-based quantitative land cover estimate in the forest steppe of southeastern Europe. It allows for the hypothesis of the naturalness of forest steppe ecosystems in this region, as well as its sensitivity to climate and anthropogenic impact, to be critically tested (i.e. see Harrison et al., 2020).
2.1 Geography, climate and vegetation
Lake Oltina (44∘09′16′′ N, 27∘38′13′′ E, 7 m a.s.l.) is located on the floodplain of the Danube River, the lower Danube Plain, in southeastern Romania (Fig. 1). It is the largest fluvial lake on the Danube floodplain with a surface area of 33 km2 and is part of a natural reserve and a Natura 2000 site (ROSPA0056; Management Plan 2016). The lake has a major tributary, Canaraua Fetii, and is connected to two smaller lakes, Ceamurlia and Iortmac, located upstream as well as to the Danube River via an artificial dam built to prevent flooding (Telteu, 2014). The climate in the study region is wet warm-temperate continental (Köppen–Geiger class Dfa), also termed excessive, with the prevalence of harsh winters and hot summers (Posea, 2005), due to the influence of air masses from continental Asia. The mean annual temperature is ca. 11 ∘C, with a mean January temperature of ca. −1 ∘C and a mean summer temperature of 25 ∘C. Annual precipitation is about 400 mm (Adamclisi meteorological station). The geology of the catchment comprises limestone and loess deposits, whereas the soils are represented mainly by Haplic and Luvic Chernozem and Phaeozem (IUSS WRB 2006). Friable steep loess deposits are frequently eroded, delivering a clastic sedimentary influx into the lake (Romanescu et al., 2010).
The main potential vegetation types of the region include forest, forest steppe and calciphile steppe (Bohn at al., 2003). The main forest types are: thermophilous mixed deciduous broadleaf forests (subtypes G21, G22 and G34 according to Bohn et al., 2003), mesophytic deciduous broadleaf forests (subtypes F49 and F67), forest steppe (subtype L13) and steppe (subtype M5). Additionally, alluvial forests (subtypes U19 and U20), halophytic vegetation (P33) and tall-reed vegetation and sedge swamps (R1) prevail (Bohn et al., 2003). CORINE (Coordination of Information on the Environment) Land Cover data (2012) indicate that the present land cover within a 20 km radius of the lake comprises ca. 65 % arable land and orchards, 19 % steppe and semi-natural grassland, and 16 % deciduous forest (S1; Grindean et al., 2019). Important tree species in forests within this radius, and thus the most relevant for the pollen source area, are Quercus pubescens, Q. pedunculiflora, Q. robur, Q. cerris, Q. virgiliana, Carpinus orientalis, Acer tataricum, Tilia tomentosa, Fraxinus excelsior and Ulmus minor, whereas hygrophilous tree taxa, growing along the Danube River, are represented by Populus nigra, P. alba, Salix alba and S. fragilis. Shrubs occur abundantly either as understory vegetation or as thickets and are composed of Fraxinus ornus, Cotinus coggygria, Prunus mahaleb, P. spinosa, P. cerasifera, Rosa canina, Pyrus communis, Crataegus monogyna, Amorpha fruticosa, Gleditsia triacanthos, Elaeagnus angustifolia and Ailanthus altissima. Natural grassland steppic species are common on calcareous slopes and include Stipa stenophylla, S. ucrainica, S. capillata, Poa angustifolia, Festuca valesiaca, Bothriochloa ischaemum, Artemisia austriaca, Agrostis tenuis, Carex humilis, Centaurea orientalis, Astragalus ponticus and Thymus marschallianus (Sârbu et al., 2009; Grindean et al., 2019).
3.1 Core collection, lithology and chronology
Sediment cores were extracted with a Livingstone piston corer (1 m long and 5 cm diameter) from the central part of the lake (1.8 m water depth) in spring 2016. The less consolidated sediment at the surface (36 cm) had previously been retrieved with a gravity corer in 2014. A lithostratigraphic description was made according to changes in texture, colour, magnetic susceptibility and the organic carbon content (loss on ignition; LOI).
Volume magnetic susceptibility (κvol) was measured with a Bartington Instruments Ltd MS2 meter and E sensor (Bartington Instruments, 2008). For LOI, samples were dried overnight at 105 ∘C and combusted for 5 h at 550 ∘C and then for 2 h at 900 ∘C, and LOI is expressed as percentage loss of the dry weight (Heiri et al., 2001). A composite sedimentary core record totalling 964 cm was constructed using the uppermost 10 cm from the gravity core and 10 overlapping Livingstone core sections. To determine the erosion and anoxic conditions in the lake, elemental geochemical concentration was measured on 190 sediment samples (extracted at ∼ 5 cm intervals along the core) and subsequently dried and homogenized using a non-destructive Niton XL3t 900 x-ray fluorescence analyser (FPXRF). An NCS DC73308 was employed as a certified reference material (CRM). Measurement follows the procedure described by Hutchinson et al. (2015). We selected the detrital element Zr as a proxy for erosion (Kylander et al., 2011) and employed the Fe : Mn ratio to reconstruct anoxic conditions in the lake (Nacher et al., 2013).
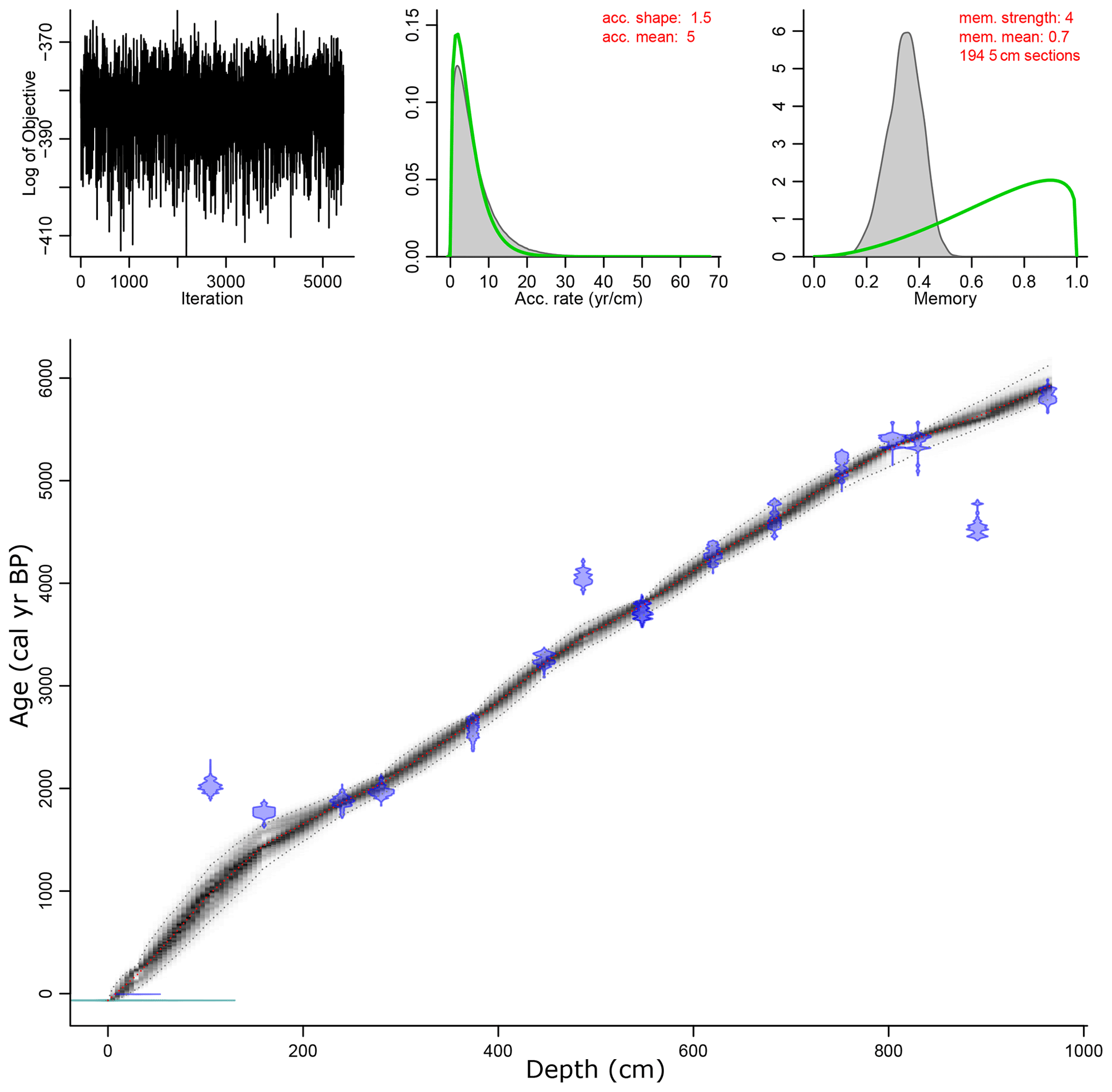
Figure 2Age–depth model for Lake Oltina using a Bayesian approach (for more details, see Appendix A1). Accumulation: acc.; memory: mem.
Table 1AMS (accelerator mass spectrometry) 14C measurements at Lake Oltina.
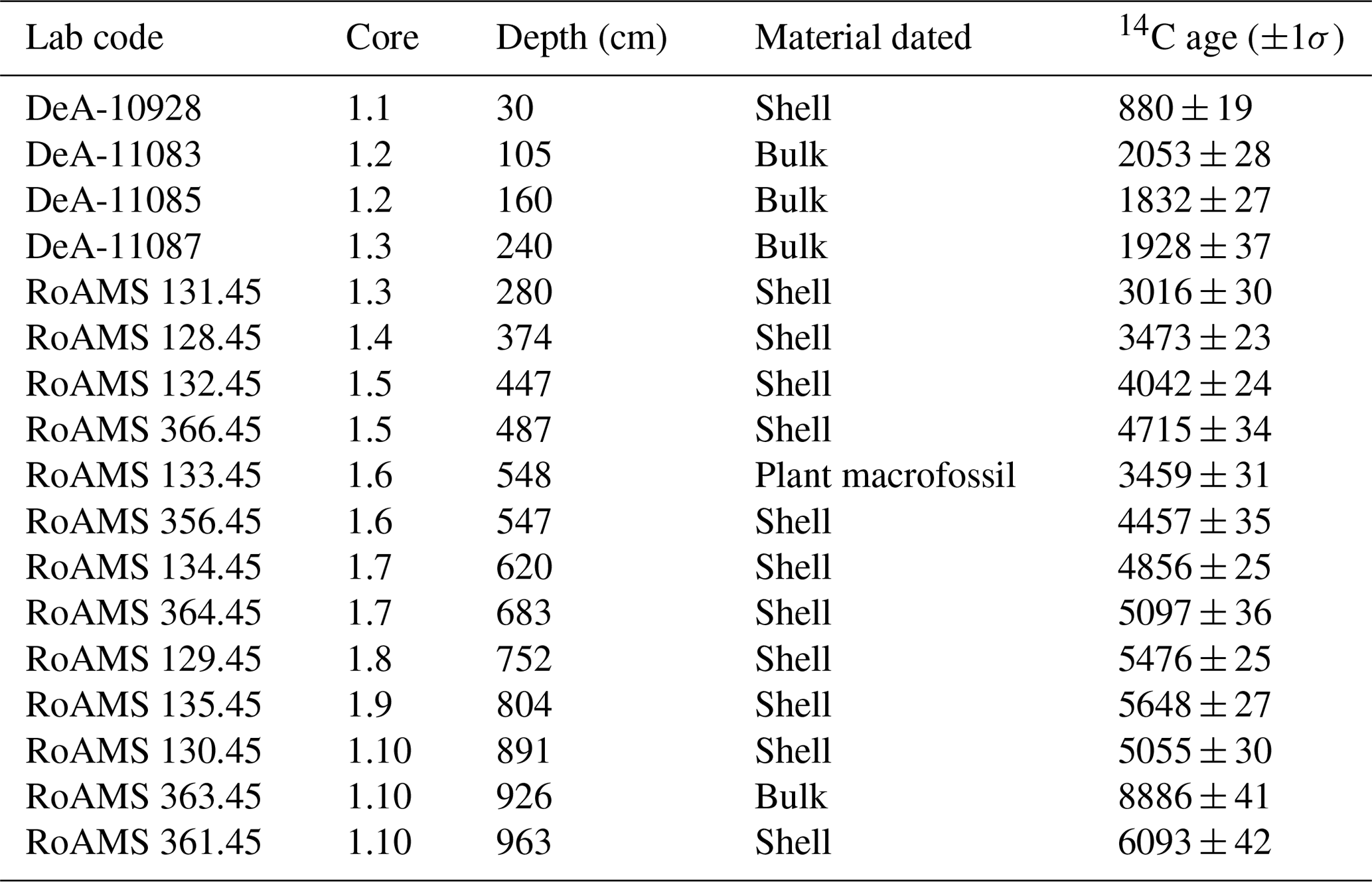
The chronology was established based on 18 AMS (accelerator mass spectrometry) 14C measurements (Table 1). Attempts to constrain the chronology of the top core via 210Pb and 137Cs gamma assay measurements failed to produce any meaningful results, probably due to surface sediment mixing. An attempt to establish a tephra-based chronology also failed due to a poor match to any known volcanic eruptions. The radiocarbon age estimates were converted into calendar years BP via the Bacon software (Blaauw and Christen, 2011) using the IntCal13 data set of Reimer et al. (2013). An age–depth curve was derived based on a smoothing spline model (Fig. 2; for details on age–depth construction, see Fig. A1 in the Appendix).
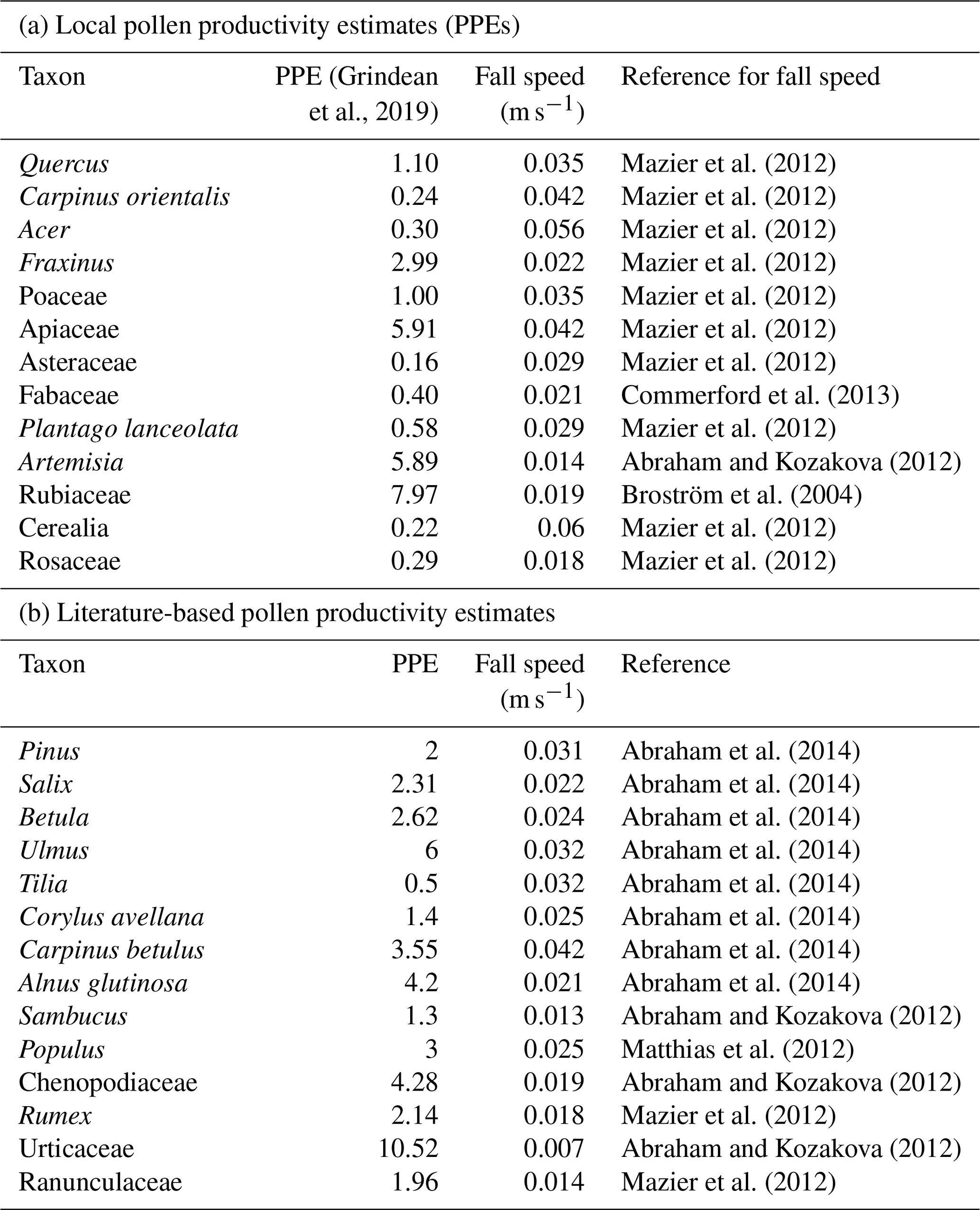
Table 2Pollen productivity estimates relative to Poaceae and their respective fall speeds used in the REVEALS model.
∗ Standardized after Mazier et al. (2012).
3.2 Vegetation reconstruction
3.2.1 Pollen-based quantitative reconstruction of land cover using the REVEALS model
To determine the past vegetation cover we used pollen analysis on samples of 1 cm3 at intervals ranging between 5 and 10 cm (a total of 105 samples) along the composite core. Sediment preparation followed the protocol of Goeury and de Beaulieu (1979). We identified the pollen grains using the atlases of Reille (1995, 1999). A minimum of 300 terrestrial pollen grains were counted at each level and used to calculate the pollen percentages. We corrected for biases in taxon-specific pollen productivities and dispersal and thereby produced a quantitative reconstruction of the vegetation cover in the region surrounding Lake Oltina using the Regional Estimates of Vegetation Abundance from Large Sites, i.e. REVEALS model (Sugita, 2007). In this model, we used pollen productivity estimates (PPEs) for the most characteristic plant taxa from the studied region. For 13 plant taxa that include five woody and eight herbaceous and shrub taxa, we used PPEs measured in this particular region (Grindean et al., 2019). We have complemented these with literature-based PPEs for 14 additional taxa that significantly contribute to the regional vegetation composition (Table 2). We used the Sugita (2007) dispersal model with default settings for neutral atmospheric conditions and wind speed (3 m s−1). The literature-based fall speed of each pollen type (Table 2) is used to model dispersal. The spatial extent of the regional vegetation is set at a 100 km radius. The vegetation cover reconstructed using REVEALS always adds up to 100 %, which means that taxa not included in the model, as well as non-pollen-producing areas, are ignored. The 27 taxa selected for our REVEALS model represent between 77 and 95 % in the terrestrial pollen sum. Significant changes in the vegetation assemblages were defined using stratigraphically constrained cluster analyses (incremental sum of squares method) of REVEALS-based vegetation cover percentages in Tilia (Grimm, 2004).
3.2.2 Vegetation reconstruction based on leaf wax n-alkanes
To determine the source of organic matter and the predominant vegetation type (Eglinton and Calvin, 1967; Ficken et al., 2000; Diefendorf et al., 2015), we measured the concentration of higher-plant-derived n-alkane homologues of 60 sediment samples selected along the composite core. n-Alkanes are an integral part of higher-plant leaf epicuticular waxes, highly resistant to degradation and among the most stable lipid components of the protective waxes coating terrestrial plant leaves (Eglinton and Eglinton, 2008; Sachse et al., 2012). They are commonly used to distinguish sources of organic matter based on their chain length (see below). n-Alkanes were extracted from freeze-dried and finely ground sediment samples (ca. 1 g dry weight) with a Büchi SpeedExtractor at 75 ∘C and 100 bar using 20 mL of a mixture of dichloromethane / methanol (9:1) for 10 min, which was repeated three times. Total lipid extracts (TLEs) were dried under a stream of N2 at 36 ∘C. The apolar fraction containing n-alkanes was eluted from the TLE by silica-gel column chromatography using hexane. n-Alkanes were subsequently isolated from the apolar fraction using urea adduction (Vasiliev et al., 2013). n-Alkane homologues were separated and quantified by gas chromatography–mass spectrometry (GC–MS) using a Thermo Finnigan Trace GC equipped with a HP-5MS column (30 m × 0.25 mm × 0.25 µm) connected to a Thermo Finnigan DSQ II mass spectrometer. The GC oven was held at 70 ∘C for 1 min and ramped at 10 ∘C min−1 to a final temperature of 280 ∘C, which was held for 15 min. n-Alkanes were identified by comparison of their mass spectra and retention time to an external standard (n-C7 to n-C40; Supelco) at a concentration of 25 ng µL. They were quantified using total ion chromatogram peak areas calibrated against the external standard. Precision of the quantification is 96 % as inferred from the standard deviation of repeated standard runs (n=5). Concentrations of individual n-alkanes were expressed as ng µL−1 dry weight of sediment.
In this study, we calculated the ratio of straight-chain n-alkanes of different chain lengths (homologues), as these have been previously used as proxies for the relative contribution of various types of plants in lacustrine sediments (e.g. Ficken et al., 2000; Zhou et al., 2005). Average chain length (ACL) is an indicator of the relative abundance of short- (C16–C20) vs. long-chain n-alkanes and may be linked to the predominance of higher taxonomic plants over lower taxonomic plants (Ficken et al., 2000; Eglinton and Calvin, 1967). Within the long-chain n-alkanes, the abundance of n-alkanes with n-C31 and n-C33 may be indicative of grass predominance, whereas n-C27 and n-C29 may indicate a predominantly tree-covered landscape (Aichner et al., 2010; Meyers, 2003). The aquatic index (Paq) quantifies the abundance of submerged and floating vascular macrophytes, which are characterized by medium-chain-length n-alkanes, relative to emergent plant types that are characterized by long-chain n-alkanes (Ficken et al., 2000). It should be noted that n-alkanes are less successful in detecting coniferous than angiosperms (Diefendorf et al., 2015) and that some overlap within the medium-chain-length alkanes n-C23 and n-C25 is possible (Aichner et al., 2010; Meyers, 2003).
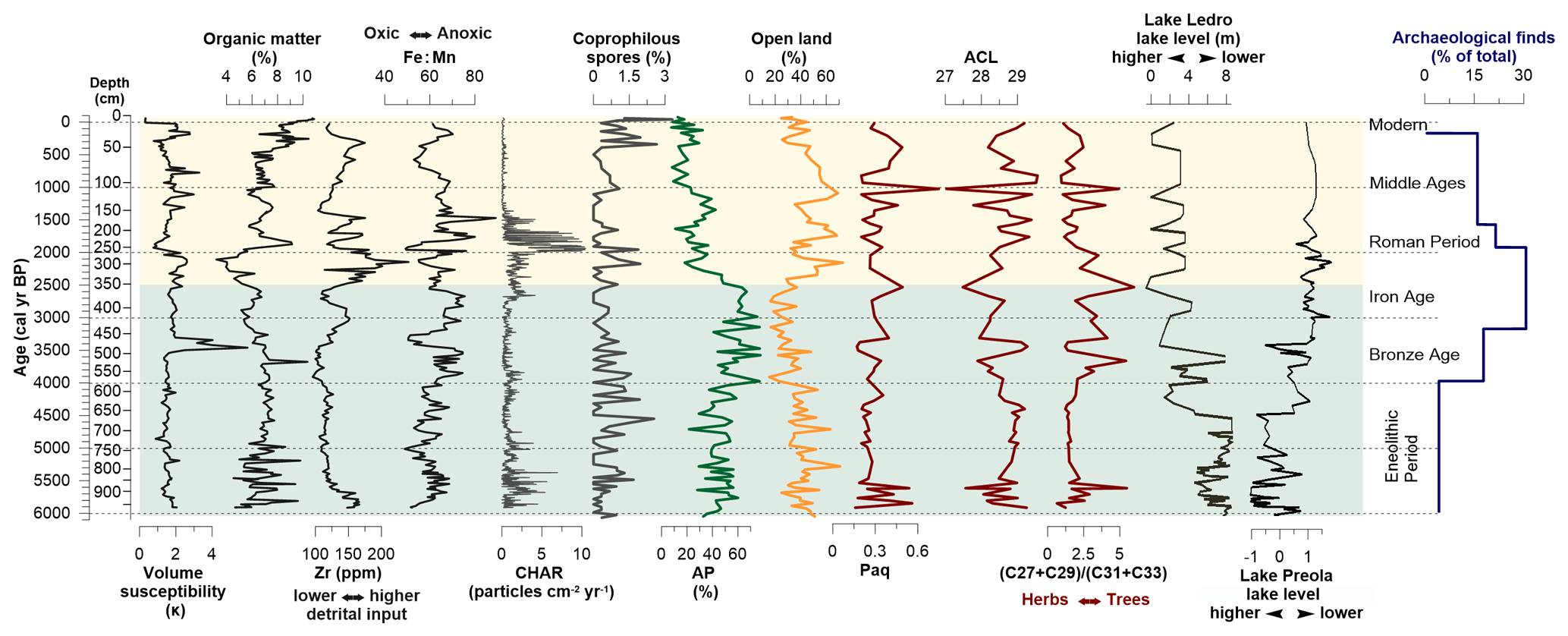
Figure 3Integrative diagram showing in-lake ecosystem and catchment changes. Lake properties: volume magnetic susceptibly, organic matter content and detrital elements (Zr and Fe : Mn ratio). Biomass burning as determined from the macro-charcoal (CHAR) record and grazing activity from coprophilous spores. Landscape cover determined from the arboreal-pollen (AP) and the open-land pollen percentages; ACL (average chain length) of higher plant and the ratio of (n-C31 + n-C33) (n-C27 + n-C29) as a proxy for abundance of higher plants, and herbs vs. trees predominance in the landscape, respectively. Regional lake level fluctuations at Lake Ledro, northern Italy (45∘ N), and Lake Preola, southern Italy (37∘ N) (Magni et al., 2013). Archaeological finds include settlements and cemeteries within a 20 km radius of Lake Oltina taken from the Repertoriul Arheologic National (National Archaeological Record of Romania; http://ran.cimec.ro, last access: 28 May 2019).
The n-alkane proxies were calculated using the following equations:
Analysis of the composition of lipid compounds in modern river sediment deposits along the Danube River shows a predominantly local source of proxies derived from branched glycerol dialkyl glycerol tetraethers (Freymond et al., 2017). In line with this finding, we presume that changes in n-alkane homologue abundance in our record integrate vegetation changes not only in the lake and near the lake but also more regionally in the lake catchment.
3.3 Regime disturbances by fire and herbivores
To determine past disturbance by fire, macroscopic charcoal particles were counted in samples of 2 cm3 extracted at 1 cm contiguous intervals. Samples were bleached, wet-sieved through a 150 µm mesh and identified under a stereomicroscope following the methodology described in Feurdean et al. (2017a). Here we report only the results for total macro-charcoal particles. We calculated the macro-charcoal accumulation rate (CHAR, particles cm−2 yr−1) by dividing the total macro-charcoal concentration by sediment deposition time (yr cm−1). To determine past grazing activity, coprophilous fungi (Sporormiella, Sordaria and Podospora) were tallied during routine pollen counting (van Geel et al., 1980; Baker et al., 2013). Percentages of coprophilous fungi were determined by adding their own sum to the total terrestrial pollen sum.
4.1 Chronology and sediment composition
The lithology of the core at Lake Oltina showed little variability throughout the profile and comprises clay, gyttja clay and sandy clay. The age–depth model indicates a rather constant sediment accumulation rate with a mean of 5 yr cm−1 and no evidence of hiatuses (Figs. 2; A1). Organic matter (OM) as determined from LOI at 550 ∘C varied between 3 % and 10 % with slightly higher values between 5000 and 3500 cal yr BP and over the past 2000 years (Fig. 3). Our selected geochemical detrital element, Zr, a proxy for erosion, showed the lowest values between 5500 and 3500 cal yr BP (up to 150 ppm) and greater, highly fluctuating values at the beginning of the record and over the 2500 years (Fig. 3). The Fe:Mn ratio, a proxy for anoxic conditions in the lake, generally displayed large variability throughout the profile, although values were higher between 4000 and 3500 cal yr BP, around 3000 cal yr BP, and between 2000 and 1500 cal yr BP (Fig. 3).
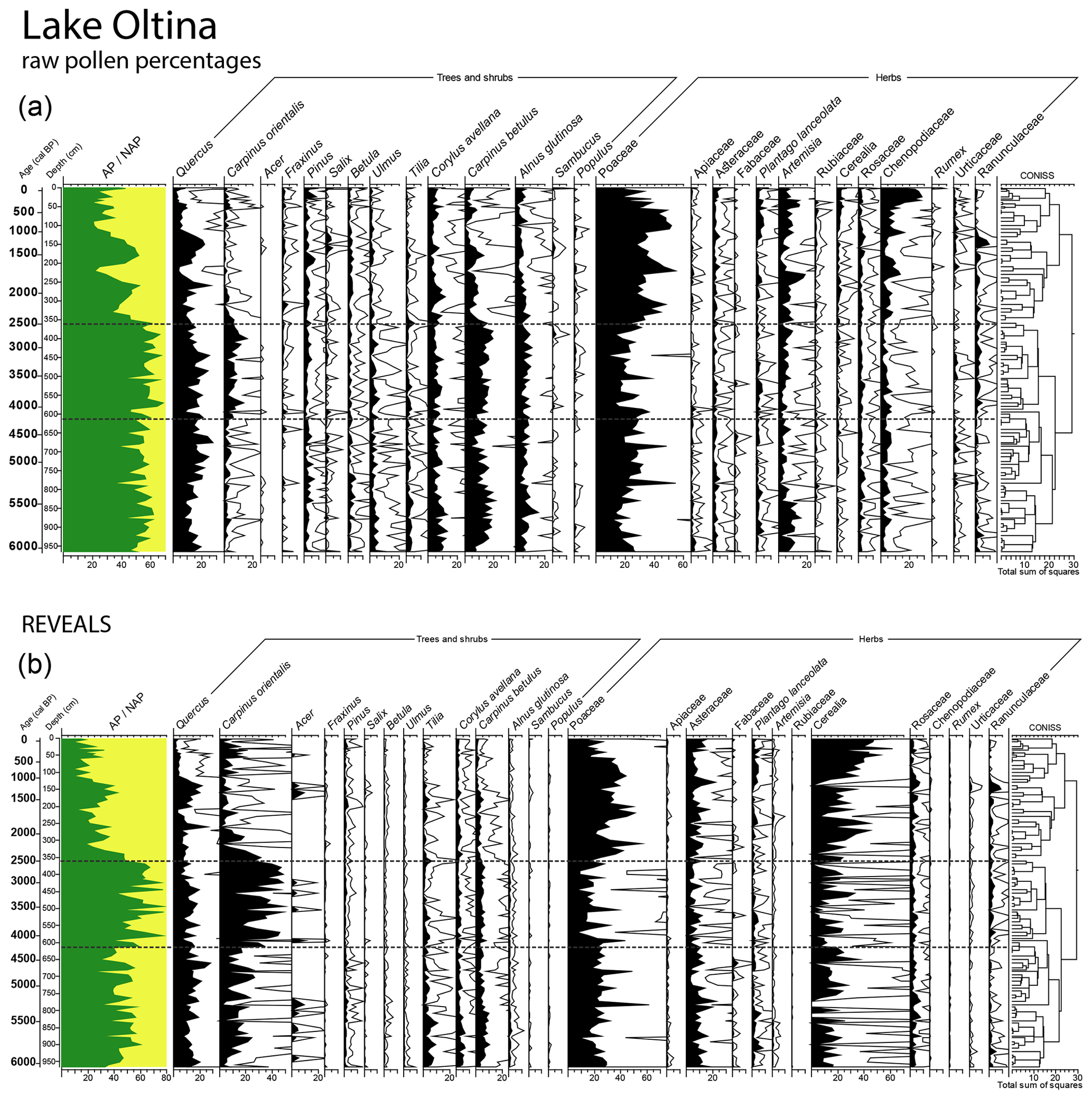
Figure 4Raw pollen percentages and estimated regional vegetation cover based on the REVEALS model for 27 taxa including trees, shrubs and herbs at Lake Oltina. Horizontal lines denote the timing of the most important changes in the vegetation assemblages. NAP: non-arboreal pollen; CONISS: stratigraphically constrained cluster analysis.
4.2 Landscape reconstruction from pollen
The cluster analysis applied on the pollen record indicated three major periods of change in land cover and vegetation openness over the last 6000 years: open temperate and xerothermic deciduous broadleaf forest between 6000 and 4200 cal yr BP, the maximum extent of broadleaf tree cover between 4200 and 2500 cal yr BP, and the expansion of grassland between 2500 cal yr BP and the present (Fig. 4). Results from the REVEALS model suggest that landscape openness was ca. 10 %–15 % greater than the estimates derived from the raw pollen data. Overall, REVEALS indicate a greater proportion of Carpinus orientalis, Tilia, Acer, Rosaceae, Cerealia and Asteraceae and a lower proportion of Corylus avellana, Betula, Ulmus, Alnus, Fraxinus, Salix, Artemisia and Chenopodiaceae than the raw pollen data (Fig. 4). Quercus, Plantago lanceolata and Poaceae show a largely similar abundance in both the raw data and REVEALS estimates.
4.2.1 6000–4200 cal yr BP: open temperate and xerothermic deciduous broadleaf forest or forest steppe
The REVEALS estimate of tree cover fluctuated around 40 %, compared to ∼ 55 % in the raw pollen percentages, and was primarily represented by Carpinus orientalis, Quercus, Carpinus betulus, Corylus avellana, Tilia and Ulmus (Fig. 4). The REVEALS model predicts an almost equal proportion of total forb (Artemisia, Chenopodiaceae, Asteraceae, Rosaceae, Brassicaceae, Plantago major, Thalictrum and Caryophyllaceae) and grass (Poaceae) in the herbaceous cover (Fig. 4). However, in the raw pollen percentages, forbs dominate the herbaceous assemblages (30 %), whereas Poaceae constitutes ∼ 20 % (Figs. 4; A2). The Cerealia cover estimate fluctuates around ∼ 20 % in the REVEALS model and was below 5 % in the raw pollen percentages. CHAR values were high between 6000 and 5000 cal yr BP and declined markedly thereafter (Fig. 3). The abundance of coprophilous fungi (Podospora, Sordaria and Sporormiella), on the other hand, rose between 5000 and 4000 cal yr BP (Figs. 3; A2).
4.2.2 4200–2500 cal yr BP: maximum extent of temperate and xerothermic deciduous broadleaf tree cover
Tree cover increased to its maximum extent in the profile (fluctuating around 55 %) and was mostly represented by Carpinus orientalis and Quercus with some occurrence of Carpinus betulus, Corylus avellana and Tilia (Fig. 4). The rise in Carpinus orientalis abundance is more evident in the REVEALS reconstruction (40 %) than in the raw pollen percentages (20 %). In the herbaceous cover, Poaceae declined the most, but there were no marked changes in pollen of primary anthropogenic indicators (Cerealia; Fig. 4). Both the abundance of CHAR and of coprophilous fungi decreased to one of the lowest in the profile (Fig. 3).
4.2.3 2500–0 cal yr BP: decline in Carpinus orientalis–Quercus tree cover and the expansion of grassland and pasture and arable cover
The tree cover declined abruptly to ca. 20 %; this was most evident for Carpinus orientalis, decreasing from 40 % to 10 % (Fig. 4). However, tree cover fluctuated strongly over the last 2500 years, with intervals of lower values (20 %) between 2500 and 1700 cal yr BP and over the last 1000 years and of increases (30 %) between 1700 and 1000 cal yr BP (Fig. 4). The REVEALS estimate also suggests an increased proportion of grass (Poaceae), cultivated cereals (Cerealia and Secale cereale) and forbs. Among forbs, ruderal taxa Asteraceae, Plantago lanceolata and Rosaceae showed the most visible increase (Figs. 4; A2). CHAR values increased gradually from the beginning of this time interval and attained a maximum in the profile between 2000 and 1700 cal yr BP, followed by the lowest values in the profile over the last 1000 years (Fig. 3). The abundance of coprophilous fungi was particularly elevated between 2500 and 2000 cal yr BP and over the last 1000 years.
4.3 n-Alkane-based lake catchment ecosystem changes
The (C27–C29) (C31–C33) ratio in Lake Oltina varied between 0.63 and 5.99 and showed the greatest values (2.84) between 4200 and 2000 cal yr BP and the lowest values between 5500 and 4200 cal yr BP (1.76) as well as over the past 2000 years (1.83; Fig. 3). The ACL varied between 27 and 30 and shows an opposite pattern to the (C27–C29) (C31–C33) ratio. Paq varied between 2 and 12 and showed the greatest values between 6000 and 5500, 4000 and 2500, and 1200 and 500 cal yr BP.
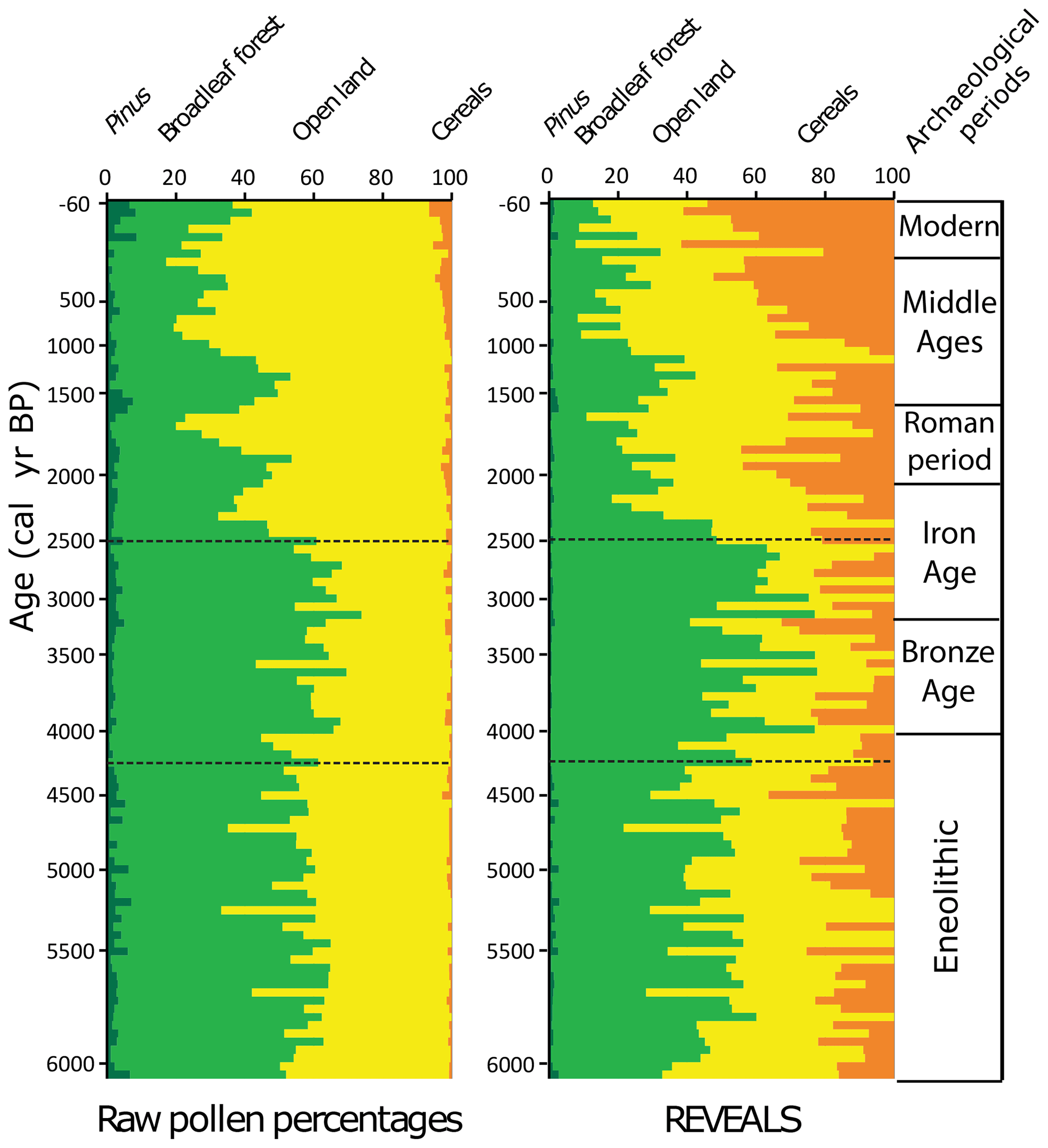
Figure 5Comparative summary of the percentage vegetation cover estimates based on the REVEALS model and raw pollen percentages at Lake Oltina. Open land cover includes all non-arboreal-pollen types, mostly indicators of pastures and grasslands. The Cerealia group includes Secale cereale, Triticum, Zea and Hordeum. Horizontal lines denote the timing of the most important changes in the vegetation assemblages. Archaeological finds as in Fig. 3.
5.1 Forest steppe and woodlands between 6000 and 2500 cal yr BP with a maximum tree cover between 4200 and 2500 cal yr BP
The pollen-based quantitative land cover reconstruction shows an average tree cover of 40 % between 6000 and 4200 cal yr BP and a tree cover maximum of 50 % between 4200 and 2500 cal yr BP in the surroundings of Lake Oltina (Figs. 4 and 5). In a pollen-based biome reconstruction, such a proportion of trees is likely to be indicative of a forest steppe or open-woodland type (Marinova et al., 2018). This woodland consists of tree taxa of xerothermic character including Quercus (likely Q. cerris and Q. pubescens) and Carpinus orientalis, along with temperate trees such as Carpinus betulus, Acer, Tilia, Ulmus and Fraxinus. Shrub (Rosaceae, Rosa canina, Sambucus, Prunus and Cornus), grass and forb communities were abundant and composed of a diverse mixture of mesophytic, xerothermic and halophilous taxa (Fig. A2). Coeval with the maximum extent in tree cover, the n-alkanes were dominated by shorter chain lengths (ACL) and a higher n(C27 + C29) (C33 C31) ratio, indicative of an increased contribution of tree-derived n-alkanes (Meyers, 2003; Aichner et al., 2010). However, the concentration of individual n-alkanes > C27 (not shown) varied with that of detrital element Zr. It was high between 6000 and 4200 cal yr BP and declined between 4200 and 2500 cal yr BP, suggesting a reduction in terrestrial plant delivery into the lake during the highest tree cover. The maximum extent of tree cover parallels a slight Fe : Mn ratio increase more evident between 4000–3500 and 3000–2500 cal yr BP. This may indicate the establishment of more anoxic conditions (Naeher et al., 2013), possibly associated with a higher lake level due to the intensification of Danube water and sediment discharge into the lake, a higher lake trophic status or less turbulent conditions. Increased anoxia linked to a higher lake trophic status and the decomposition of organic matter is supported by the slight increase of submerged aquatic macrophyte (Paq; Fig. 3). The presence of more aquatic plants in the lake at the time of the rise in the Fe : Mn ratio is also reflected in the increased abundance of aquatic and wetland taxa (Potamogeton, Myriophyllum and Typha/Sparganim; Fig. 4). On the other hand, low values of the lithogenic element Zr between 4200 and 2500 cal yr BP indicate more stable slope conditions with lower catchment run off, which might support the hypothesis that increased anoxia may have resulted from less turbulent conditions in the lake.
On a regional scale, the 6000–4200 cal yr BP interval was characterized by contrasting climate conditions north and south of 45∘ N latitude, due to differences in the dynamics of the storm tracks carrying moisture from the North Atlantic Ocean and Mediterranean Sea (Persoiu et al., 2017). Whilst southern Europe lake levels were low prior to 4500 cal yr BP, those in central Europe showed high stands during the same period (Magny et al., 2013). An opposite pattern is, however, visible after 4500 cal yr BP, when southern Europe lake levels increased, whereas those in central Europe declined. The lake level increase after 4500 cal yr BP also paralleled an intensification of fluvial activity in several rivers in southern Romania and the southern part of the Danube River at this time (Bozilova and Tonkov, 1998; Filipova-Marinova et al., 2007; Howard et al., 2004). Coastal lakes along the Black Sea were also at higher water levels at this time, whereas the Black Sea's level fluctuated strongly (Lamy et al., 2006). Lake Oltina is situated at 45∘ N and thus at the transition between these contrasting southern and northern Europe changes in hydro-climatic conditions. Our evidence of the maximum expansion of the tree cover and possibly an association with wetter conditions after 4500 cal yr BP may indicate the response of tree cover to increased moisture availability in the region (Fig. 3). A greater-than-present extent of tree cover at 6000 cal yr BP in response to higher precipitation has also been simulated for this region (Patriche et al., 2020).
Macro-charcoal-based reconstruction of biomass burning and thus disturbance by fire was low between 4200 and 2500 cal yr BP at the time of greater forest cover and wetter climate conditions (Fig. 3). This fire–climate–vegetation relationship is typical for a temperate environment but contrasts with the pattern found in environments with low vegetation productivity, where increased moisture tends to enhance vegetation productivity and therefore fuel availability (Pausas and Ribeiro 2013; Feurdean et al., 2020). Disturbance by herbivores, as inferred from the abundance of coprophilous spores, showed moderate values at the beginning of the record but declined during the interval of tree cover increase (Fig. 3). This may point to some impact by herbivores on the degree of forest openness, i.e. increased tree cover with a decline in grazing activity. However, given the large size of the study lake, the distance from the lakeshore to the coring point might have limited the transportation of these spores and have an impact on how representative their presence might be (Baker et al., 2013).
5.2 Transition from forest steppe to cultural steppe over the last 2500 cal yr BP
Tree cover dropped from 50 % at 2500 cal yr BP to ∼ 20 % at 2200 cal yr BP (Fig. 5). Tree species composition retained a xerothermic character, although Carpinus orientalis, typical of hot and dry climatic and soil conditions (Sikkema and Caudullo, 2016), declined most strongly from 40 % to 10 % (Fig. 4). The pollen-based reduction in tree cover was concurrent with a decreasing n(C27 + C29) (C33 C31) ratio, characteristic of an increased contribution of grasses, further suggesting a tree loss (Fig. 4). An average of 20 % tree cover at 2200 cal yr BP is close to the current tree cover in the surroundings of Lake Oltina (15 %), which suggests the opening up of the forest steppe to a similar extent as today. Anthropogenic conversion of broadleaf forests to agricultural land from 2500 cal yr BP onwards is suggested by the rise in the pollen of Cerealia (Secale cereale, Triticum, Zea and Hordeum), grassland (Poaceae), and pastoral and ruderal indicators (P. lanceolata, Urtica, Rumex, Chenopodiaceae, Artemisia, Asteraceae and Apiaceae; Figs. 4, 5 and A2). The levels of pollen of grazing indicators and nutrient-enriched soils (Plantago lanceolata, Urtica and Rumex), as well as coprophilous fungi, are particularly elevated after 2200 cal yr BP and may reflect more intensive animal husbandry. A higher representation of Cerealia in the REVEALS estimate (40 %) than the raw pollen record (5 %) is not surprising given the poor productivity and dispersal of most of the Cerealia pollen types. However, the magnitude of this difference is greater than generally reported in the literature. Nevertheless, the good match between the proportion of arable cover in the REVEALS and current land cover maps from CORINE for recent times suggest that our REVEALS reconstruction best reflects that of cultivated land. Uncertainties in pollen-based land cover reconstruction are common and connected to the availability and accuracy of the productivity estimates (PPEs), changes in cropland and grassland management, and the general assumptions of the REVEALS model (Sugita, 2017; Hellman et al., 2009; Trondman et al., 2015; Feurdean et al., 2017b). Our Cerealia pollen includes Triticum, Zea, Hordeum and Secale cereale, for which we have used productivity estimates derived from the calibration of local surface pollen samples with a vegetation inventory (Grindean et al., 2019). These productivity estimates are considerably lower than the average for Europe (0.22 vs. 1.85; Mazier et al., 2012) and are therefore the main cause of the high proportion of Cerealia cover reconstructed by REVEALS and the disparity between the outcome of this study and others elsewhere in Europe. Furthermore, the crop species included in Cerealia also vary regionally and with time, which may also introduce further variation when applying their PPEs for landscape reconstruction over an extended period of time. Lastly, the occurrence of wild-grass species with pollen that fall in the Cerealia pollen type (all Poaceae grains larger than 40 µm) may have led to an overestimation of the proportion of Cerealia at certain times in the past. Biomass burning increased concurrently with the spread of pasture and grassland communities, which may indicate the use of fire for land use management (Fig. 3). However, fire activity fell to its lowest level in the profile in the last 1000 years, probably due to a decline in biomass availability associated with intensive land use and landscape fragmentation typical of lowlands (Marlon et al., 2016; Feurdean et al., 2013, 2020).
The timing of forest loss coincides with the peak number of archaeological finds in the Iron Age but is pre-dated by the abundant archaeological finds of the Bronze Age, i.e. 4000 cal yr BP (Fig. 5). Interestingly, the typology of the houses in this area (i.e. small houses three-quarters buried in the ground with one-quarter above ground comprising mud brick walls, roofed with straw or reeds typical during the past 3000 years) also reflects the limited availability of timber for building (Ailincăi, 2009). Historical records show that, due to its smaller size, Carpinus orientalis was managed in coppice stands for household items and fuel production as firewood or charcoal (Goldstein et al., 1995; San-Miguel-Ayanz et al., 2016), and this may have also been the cause of its marked reduction from 2500 cal yr BP at Lake Oltina. Quercus decline on the other hand was more modest except for the last 1000 years. Historically, the aftermaths of the Slavic (first millennium CE) and Mongol (1224 CE) invasions could be a reason for a strong Quercus decline around this time (Matei, 1984; Epure, 2004). Oak was preferentially used for fortification in southwestern Romania (Sava et al., 2019; Gumnior et al., 2020). Nevertheless, tree cover and Quercus, in particular, recorded episodic rises with the strongest being visible between 1700 and 1300 cal yr BP. This reforestation phase appears to coincide with the post-Roman decline in rural settlement and the subsequent re-growth of secondary forests (Roberts et al., 2018). However, selective oak preservation, due to its economic significance, may have also led to this pattern of Quercus increases, as documented over large areas in Europe (Gardner et al., 2002; Feurdean et al., 2011; Jamrichová et al., 2017; Gumnior et al., 2020). Models of deforestation rates, using a scenario that accounts for population history and technological advances, suggest that the extent of deforestation in the lower Danube basin increased continuously from 4000 cal yr BP and rapidly doubled after 1000 CE (Kaplan et al., 2009; Giosan et al., 2012), thus much later than our pollen-based land cover reconstruction. Assuming no age correction for the hard-water effect would result in an age 1000 years older than that calculated by taking into account this dating limitation, which pushes the timing of major deforestation back to 3500 cal yr BP. This corroborates with the timing of deforestation found in Transylvania, central Romania, based on the REVEALS model (Feurdean et al., 2015) and with the onset of a local increase in archaeological finds of the Bronze Age (Fig. 5).
In-lake and catchment changes are also apparent in the Lake Oltina around the onset of forest loss. A slight increase in the Fe : Mn ratio, along with the sharp rise in Zr concentrations between 2500 and 1500 cal yr BP, may reflect soil erosion and detrital input into the lake associated with a diminished tree cover (Fig. 3), an inference also supported by an abrupt rise in charcoal and n-alkane > C27 concentrations. Intensification in fluvial activity in several rivers in southern Romania (Howard et al., 2004; Persoiu and Radoane, 2017), as well as on a wider scale in central Europe (Wirth et al., 2013), has been reconstructed after 3000 cal yr BP. The Danube's water and sediment discharge could therefore be another source of detrital and terrestrial plant delivery into the lake. In contrast, the Black Sea level lowered, which also suggests drier conditions in the eastern Mediterranean and over the Black Sea around the time of deforestation (Lamy et al., 2006), in line with the drier conditions in southwestern Romania found between 2000 and 1000 cal yr BP (Dragusin et al., 2014). Southern European lake levels remained high at the time of deforestation, while those from central Europe (Magny et al., 2011, 2013) and central Romania declined (Feurdean et al., 2013), though others in Romania increased (Magyari et al., 2009).
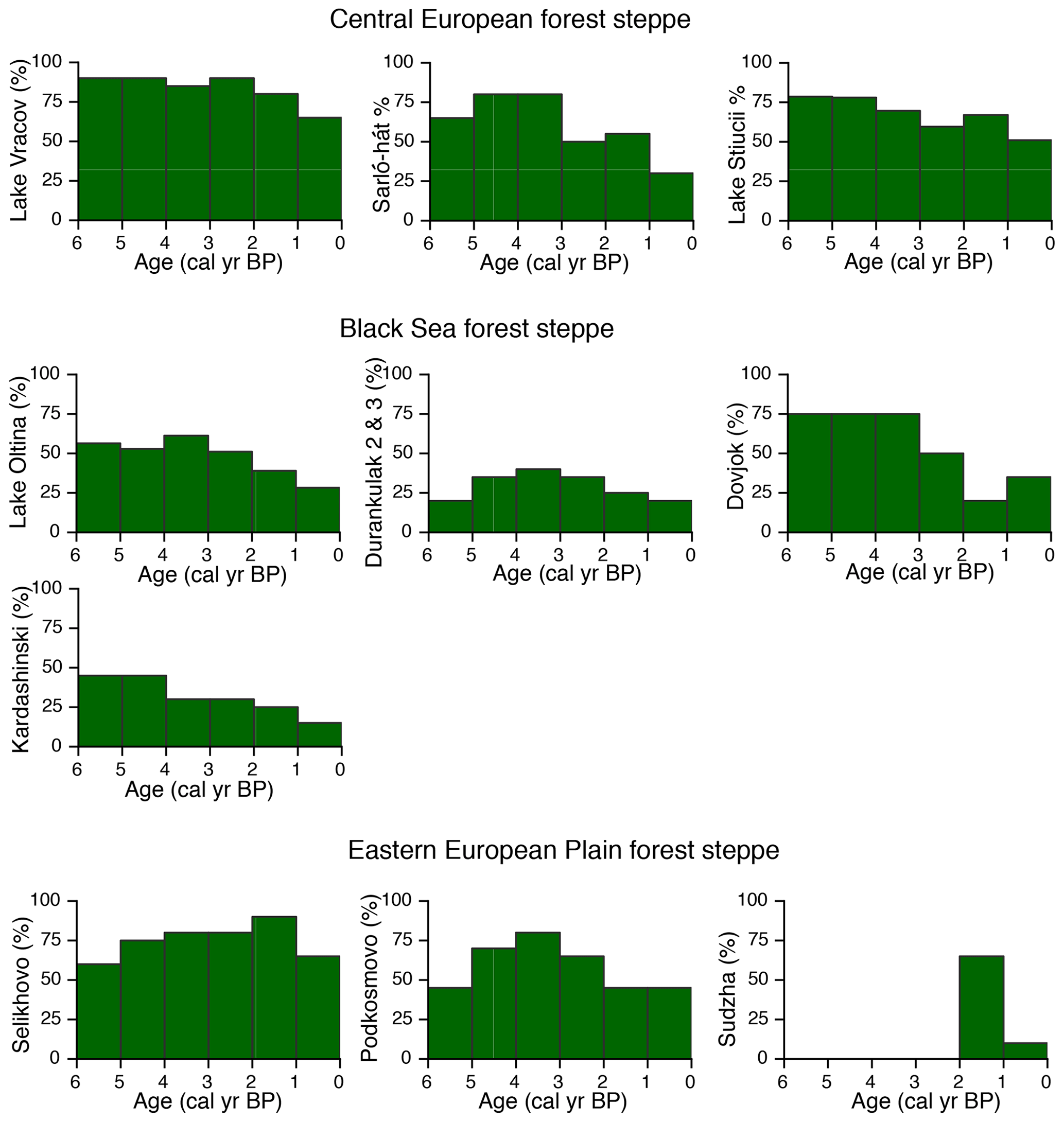
Figure 6Arboreal-pollen percentages illustrating temporal trends in deforestation in three different sub-regions along a west–east transect across the European forest steppe region. For the location of individual sites, see Fig. 1 and Table 3.
Table 3Compilation of palaeoecological records along a west–east transect of the European forest steppe. Cz: Czech Republic; Hu: Hungary; RO: Romania; BG: Bulgaria; UA: Ukraine; RU: Russia.
5.3 Comparison with other European forest steppe regions
Our quantitative record of vegetation cover indicates a higher-than-present tree cover across the landscape of the eastern lower Danube Plain between 6000 and 4200 cal yr BP with an absolute maximum of 50 % (60 % raw pollen percentages) between 4200 and 2500 cal yr BP (Fig. 6). The composition and structure of Mid to Late Holocene vegetation of the eastern lower Danube Plain resembled, to a large degree, that of other European forest steppe areas, although particularities also exist. Whilst in central-eastern European forest steppe Quercus and Carpinus betulus were the dominant tree species, with a lower occurrence of Tilia, Ulmus, Corylus and Pinus (Magyari et al., 2010; Feurdean et al., 2015; Kuneš et al., 2015), on the Eastern European Plain Tilia and Quercus and, in some places, Pinus were the dominant tree taxa (Kremenetski et al., 1995; Novenko et al., 2016, 2018; Shumilovskikh et al., 2018, 2019). Forests in the Black Sea region also included thermophilus taxa, i.e. Quercus cerris and C. orientalis (this study; Marinova and Atanassova, 2006; Tonkov et al., 2014). Remarkably, Carpinus orientalis was found to be significantly more abundant around the Black Sea coast, i.e. Romania (20 %) and Bulgaria (> 5 %) with only scattered occurrences in Ukraine, whilst it was absent from central European forest steppe. Our pollen–vegetation calibration model shows that the adjusted abundance of C. orientalis is twice that recorded by its raw pollen percentages (Fig. 4).
Mid Holocene landscape openness near Lake Oltina (ca. 45 % raw pollen percentages) appears to fall in between that of the steppe in southeastern Bulgaria (60 %–80 %; Tonkov et al., 2014), the forest steppe of Ukraine (25 %; Kremenetski et al., 1995, 1999) and that of the Eastern European Plain (20 %–50 %; Shumilovskikh et al., 2018). However, landscape openness was greater than in other forest steppe sites from central-eastern Europe, i.e. Romania (Feurdean et al., 2015; Tantau et al., 2006), Hungary (Willis et al., 1997; Magyari et al., 2010), the Czech Republic and Slovakia (Pokorny et al., 2015; Hajkova et al., 2013; Kuneš et al., 2015), where it varied between 10 % and 35 % (Fig. 6). The composition of herbaceous plant cover included grasses (Poaceae) and a diversity of forbs thriving on a wide variety of habitats ranging from dry and saline soils (Artemisia, Chenopodiaceae, Asteraceae Compositae and Asteraceae Tubuliflorae) to dry and wet meadows (Filipendula, Galium, Anthemis, Aster, Caryophyllaceae, Euphorbia, Helianthemum, Hypericum, Fabaceae, Plantago lanceolata, P. major/P. media, Teucrium, Thalictrum and Verbascum; Figs. 4; A2). Notably, however, the proportion of steppe and saline elements (Artemisia and Chenopodiaceae) was greater at sites located in the Black Sea region, in agreement with greater temperature seasonality, lower precipitation and the occurrence of saline soils in this region.
The comparison of pollen records from the European forest steppe shows a west-to-east gradient in the timing and magnitude of deforestation (Fig. 6). For example, the timing of major anthropogenic ecosystem transformation in the lower Danube Plain from about 2500 cal yr BP falls in between that of other records in lowland areas in central-eastern and southeastern Europe, where it generally occurred after 3000 cal yr BP (Fig. 6). However, this is earlier than on the Eastern European Plain, where it mostly occurred after 2000 cal yr BP. On the Thracian Plain, southeastern Bulgaria, anthropogenic deforestation was, however, noted already from 4000 cal yr BP (Connor et al., 2013). The anthropogenically driven opening up of the forest steppe soon reached a similar extent as today in most regions, which then remained open until the present day, although climate conditions could have allowed for the recovery of tree cover. Notably, however, the study region is increasingly confronted by desertification (European Environmental Agency, 2016). Given its dry character, the conversion of forests to cropland may have acted as a positive feedback to the warm and dry climate, enhancing evaporation and altering the moisture balance, further contributing to the tendency towards the aridization of Lower Danube landscapes. Ongoing climate change (warmer temperatures and a decline in precipitation), coupled with agricultural intensification, will probably exacerbate the process of desertification.
The pollen-based vegetation modelling applied here (REVEALS) provides the first, long-term quantitative reconstruction of land cover changes across the lower Danube Plain (southeastern Romania) and in southeastern Europe. Enhanced moisture availability likely led to a more extensive tree cover between 6000 and 2500 cal yr BP and its maximum of 50 % between 4200 and 2500 cal yr BP. This woodland consisted of tree taxa of xerothermic character including Quercus (likely Q. cerris and Q. pubescens) and Carpinus orientalis, and temperate trees such as Carpinus betulus, Acer, Tilia, Ulmus, Fraxinus. However, the proportion of xerophilous tree taxa, C. orientalis, increased between 4200 and 2500 cal yr BP. The forests of the lower Danube Plain were intensively cleared and converted to agricultural land and pasture and semi-natural grasslands from the Iron Age (2500 cal yr BP). The landscapes became deforested to present-day levels (ca. 20 % tree cover) 2200 years ago. Tree cover remained low throughout the last 2 millennia, demonstrating the continuous anthropogenic pressure on the region. The permanent loss of the tree cover is visible across a west–east gradient of the central-eastern European forest steppe region, highlighting its sensitivity to anthropogenic impact. Given the dry character of the study region, deforestation and land conversion to agriculture may have additionally enhanced evaporation, altering the moisture balance and further contributing to the tendency towards aridization. This palaeoecological study also demonstrates that, at a broad spatial scale, the natural vegetation of the eastern Romanian Plain under climatic conditions similar to today is forest steppe and woodlands, which is in agreement with expert-based assessments of potential tree cover. However, tree cover extent and composition have been neither stable in time nor solely shaped by the climate with disturbances by fire and grazing and later through anthropogenic impact, playing an important role. In comparison to pollen-based vegetation reconstruction, the PNV assumes a lower proportion of C. orientalis and a higher proportion of Quercus.
We also show that both the extension and decline in tree cover determined by pollen, a well-established proxy for past vegetation change, is also reflected in the n-alkane record, which indicates their potential as a reliable record of tree vs. grass cover changes in dry regions where reliable pollen records are difficult to obtain.
A1 Lake Oltina chronology
A chronology for Lake Oltina was established on the basis of 17 AMS 14C measurements (Table 1). Attempts to constrain the chronology of the top core via 210Pb and 137Cs gamma assay measurements failed to produce any meaningful results probably due to sediment mixing. An attempt via the geochemical identification of tephra also failed due to a poor match to any known volcanic eruption. The radiocarbon age estimates were converted into calendar years BP via the Bacon software (Blaauw and Christen, 2011) using the IntCal13 data set of Reimer et al. (2013). An age–depth curve was derived based on a smoothing spline model. Calendar age point estimates for depths were based on weighted average age–depth curves and also by taking into account the error range of the calibrated ages. Due to the lack of plant macrofossils, we used mostly shells and bulk material for the radiocarbon measurements. Consequently, age determination was problematic due to the low organic carbon levels and possible hard-water effects (Table A1).
We have attempted to correct for the reservoir effect in the following ways. Firstly, we compared our youngest radiocarbon date on the shell sample from 30 cm in depth (880 uncal yr BP) with the potential sediment age of recent samples based on geochemistry, mineral magnetic measurements and a specific pollen marker; 30 cm geochemical elements (particularly Pb) potentially associated with regional industrialization (after 1850) show concentrations above the background levels that would naturally be found, suggesting an additional anthropogenic input (Fig. A1). Further, mineral magnetic properties (X) also show an increase from 30 cm that might reflect an anthropogenic influence on the sediment (Fig. A1). Rose et al. (2009), Akinyemi et al. (2013) and Hutchinson et al. (2016) note atmospherically derived inputs of trace elements and heavy metals and mineral magnetic particles in the Romanian Carpathian Mountains from the start of the 20th century with peaks from the 1950s. Similarly, Begy et al. (2012) attribute peaks in heavy metals in a lake in the Danube delta to industrial and traffic pollution from the 1950s. We noted the occurrence of pollen of Ambrosia, an invasive species that arrived and spread after 1850, which also increased at this depth (Fig. A1). Taken together, results from the geochemistry, mineral magnetic measurements and pollen agree in suggesting that the age of the sediment at 30 cm must be after 1850 CE. Based on the difference of the two age estimates (880 years on the shell) and ∼ 50–100 years (via the geochemistry, mineral magnetic measurement and pollen), the age offset at 30 cm is about 700 years.
Secondly, for older sediments, we compared the radiocarbon date of the terrestrial macrofossil sample at a depth of 548 cm (3459 ± 3) with the shells from the same layer (4457 ± 35). Here the age difference between the two measurements is 1000 years, which is close to the 700 years age offset observed at 30 cm. Consequently, we have estimated the hard-water effect at about 800±200 years. In the Bacon model an age offset of 1000 years was specified for all radiocarbon dating on shells (Fig. 2). Furthermore, all measurements performed on bulk samples were rejected from the age–depth model. This is because the reservoir effect on bulk sediment is much larger than for shell, with a much larger possible error. In addition, in at least two age measurements on bulk samples (at 150 and 230 cm), the H fraction was very different from the L fraction, which indicates that the bulk organic matter is a composite organic material of a very different age. The results of our final model provide an age–depth curve with fewer age–depth reversals than seen when including the bulk samples.
Table A1AMS 14C measurements at Lake Oltina showing different age of the H and L fractions of bulk samples for two of the four samples performed using bulk material. HEKAL: HEKAL AMS Lab, Isotoptech Zrt.

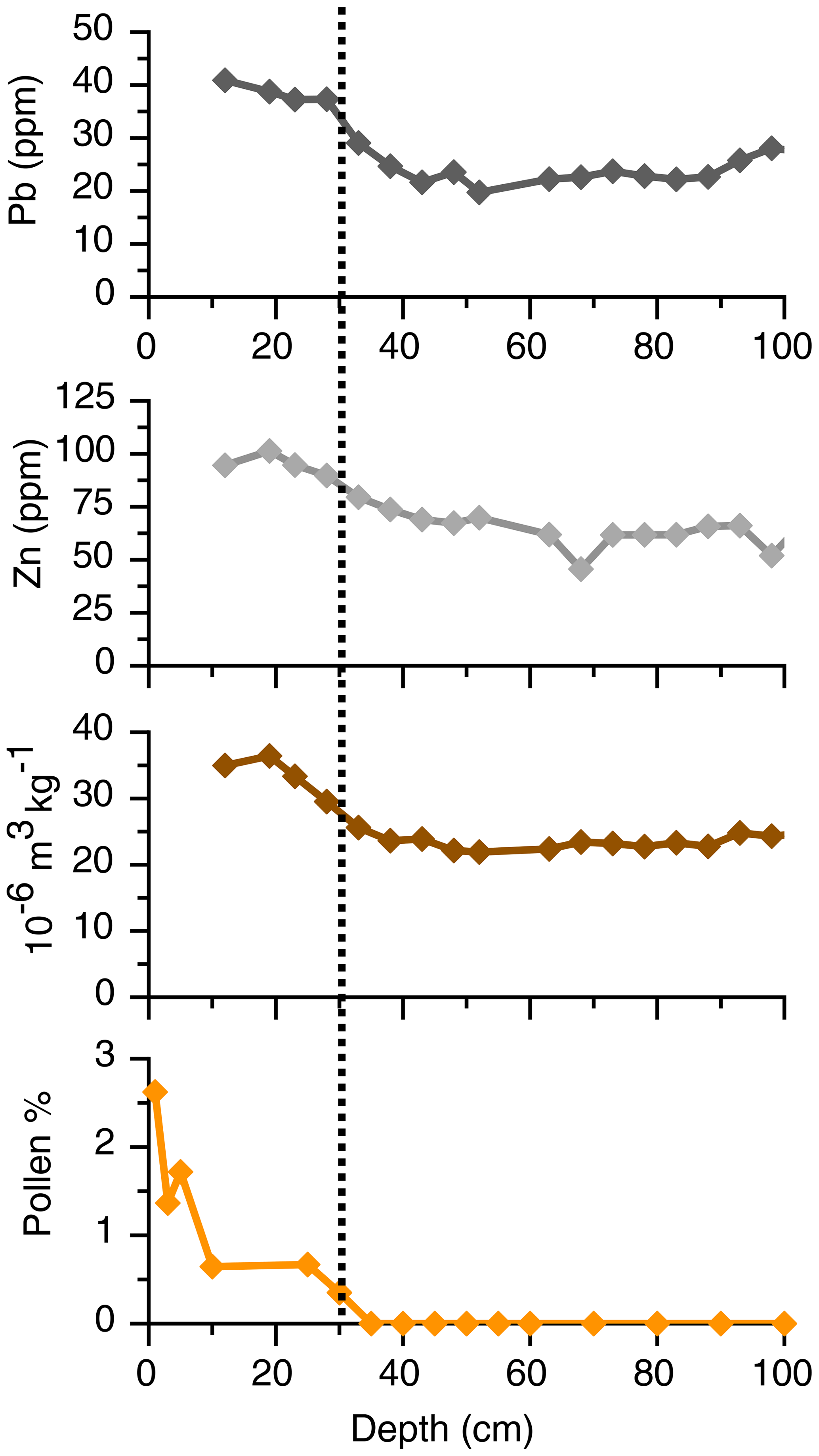
Figure A1Selected geochemical elements (Pb and Zn), mineral magnetic measurements (magnetic susceptibility) and pollen (Ambrosia), displaying their simultaneous increase after 30 cm (vertical dashed line) reflecting the post-1850 CE trend of regional industrialization and the known spread of an invasive weed.

Figure A2Full-pollen diagram for Lake Oltina grouped on trees, shrubs herbs, wetland and aquatic taxa as well as coprophilous fungi.
All essential input data can be requested from the corresponding author.
The supplement related to this article is available online at: https://doi.org/10.5194/bg-18-1081-2021-supplement.
AF designed the study. AF, AD, AP and MB performed the fieldwork. RG, IT and AF performed the pollen analysis. GF and EMN analysed the biomarkers. GF and SMH performed the geochemistry analysis. AD performed the macro-charcoal analysis. ST, AP and AF performed the C14 measurements and age–depth model results. RG, AN and AF performed the REVEALS modelling. AF prepared the paper with contributions and input from all authors.
The authors declare that they have no conflict of interest.
We thank the managers at Lake Oltina and Natura 2000 for granting and facilitating access to the lake for sediment sampling. Gabriela Florescu and Eva Niedermeyer thank Ulrich Treffert for laboratory support regarding biomarker analysis. Roxana Grindean thanks Sorina Farcas for granting laboratory access for pollen preparation. We thank Rebecca Kearney for her work on tephrostratigraphy, Mihaly Molnar for the suggestions on the construction of the age model and David, a student assistant, for assistance in the field. Finally, we thank the two reviewers, Simon Connor and Natalie Schroeter, for their constructive comments on the paper.
This research has been supported by the Deutsche Forschungsgemeinschaft (grant nos. FE_1096/4 and FE_1096/6) and the CNCS-UEFISCDI (grant nos. PN-II-RU-TE-2014-4-2445 and PN-III-P4-ID-PCE-2016-0711).
This open-access publication was funded
by the Goethe University Frankfurt.
This paper was edited by Sönke Zaehle and reviewed by Simon Connor and Natalie Schroeter.
Abraham, V. and Kozáková, R.: Relative pollen productivity estimates in the modern agricultural landscape of Central Bohemia (Czech Republic), Rev. Palaeobot. Palyno., 179, 1–12, https://doi.org/10.1016/j.revpalbo.2012.04.004, 2012.
Abraham, V., Oušková, V., and Kuneš, P.: Present-day vegetation helps quantifying past land cover in selected regions of the Czech Republic, Ploe One, 9, e100117, https://doi.org/10.1371/journal.pone.0100117, 2014.
Abraham, V., Kuneš, P., Petr, L., Svobodová, H. S., Kozakova, R., Jamrichova, E., Švarcová, M. G., and Pokorný, P.: A pollen-based quantitative reconstruction of the Holocene vegetation updates a perspective on the natural vegetation in the Czech Republic and Slovakia, Preslia, 88, 409–434, 2016.
Aichner, B., Herzschuh, U., Wilkes, H., Vieth, A., and Böhner, J.: δ D values of n-alkanes in Tibetan lake sediments and aquatic macrophytes – A surface sediment study and application to a 16 ka record from Lake Koucha, Org. Geochem., 41, 779–790, https://doi.org/10.1016/j.orggeochem.2010.05.010, 2010.
Ailincăi, S. C.: Începuturile epocii fierului în Dobrogea, PhD thesis, Faculty of History, Alexandru Ioan Cuza University, Romania, 2009 (in Romanian).
Akinyemi, F. O., Hutchinson, S. M., Mındrescu, M., and Rothwell, J. J.: Lake sediment records of atmospheric pollution in the Romanian Carpathians, Quat. Int., 293, 105–113, doi:10.1016/j.quaint.2012.01.022, 2013.
Baker, G. J., Bhagwat, S. A., and Willis, K. J.: Do dung fungal spores make a good proxy for past distribution of large herbivores? Quaternary Sci. Rev., 62, 21–31, https://doi.org/10.1016/j.quascirev.2012.11.018, 2013.
Bartington Instruments: Operation Manual for MS2 Magnetic Susceptibility System. Bartington Instruments Limited, Witney, Oxford, 90 p., available at: https://www.bartington.com/ms2-ms3-manuals/ (last access: 23 May 2018), 2008.
Begy, R. C., Kelemen, S., Simon, H., and Tănăselia, C.: The history of the sedimentation processes and heavy metal pollution in the Central Danube Delta (Romania), Geochronometria, 8, https://doi.org/10.1515/geochr-2015-0090, 2018.
Blaauw, M. and Christen, J. A.: Flexible paleoclimate age-depth models using an autoregressive gamma process, Bayesian Anal. 6, 457–474, https://projecteuclid.org/euclid.ba/1339616472, 2011.
Bozilova, E. and Tonkov, S.: Palaeoecological studies in Lake Durankulak, Ann. Univ. Sofia Fac. Biol., 2, 25–30, 1985.
Bozilova, E. and Tonkov, S.: Towards the vegetation and settlement history of the southern Dobrudza coastal region, North-eastern Bulgaria: a pollen diagram from Lake Durankulak, Veg. Hist. Archaeobot., 7, 141–148, 1998.
Bălăşescu, A. and Radu, V.: Omul şi animalele. Strategii şi resurse la comunităţile Hamangia şi Boian, Cetatea de Scaun, Targoviste, Romania, 2004 (in Romanian).
Broström, A., Sugita, S., and Gaillard, M. J.: Pollen productivity estimates for the reconstruction of past vegetation cover in the cultural landscape of southern Sweden, Holocene, 14, 368–381, https://doi.org/10.1191/0959683604hl713rp, 2004.
Bohn, U., Gollub, G., Hettwer, C., Neuhuslov., Z., Raus, Th., Schlüter, H., and Weber, H.: Karte der natürlichen Vegetation Europas/Map of the Natural Vegetation of Europe, Maßstab/Scale 1:2.500.000, Interaktive/Interactive CD-ROM-Erläuterungstext, Legende, Karten/Explanatory Text, Legend, Maps, Landwirtschaftsverlag, Münster, 2000/2003.
Chiarucci, A., Araújo, M. B., Decocq, G., Beierkuhnlein, C., and Fernández-Palacios, J. M.: The concept of potential natural vegetation: an epitaph?, J. Veg. Sci., 2, 1172–1178, https://doi.org/10.1111/j.1654-1103.2010.01218.x, 2010.
Cleary, D. M., Feurdean, A., Tan?ău, I., and Forray, F. L.: Pollen, δ15N and δ13C guano-derived record of late Holocene vegetation and climate in the southern Carpathians, Romania, Rev. Palaeobot. Palyno., 265, 62–75, https://doi.org/10.1016/j.revpalbo.2019.03.002, 2019.
Commerford, J. L., McLauchlan, K. K., and Sugita, S.: Calibrating vegetation cover and grassland pollen assemblages in the Flint Hills of Kansas, USA, Am. J. Plant Sci., 4, 1–10, https://doi.org/10.4236/ajps.2013.47A1001, 2013.
Connor, S. E., Ross, S. A., Sobotkova, A., Herries, A. I., Mooney, S. D., Longford, C., and Iliev, I.: Environmental conditions in the SE Balkans since the Last Glacial Maximum and their influence on the spread of agriculture into Europe, Quaternary Sci. Rev., 68, 200–215, https://doi.org/10.1016/j.quascirev.2013.02.011, 2013.
Dengler, J., Janišová, M., Török, P., and Wellstein, C.: Biodiversity of Palaearctic grasslands. A synthesis, Agr. Ecosyst. Environ., 182, 1–14, https://doi.org/10.1016/j.agee.2013.12.015, 2014.
Diefendorf, A. F., Leslie, A. B., Wing, S. L.: Leaf wax composition and carbon isotopes vary among major conifer groups, Geochim. Cosmochim. Ac., 170, 145–156, https://doi.org/10.1016/j.gca.2015.08.018., 2015.
Drăguşin, V., Staubwasser, M., Hoffmann, D. L., Ersek, V., Onac, B. P., and Veres, D.: Constraining Holocene hydrological changes in the Carpathian–Balkan region using speleothem δ18O and pollen-based temperature reconstructions, Clim. Past, 10, 1363–1380, https://doi.org/10.5194/cp-10-1363-2014, 2014.
Eglinton, G. and Calvin, M.: Chemical fossils, Scientific American, 216, 32–43, 1967.
Eglinton, T. I. and Eglinton, G.: Molecular proxies for paleoclimatology, Earth Planet. Sc. Let., 275, 1–16, https://doi.org/10.1016/j.epsl.2008.07.012, 2008.
European Environmental Agency: Mapping and assessing the condition of Europe's ecosystems: progress and challenges – 3rd report, 2016.
Epure, V. A.: Invazia mongolă în Ungaria si spatiul românesc. ROCSIR - Revista Româna de Studii Culturale, 13–26, 2004, last access: 28 May 2020.
Feurdean, A., Tanţău, I., and Fărcaş, S.: Temporal variability in the geographical range and abundance of Pinus, Picea abies, and Quercus in Romania, Quaternary Sci. Rev., 30, 3060–3075, https://doi.org/10.1016/j.quascirev.2011.07.005, 2011.
Feurdean, A., Liakka, J., Vanniere, B., Marinova, E., Hutchinson, S. M., Mossbruger V., and Hickler, T.: 12,000-Years of fire regime drivers in the lowlands of Transylvania (Central-Eastern Europe): a data-model approach, Quaternary Sci. Rev., 81, 48–61, https://doi.org/10.1016/j.quascirev.2013.09.014, 2013.
Feurdean, A., Perşoiu, A., Tanţău, I., Stevens, T., Magyari, E. K., Onac, B. P., Marković, S., Andrič, M., Connor, S., Fărcaş, S., Gałka, M., Gaudeny, T., Hoek, W., Kolaczek, P., Kuneš, P., Lamentowicz, M., Marinova, E., Michczyńska, D. J., Perşoiu, I., Płóciennik, M., Słowiński, M., Stancikaite, M., Sumegi, P., Svensson, A., Tămaş, T., Timar, A., Tonkov, S., Toth, M., Veski, S., Willis, K. J., and Zernitskaya, V.: Climate variability and associated vegetation response throughout Central and Eastern Europe (CEE) between 60 and 8 ka, Quaternary Sci. Rev., 106, 206–224, https://doi.org/10.1016/j.quascirev.2014.06.003, 2014.
Feurdean, A., Marinova, E., Nielsen, A. B., Liakka, J., Veres, D., Hutchinson, S. M., Braun, M., Timar-Gabor, A., Astalos, C., Mossbruger, V., and Hickler, T.: Origin of the forest steppe and exceptional grassland diversity in Transylvania (central-eastern Europe), J. Biogeogr., 42, 951–963, https://doi.org/10.1111/jbi.12468, 2015.
Feurdean, A., Veski, S., Florescu, G., Vannière, B., Pfeiffer, M., O'Hara, R. B., Stivrins, N., Amon, L., Heinsalu, A., Vassiljev, J., Hickler, T.: Broadleaf deciduous forest counterbalanced the direct effect of climate on Holocene fire regime in hemiboreal/boreal region (NE Europe). Quaternary Sci. Rev, 169, 378-390, https://doi.org/10.1016/j.quascirev.2017.05.024, 2017a.
Feurdean, A., Munteanu, C., Kuemmerle T., Nielsen, A. B., Hutchinson, S. M., Ruprecht, E., Parr, C. L., Persoiu, A., and Hickler, T.: Long-term land-cover/use change in a traditional farming landscape in Romania inferred from pollen data, historical maps and satellite images, Reg. Environ. Change, 17, 2193–2207, https://doi.org/10.1007/s10113-016-1063-7, 2017b.
Feurdean, A., Ruprecht, E., Molnár, Z., Hutchinson, S. M., and Hickler, T.: Biodiversity-rich European grasslands: Ancient, forgotten ecosystems, Biol. Conser., 228, 224–232, https://doi.org/10.1016/j.biocon.2018.09.022, 2018.
Feurdean, A., Vannière, B., Finsinger, W., Warren, D., Connor, S. C., Forrest, M., Liakka, J., Panait, A., Werner, C., Andrič, M., Bobek, P., Carter, V. A., Davis, B., Diaconu, A.-C., Dietze, E., Feeser, I., Florescu, G., Gałka, M., Giesecke, T., Jahns, S., Jamrichová, E., Kajukało, K., Kaplan, J., Karpińska-Kołaczek, M., Kołaczek, P., Kuneš, P., Kupriyanov, D., Lamentowicz, M., Lemmen, C., Magyari, E. K., Marcisz, K., Marinova, E., Niamir, A., Novenko, E., Obremska, M., Pędziszewska, A., Pfeiffer, M., Poska, A., Rösch, M., Słowiński, M., Stančikaitė, M., Szal, M., Święta-Musznicka, J., Tanţău, I., Theuerkauf, M., Tonkov, S., Valkó, O., Vassiljev, J., Veski, S., Vincze, I., Wacnik, A., Wiethold, J., and Hickler, T.: Fire hazard modulation by long-term dynamics in land cover and dominant forest type in eastern and central Europe, Biogeosciences, 17, 1213–1230, https://doi.org/10.5194/bg-17-1213-2020, 2020.
Ficken, K. J., Li, B., Swain, D. L., and Eglinton, G.: An n-alkane proxy for the sedimentary input of submerged/floating freshwater aquatic macrophytes, Org. Geochem., 31, 745–749, https://doi.org/10.1016/S0146-6380(00)00081-4, 2000.
Filipova-Marinova, M.: Archaeological and paleontological evidence of climate dynamics, sea-level change, and coastline migration in the Bulgarian sector of the Circum-Pontic region, in: The Black Sea flood question: Changes in Coastline, Climate and Human Settlement, edited by: Yanko-Hombach, V., Gilbert, A. S., Panin, N., and Dolukhanov, P. M., Springer, Dordrecht, Germany, 453–482, 2007.
Freymond, C. V., Peterse, F., Fischer, L. V., Filip, F., Giosan, L., Eglinton, T. I.: Branched GDGT signals in fluvial sediments of the Danube River basin: Method comparison and longitudinal evolution, Organic Geochemistry, 103, 88–96, https://doi.org/10.1016/j.orggeochem.2016.11.002, 2017.
Gardner, A. R.: Neolithic to Copper Age woodland impacts in northeast Hungary? Evidence from the pollen and sediment chemistry records, The Holocene, 12, 541–553, https://doi.org/10.1191/0959683602hl561rp, 2002.
Giosan, L., Coolen, M. J., Kaplan, J. O., Constantinescu, S., Filip, F., Filipova-Marinova, M., and Thom, N.: Early anthropogenic transformation of the Danube-Black Sea system, Sci Rep.-UK, 2, 582 https://doi.org/10.1038/srep00582, 2012.
Goeury, C. and de Beaulieu, J. L.: A propos de la concentration du pollen a l'aide de la liqueur de Thoulet dans les sediments mineraux, Pollen Spores, 21, 239–251, 1979.
Goldstein, M., Simonetti, G., Watschinger, M.: Alberi d'Europa – Guide pratiche Mondadori illustrati, Italy, 1995.
Grimm, E.: TGView 2.0.2., Springfield (IL): Illinois State Museum, Research and Collection Center, 2004.
Grindean, R., Nielsen, A. B., Feurdean, A., Tanţău, I.: Relative pollen productivity estimates in the forest steppe landscape of southeastern Romania, Rev. Palaeobot. Palyno., 264, 54–63, https://doi.org/10.1016/j.revpalbo.2019.02.007, 2019.
Gumnior, M., Herbig, C., Krause, R., Urdea, P., Ardelean, A. C., and Bălărie, A.: Palaeoecological evidence from buried topsoils and colluvial layers at the Bronze Age fortification Corneşti-Iarcuri, SW Romania: results from palynological, sedimentological, chronostratigraphical and plant macrofossil analyses, Veget Hist Archaeobot., 29, 173–188, https://doi.org/10.1007/s00334-019-00762-1, 2020.
Hájková, P., Jamrichová, E., Horsák, M., and Hájek, M.: Holocene history of a Cladium mariscus-dominated calcareous fen in Slovakia: vegetation stability and landscape development, Preslia, 85, 289–315, 2013.
Harrison, S. P., Gaillard, M.-J., Stocker, B. D., Vander Linden, M., Klein Goldewijk, K., Boles, O., Braconnot, P., Dawson, A., Fluet-Chouinard, E., Kaplan, J. O., Kastner, T., Pausata, F. S. R., Robinson, E., Whitehouse, N. J., Madella, M., and Morrison, K. D.: Development and testing scenarios for implementing land use and land cover changes during the Holocene in Earth system model experiments, Geosci. Model Dev., 13, 805–824, https://doi.org/10.5194/gmd-13-805-2020, 2020.
Hansen, S., Toderas, M., and Wunderlich, J.: Pietrele, Rumänien, Neolithische and Kuferzeritlische Siedlung am See 5200-4200 v. Chs. Eurasiean Abtailung des Deutschen Archäologischen Intituts, e-Forschungsberichte des Dai 2015 - Faszikel 3, 2015.
Heiri, O., Lotter, A. F., and Lemcke, G.: Loss-on-ignition as a method for estimating organic and carbonate content in sediments: reproducibility and comparability of results, J. Paleolimnol., 25, 101–110, https://doi.org/10.1023/A:1008119611481, 2001.
Hellman, S., Gaillard, M. J., Bunting, M. J., and Mazier, F.: Estimating the relevant source area of pollen in the past cultural landscapes of southern Sweden–a forward modelling approach, Rev. Palaeobot. Palynol., 153, 259–271, https://doi.org/10.1016/j.revpalbo.2008.08.008, 2009.
Howard, A. J., Macklin, M. V., Bailey, D. W., Mills, S., Andreescu, R.: Late Glacial and Holocene river development in the Teleorman Valley on the southern Romanian Plain, J. Quaternary Sci., 19, 271–280, https://doi.org/10.1016/j.quageo.2010.01.002, 2004.
Hutchinson, S. M., Akinyemi, F. O., Mîndrescu, M., Begy, R., and Feurdean, A.: Recent sediment accumulation rates in contrasting lakes in the Carpathians (Romania): impacts of shifts in socio-economic regime, Reg. Environ. Change, 16, 501–513. https://doi.org/10.1007/s10113-015-0764-7, 2016
IPBES 2019: Summary for policymakers of the global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services, edited by: Díaz, S., Settele, J., Brondízio, E.S., Ngo, H. T., Guèze, M., Agard, J., Arneth, A., Balvanera, P., Brauman, K. A., Butchart, S. H. M., Chan, K. M. A., Garibaldi, L. A., Ichii, K., Liu, J., Subramanian, A. M., Midgley, G. F., Miloslavich, P., Molnár, Z., Obura, D., Pfaff, A., Polasky, S., Purvis, A., Razzaque, J., Reyers, B., Roy Chowdhury, R., Shin, Y. J., Visseren-Hamakers, I. J., Willis, K. J., and Zayas, C. N., IPBES secretariat, Bonn, Germany, 2019.
IPCC, 2014: Summary for policymakers, in: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, edited by: Field, C. B., Barros, V. R., Dokken, D. J., Mach, K. J., Mastrandrea, M. D., Bilir, T. E., Chatterjee, M., Ebi, K. L., Estrada, Y. O., Genova, R. C., Girma, B., Kissel, E. S., Levy, A. N., MacCracken, S., Mastrandrea, P. R., and White, L. L., Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA.
Jamrichová, E., Petr, L., Jiménez-Alfaro, B., Jankovská, V., Dudová, L., Pokorný, P., Kołaczek, P., Zernitskaya, V., Čierniková, M., Břízová, E., and Syrovátka, V.: Pollen-inferred millennial changes in landscape patterns at a major biogeographical interface within Europe, J. Biogeogr., 44, 2386–2397, https://doi.org/10.1111/jbi.13038, 2017.
Jackson, S. T.: Natural, potential and actual vegetation in North America, J. Veg. Sci., 24, 772–776, https://doi.org/10.1111/jvs.12004, 2013.
Jamrichová, E., Bobek, P., Šolcová, A., Tkác, P., Hédl, R., and Valachovič, M.: Lowland pine forests in the northwestern Pannonian Basin: between natural vegetation and modern plantations, Reg. Environ. Change., 19, 2395–2409, https://doi.org/10.1007/s10113-019-01555-y, 2019.
Kaplan, J. O., Krumhardt, K. M., and Zimmermann, N.: The prehistoric and preindustrial deforestation of Europe, Quat. Sci. Rev., 28, 3016–3034, https://doi.org/10.1016/j.quascirev.2009.09.028, 2009.
Kremenetski, C. V., Chichagova, O. A., and Shishlina, N. A.: Palaeoecological evidence for Holocene vegetation, climate and land-use change in the low Don basin and Kalmuk area, southern Russia, Veget. Hist. Archaeobot., 8, 233–246, https://doi.org/10.1007/BF01291776, 1999.
Kremenetski, C. V.: Holocene vegetation and climate history of southwestern Ukraine, Rev. Palaeobot. Palyno., 85, 289–301, https://doi.org/10.1016/0034-6667(94)00123-2, 1995.
Kropf, M., Bardy, K., Höhn, M., and Plenk, K.: Phylogeographical structure and genetic diversity of Adonis vernalis L. (Ranunculaceae) across and beyond the Pannonian region, Flora, 262, https://doi.org/10.1016/j.flora.2019.151497, 2020.
Kuneš, P., Svobodová-Svitavská, H., Kolár, J., Hajnalová, M., Abraham, V., Macek, M., Tkác, P., and Szabó, P.: The origin of grasslands in the temperate forest zone of east-central Europe: long-term legacy of climate and human impact, Quaternary Sci. Rev., 116, 15–27, https://doi.org/10.1016/j.quascirev.2015.03.014, 2015.
Kylander, M. E., Ampel, L., Wohlfarth, B., and Veres, D.: High-resolution X-ray fluorescence core scanning analysis of Les Echets (France) sedimentary sequence: new insights from chemical proxies, J. Quatern. Sci., 26, 109–117, https://doi.org/10.1002/jqs.1438., 2011.
Lamy, F., Arz, H. W., Bond, G. C., Bahr, A., and Pätzold, J.: Multicentennial-scale hydrological changes in the Black Sea and northern Red Sea during the Holocene and the Arctic/North Atlantic Oscillation, Paleoceanography, 21, PA1008, https://doi.org/10.1029/2005PA001184, 2006.
Magny, M., Bossuet, G., Ruffaldi, P., Leroux, A., and Mouthon, J.: Orbital imprint on Holocene palaeohydrological variations in west- central Europe as reflected by lake-level changes at Cerin (Jura Mountains, eastern France), J. Quaternary Sci., 26, 171–177, https://doi.org/10.1002/jqs.1436, 2011.
Magny, M., Combourieu-Nebout, N., de Beaulieu, J. L., Bout-Roumazeilles, V., Colombaroli, D., Desprat, S., Francke, A., Joannin, S., Ortu, E., Peyron, O., Revel, M., Sadori, L., Siani, G., Sicre, M. A., Samartin, S., Simonneau, A., Tinner, W., Vannière, B., Wagner, B., Zanchetta, G., Anselmetti, F., Brugiapaglia, E., Chapron, E., Debret, M., Desmet, M., Didier, J., Essallami, L., Galop, D., Gilli, A., Haas, J. N., Kallel, N., Millet, L., Stock, A., Turon, J. L., and Wirth, S.: North–south palaeohydrological contrasts in the central Mediterranean during the Holocene: tentative synthesis and working hypotheses, Clim. Past, 9, 2043–2071, https://doi.org/10.5194/cp-9-2043-2013, 2013.
Magyari, E. K., Chapman, J. C., Passmore, D. G., Allen, J. R. M., Huntley, J. P., and Huntley, B.: Holocene persistence of wooded steppe in the Great Hungarian Plain, J. Biogeogr., 37, 915–35, https://doi.org/10.1111/j.1365-2699.2009.02261.x., 2010.
Magyari, E., Buczk0, K., Jakab, G., Braun, M., Pßl, Z., Karßtson, D., and Pap, I.: Palaeolimnology of the last crater lake in the Eastern Carpathian Mountains: a multiproxy study of Holocene hydrological changes, Hydrobiologia 631, 29–63, https://doi.org/10.1007/s10750-009-9801-1, 2009.
Marinova, E. and Atanassova, J.: Anthropogenic impact on vegetation and environment during the Bronze Age in the area of Lake Durankulak, NE Bulgaria: Pollen, microscopic charcoal, non-pollen palynomorphs and plant macrofossils, Rev. Palaeobot. Palynol., 141, 165–178, https://doi.org/10.1016/j.revpalbo.2006.03.011, 2006.
Marinova, E., Harrison, S. P., Bragg, F., Connor, S., De Laet, V., Leroy, S. A., Mudie, P., Atanassova, J., Bozilova, E., Caner, H., Cordova, C., Djamali, M., Filipova-Marinova, M., Gerasimenko, N., Jahns, S., Kouli, K., Kotthoff, U., Kvavadze, E., Lazarova, M., Novenko, E., Ramezani, E., Röpke, A., Shumilovskikh, L., Tanţǎu, I., and Tonkov, S.: Pollen-derived biomes in the Eastern Mediterranean–Black Sea–Caspian-Corridor, J. Biogeogr., 45, 484–499. https://doi.org/10.1111/jbi.13128, 2018.
Marlon, J. R., Kelly, R., Daniau, A.-L., Vannière, B., Power, M. J., Bartlein, P., Higuera, P., Blarquez, O., Brewer, S., Brücher, T., Feurdean, A., Romera, G. G., Iglesias, V., Maezumi, S. Y., Magi, B., Courtney Mustaphi, C. J., and Zhihai, T.: Reconstructions of biomass burning from sediment-charcoal records to improve data–model comparisons, Biogeosciences, 13, 3225–3244, https://doi.org/10.5194/bg-13-3225-2016, 2016.
Matei, H. C.: O istorie a lumii antice, Albatros, Bucuresti, 1984 (in Romanian).
Matthias, I., Nielsen, A. B., and Giesecke, T.: Evaluating the effect of flowering age and forest structure on pollen productivity estimates, Veg. Hist. Archaeobotany, 21, 471–484, doi:10.1007/s00334-012-0373-z, 2012.
Mapping and assessing the condition of Europe's ecosystems: progress and challenges – 3rd report, European Environmental Agency, 2016.
Mazier, F., Gaillard, M. J., Kuneš, P., Sugita, S., Trondman, A. K., and Broström, A.: Testing the effect of site selection and parameter setting on REVEALS-model estimates of plant abundance using the Czech Quaternary Palynological Database, Rev. Palaeobot. Palyno., 187, 38–49, https://doi.org/10.1016/j.revpalbo.2012.07.017, 2012.
Meyers, P. A.: Applications of organic geochemistry topaleolimnological reconstructions: a summary of examples from the Laurentian Great Lakes, Org. Geochem., 34, 261–289, https://doi.org/10.1016/S0146-6380(02)00168-7, 2003.
Naeher, S., Gilli, A., North, R. P., Hamann, Y., and Schubert, C. J.: Tracing bottom water oxygenation with sedimentary Mn/Fe ratios in Lake Zurich, Switzerland, Chem. Geol., 352, 125–133, https://doi.org/10.1016/j.chemgeo.2013.06.006, 2013.
Novenko, E. Y., Eremeeva, A. P., and Chepurnaya, A. A.: Re-construction of Holocene vegetation, tree cover dynamics and human disturbances in central European Russia, using pollen and satellite data sets, Veg. Hist. Archaeobot., 23, 109–119, https://doi.org/10.1007/s00334-013-0418-y, 2014.
Novenko, E., Tsyganov, A., Payne, R., Mazei, N., Volkova, E., Chernyshov, V., Kupriyanov, D., and Mazei, Y.: Vegetation dynamics and fire history at the southern boundary of the forest vegetation zone in European Russia during the middle and late Holocene, Holocene, 28, 308–322, https://doi.org/10.1177/0959683617721331, 2018.
Novenko, E. Y., Tsyganov, A. N., Rudenko, O. V., Volkova, E. V., Zuyganova, I. S., Babeshko, K. V., Olchev, A. V., Losbenev, N. I., Payne, R. J., and Mazei, Y. A.: Mid- and late-Holocene vegetation history, climate and human impact in the forest-steppe ecotone of European Russia: new data and a regional synthesis, Biodivers. Conserv, 25, 2453–2472, https://doi.org/10.1007/s10531-016-1051-8, 2016.
Nowacki, D., Langan, C. C. M., Kadereit, A., Pint, A., and Wunderlich, J.: “Lake Gorgana”-A paleolake in the Lower Danube Valley revealed using multi-proxy and regionalisation approaches, Quaternary Int., 511, 107–123, https://doi.org/10.1016/j.quaint.2018.09.021, 2019.
Pausas, J. G. and Ribeiro, E.: The global fire–productivity relationship, Global Ecol. Biogeogr., 22, 728–36, https://doi.org/10.1111/geb.12043, 2013.
Patriche, C. V., Pîrnău, R. G., and Roşca, B.: Reconstructing the spatial distribution of large vegetation formations during the mid-Holocene in Romania: a predictive modeling approach. J. Quaternary Sci, 35, 1081–1088, https://doi.org/10.1002/jqs.3243.2020, 2020.
Perșoiu, A., Onac, B. P., Wynn, J. G., Blaauw, M., Ionita, M., and Hansson, M.: Holocene winter climate variability in central and eastern Europe, Sci. Rep.-UK, 7, 1196, https://doi.org/10.1038/s41598-017-01397-w, 2017.
Perșoiu, I. and Radoane, M.: Fluvial Activity During the Holocene, in: Landform Dynamics and Evolution in Romania, edited by: Radoane M. and Vespremeanu-Stroe A., Springer Geography, Springer, Cham, https://doi.org/10.1007/978-3-319-32589-7_20, 2017.
Planul de Management pentru ariile naturale protejate: ROSCI0172 Pădurea şi Valea Canaraua Fetii-Iortmac [Management Plan for Protected areas: ROSCI0172 Forest and Valley Canaraua Fetii-Iortmac], available at: http://www.mmediu.ro/app/webroot/uploads/files/2016-05-12_PM_si_R__Canaraua_Fetii_Iortmac.pdf, (last accesse: 28 May 2019), 2016 (in Romanian).
Plenk, K., Willner, W., Demina, O., Höhn, M., Kuzemko, A., Vassilev, K., and Kropf, M.: Phylogeographic evidence for long-term persistence of the Eurasian steppe plant Astragalus onobrychis in the Pannonian region (eastern Central Europe), Flora, 264, 151555, https://doi.org/10.1016/j.flora.2020.151555, 2020.
Pokorný, P., Chytrý, M., Juřičková, L., Sádlo, J., Novák, J., and Ložek, V.: Mid-Holocene bottleneck for central European dry grasslands: Did steppe survive the forest optimum in northern Bohemia, Czech Republic?, Holocene, 25, 716–726, https://doi.org/10.1177/0959683614566218, 2015.
Posea, G.: Geomorfologia Romaniei: relief, tipuri, geneza, evolutie, regionare. Editura fundatiei' Romania de Maine' Bucuresti, 444 pp., 2005.
Preoteasa, L., Vespremeanu-Stroe, A., Panaiotu, C., Rotaru, S., Ţuţuianu, L., Sava, T., Bîrzescu,, I., Dimofte, D., Sava, G., Mirea, D.A., and Ailincăi, S.: Neolithic to modern period paleogeographic transformations in southern Danube Delta and their impact on human settlements from Enisala–Babadag region, Quatern. Int., 504, 139–152, https://doi.org/10.1016/j.quaint.2018.09.010, 2018.
Poynter, J. and Eglinton, G.: Molecular composition of three sediments from hole 717c: The Bengal fan, in: Proceedings of the Ocean Drilling Program: Scientific results, 116, 155–161, 1990.
Reille, M.: Pollen et Spores d'Europe et d'Afrique du Nord. Supplément 1. Laboratoire de Botanique Historique et Palynologie, Marseille, France, 1995 (in French).
Reille, M.: Pollen et Spores d'Europe et d'Afrique du Nord. Supplément 2. Laboratoire de Botanique Historique et Palynologie, Marseille, France, 1999 (in French).
Reimer, P. J., Bard, E., Bayliss, A., Beck, J. W., Blackwell, P. G., Bronk Ramsey, C., Buck, C. E., Cheng, H., Edwards, R. L., Friedrich, M., Grootes, P. M., Guilderson, T. P., Haflidason, H., Hajdas, I., Hatté, C., Heaton, T. J., Hoffmann, D. L., Hogg, A. G., Hughen, K. A., Kaiser, K. F., Kromer, B., Manning, S. W., Niu, M., Reimer, R. W., Richards, D. A., Scott, E. M., Southon, J. R., Staff, R. A., Turney, C. S. M., and van der Plicht, J.: IntCal13 and Marine13 radiocarbon age calibration curves 0-50,000 years cal BP, Radiocarbon, 55, 1869–1887, https://doi.org/10.2458/azu_js_rc.55.16947, 2013.
Roberts, N., Fyfe, R. M., Woodbridge, J., Gaillard, M. J., Davis, B.A., Kaplan, J. O., Marquer, L., Mazier, F., Nielsen, A. B., Sugita, S., and Trondman, A. K.: Europe's lost forests: a pollen-based synthesis for the last 11,000 years, Sci. Rep.-UK, 158, 716, https://doi.org/10.1038/s41598-017-18646-7, 2018.
Romanescu, G. T., Dinu, C. I., Radu, A. L., and Török, L. I.: Ecologic characterization of the fluviatile limans in the South-West Dobrudja and their economic implications (Romania), Carpath. J. Earth Env., 5, 25–38, 2010.
Rose, N. L., Cogalniceanu, D., Appleby, P. G., Bancelj, A., Camarero, L., and Fernandez, P.: Atmospheric contamination and ecological changes inferred from the sediment record of Lacul Negru in the Retezat National Park, Advances in Limnology, 62, 319e350, 2009.
Rull, V.: Long-term vegetation stability and the concept of potential natural vegetation in the Neotropics, J. Veg. Sci., 26, 603–607, https://doi.org/10.1111/jvs.12278. 2015.
Sachse, D., Billault, I., Bowen, G. J., Chikaraishi, Y., Dawson, T. E., Feakins, S. J., Freeman, K. H., Magill, C. R., McInerney, F. A., van der Meer, M. T. J., Polissar, P., Robins, R. J., Sachs, J. P., Schmidt, H. L., Sessions, A. L., White, J. W. C., West, J. B., and Kahmen, A.: Molecular paleohydrology: interpreting the hydrogenisotopic composition of lipid biomarkers from photosynthesizing organisms, Annu. Rev. Earth. Pl. Sc., 40, 221–249, https://doi.org/10.1146/annurev-earth-042711-105535, 2012.
San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., and Mauri, A.: European atlas of forest tree species, Publications Office of the European Union, Luxembourg, 2016.
Sârbu, A., Negrean, G., and Sârbu, I.: The grasslands of the Dobrogea, Romania, in: Grasslands in Europe of High Nature Value, edited by: Veen, P., Jefferson, R., Smidt, J., and van der Straaten, KNNV Publishing, The Netherlands, 218–225, 2009.
Sava, V., Gogaltan, F., and Krause, R.: First Steps in the Dating of the Bronze Age Mega-Fort in Sântana-CetateaVeche (Southwestern Romania), in: Bronze Age Fortresses in Europe. Proceedings of the Second International LOEWE Conference, 9–13 October 2017 in Alba Julia (Bonn), 2019.
Sikkema, R. and Caudullo, G.: Carpinus orientalis in Europe: distribution, habitat, usage and threats, in: European Atlas of Forest Tree Species, edited by: San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., and Mauri, A., Publication Office of the European Union, Luxembourg, 2016.
Shumilovskikh, L. S., Novenko, E., and Giesecke, T.: Long-term dynamics of East-European forest-steppe ecotone, J. Veg. Sci., 29, 416–426, https://doi.org/10.1111/jvs.12585, 2018.
Shumilovskikh, L. S., Rodinkova, V., Rodionova, A., Troshina, A., Ershova, E., Novenko, E., Zazovskaya, E., Sycheva, S. A., Kiselev, D., Schlütz, F., and Schneeweiß, J.: Insights in the late Holocene vegetation history of the East European forest-steppe: case study Sudzha (Kursk region, Russia), Veg. Hist. Archaeobot, 28, 513–528, https://doi.org/10.1007/s00334-018-00711-4, 2019.
Sugita, S.: Theory of quantitative reconstruction of vegetation I: pollen from large sites REVEALS regional vegetation composition, Holocene, 17, 229–241, https://doi.org/10.1177/0959683607075837, 2007.
Tantau, I., Reille, M., de Beaulieu, J.-L., and Farcas, S.: Late Glacial and Holocene vegetation history in the southern part of Transylvania (Romania): pollen analysis of two sequences from Avrig, J. Quaternary Sci., 21, 49–61, https://doi.org/10.1002/jqs.937, 2006.
Telteu, C. E.: Phytoplankton Diversity in the Oltina And Bugeac Lakes and their Dependency on the Climatic Conditions and Physico–Chemical Parameters, Lakes Reservoirs Ponds, 8, 82–95, 2014.
Tomescu, A.: Evaluation of Holocene pollen records from the Romanian Plain, Rev. Palaeobot. Palyno., 109, 219–233, 2000.
Tonkov, S., Marinova, E., Filipova-Marinova, M., Bozilova, E.: Holocene palaeoecology and human environ- mental interactions at the coastal Black Sea Lake Durankulak, northeastern Bulgaria, Quatern. Int., 10, 277–286, https://doi.org/10.1016/j.quaint.2013.12.004, 2014.
Trondman, A. K., Gaillard, M. J., Mazier, F., Sugita, S., Fyfe, R., Nielsen, A. B., Twiddle, C., Barratt, P., Birks, H. J. B., Bjune, A. E., Björkman, L., Broström, A., Caseldine, C., David, R., Dodson, J., Dörfler, W., Fischer, E., van Geel, B., Giesecke, T., Hultberg, T., Kalnina, L., Kangur, M., van der Knaap, P., Koff T., Kuneš, P., Lagerås, P,. Latałowa, M., Lechterbeck, J., Leroyer, C., Leydet, M., Lindbladh, M., Marquer, L., Mitchell, F. J. G., Odgaard, B. V., Peglar, S. M., Persson, T., Poska, A., Rösch, M., Seppä, H., Veski, S., and Wick, L.: Pollen-based quantitative reconstructions of Holocene regional vegetation cover (plant functional types and land-cover types) in Europe suitable for climate modelling, Glob. Chang. Biol., 21, 676–697, https://doi.org/10.1111/gcb.12737, 2015.
van Geel, B., Bohncke, S. J. P., and Dee, H.: A palaeoecological study of an upper Late Glacial and Holocene sequence from “De Borchert”, The Netherlands, Rev. Palaeobot. Palynol., 31, 367–448, https://doi.org/10.1016/0034-6667(80)90035-4, 1980/81.
Vasiliev, I., Reichart, G. J., and Krugsman, W.: Impact of the Messinian Salinity Crisis on Black Sea hydrology – insights from hydrogen isotopes analysis on biomarkers, Earth Planet. Sc. Lett., 362, 272–282, https://doi.org/10.1016/j.epsl.2012.11.038, 2013.
Vincze, I., Finsinger, W., Jakab, G., Braun, M., Hubay, K., Veres, D., Deli, T., Szalai, Z., Szabó, Z., Magyari, E.: Paleoclimate reconstruction and mire development in the Eastern Great Hungarian Plain for the last 20,000 years, Rev. Palaeobot. Palynol., 271, 104–112, https://doi.org/10.1016/j.revpalbo.2019.104112, 2019.
Weninger B., Clare L., Rohling E., Bar-Yosef O., Böhner U., Budja M., Bundschuh M., Feurdean A., Gebe H. G., Jöris O., Linstädter J., Mayewski P., Mühlenbruch T., Reingruber A., Rollefson G., Schyle D., ThissenL., Todorova H., and Zielhofer C.: The impact of rapid climate change on prehistoric societies during the Holocene in the Eastern Mediterranean, Doc. Praehist., 36, 7–59. https://doi.org/10.4312/dp.36.2, 2009.
Walter, H.: Vegetationsmonographien der einzelnen Großräume, Band VII, Die Vegetation Osteuropas, Nord- und Zentralasiens, Gustav Fischer Verlag, Stuttgart, 1974 (in German).
Wunderlich, J., Herbig, C., Marinova, E., Nowacki, D., and Röpke, A.: Landschafts- und Flussgeschichte der unteren Donau im Umfeld des Siedlungshügels Magura Gorgana, Südrumänien: In: Verzweigungen. Eine Würigung für AJ Kalis und J Meurers-Balke, Fraunkfurter Archäologische Schriften, 18, 333–350, Bonn, 2012 (in German).
Willis, K. J., Braun, M., Sumegi, P., and Toth, A.: Does soil change cause vegetation change or vice versa? A temporal perspective from Hungary, Ecology, 78, 740–750, https://doi.org/10.1890/0012-9658, 1997.
Willis, K. J. and Birks, H. J. B.: What is natural? The need for a long-term perspective in biodiversity conservation, Science, 314, 1261–1265, https://doi.org/10.1126/science.1122667, 2006.
Wirth, S. B., Glur, L., Gilli, A., and Anselmetti, F. S.: Holocene flood frequency across theCentral Alps e solar forcing and evidence for variations in North Atlantic atmospheric circulation, Quat. Sci. Rev., 80, 112–128, https://doi.org/10.1016/j.quascirev.2013.09.002, 2013.
Whittaker, R. J., Willis, K. J. and Field, R.: Scale and species richness: towards a general, hierarchical theory of species diversity, J. Biogeogr., 28, 453–470, https://doi.org/10.1046/j.1365-2699.2001.00563.x. 2001.
Zhou, W. J., Xie, S. C., Meyers, P. A., and Zheng, Y. H.: Reconstruction of late glacial and Holocene climate evolution in southern China from geolipids and pollen in the Dingnan peat sequence, Org. Geochem., 36, 1272–1284, https://doi.org/10.1016/j.orggeochem.2005.04.005, 2005.