the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 24 Mar 2022
| 24 Mar 2022
Geodiversity and biodiversity on a volcanic island: the role of scattered phonolites for plant diversity and performance
David Kienle
Anna Walentowitz
Leyla Sungur
Alessandro Chiarucci
Severin D. H. Irl
Anke Jentsch
Ole R. Vetaas
Richard Field
Carl Beierkuhnlein
Oceanic islands are cradles of endemism, contributing substantially to global biodiversity. A similarity in magmatic origin translates into high global comparability of substrates of volcanic islands on the oceanic crust with, however, slightly chemically or physically differentiated petrography in some places. Phonolites are examples of rare localities with intermediate chemical characteristics between felsic and mafic and with diverse textures. They contribute to habitat heterogeneity and offer specific growth conditions in a significantly different matrix of basaltic substrates. The explicit contribution of geodiversity to island biodiversity has been little studied, despite growing evidence of its importance on continents. On the island of La Palma, Canary Islands, isolated phonolitic rocks are conspicuous due to their light colour and specific shape. Although these outcrops only cover small areas, their unique form and composition increase within-island geodiversity. To investigate how this affects biodiversity on La Palma, we sampled all vascular plant species in 120 plots on four sets of paired sites in order to test if plant diversity and performance is enhanced on phonolitic rocks compared to basaltic rocks. We recorded species number and abundance as well as individual plant height and diameter as proxies for aboveground resource allocation and tested for differences in vegetation cover and species composition between the bedrock types. We found higher species richness and abundance on phonolites than neighbouring basaltic substrates, and individuals of the same species were larger (in height and diameter) on phonolites compared to neighbouring basalt. An endemic woody species with two distinct varieties even appears almost exclusively on the small surfaces of phonolitic rock. Despite extremely limited spatial extent, phonolitic rocks can play an important role in plant biodiversity on islands.
- Article
(2948 KB) - Full-text XML
- BibTeX
- EndNote





Biodiversity is known to depend mainly on abiotic drivers, such as climate and topography (Field et al., 2009). However, the importance and explicit impacts of geodiversity on biodiversity have long been insufficiently researched and partly ignored. Only recently has the topic started to receive more attention (e.g. Gray, 2004; Lawler et al., 2015; Bailey et al., 2017; Alahuhta et al., 2020; Barajas-Barbosa et al., 2020). Geodiversity is in many respects an abiotic equivalent to biodiversity (Gray, 2011) and represents the variability of chemical components, surface structure, and edaphic and hydrological features (Gray, 2004; Bailey et al., 2017). This variability contributes to habitat diversity and thus affects biodiversity patterns via the provisioning of ecological niches (Liu et al., 2013; Gillespie and Roderick, 2014; Bailey et al., 2017). Geological elements provide unique or distinctive habitats for plants and insects, deliver initial growth conditions for vegetation or fungi formation, and are part of nutrient cycling and soil–atmosphere interactions (Tukiainen et al., 2016).
Biodiversity is distributed unevenly throughout the world (Gaston, 2000), with a disproportionately large contribution from oceanic islands due to their high endemic richness (Kier et al., 2009). Substrates that differ in geochemistry and petrography are likely to be relevant for biodiversity on oceanic islands, where most rocks commonly share similar volcanic genesis, resulting in only slight differences in the parent material. Distinct substrates with limited extent, such as individual rock types, may function as a second isolating abiotic filter for populations in addition to the spatial isolation of oceanic islands that are known to be of outstanding importance for speciation at the global scale (Kier et al., 2009). Specific rock habitats, particularly rocks that exhibit petrographic and geochemical substrates such as serpentinites, are known to be rich in habitat-specific endemics (e.g. Harrison et al., 2006; Kazakou et al., 2010). Those species evolved specific adaptations to the unique nutrient contents and soil conditions and the presence of heavy metals that cannot be tolerated by other plant species (Harrison and Rajakaruna, 2011). This phenomenon is known as well on continents, where substrates such as serpentinite and gypsum outcrops host specialised floras and contribute to broad-scale diversity (see, e.g. Chiarucci et al., 1998; Pausas et al., 2003). It underlines the relevance of understanding the importance of geodiversity for insular biodiversity, which is particularly vulnerable to extinction due to highly restricted ranges and small population sizes of insular endemic species (Paulay, 1994). Phonolites are rocks that occur at volcanic intraplate settings in insular and continental contexts worldwide (Garcia et al., 1986; Ackerman et al., 2015; Hagos et al., 2017). They exist in a variety of geologic outcrops formed by volcanic activity. Such outcrops mainly exist on continents, where they are often linked to faults and tectonic activity. Major components of these extrusive igneous rocks (formed from lava with low silica content) are alkaline feldspars together with foid minerals, nepheline, and pyroxene (Abratis et al., 2015; Ackerman et al., 2015) or their conversion products.
On the island of La Palma (Canary Islands, Spain), several phonolitic rock outcrops are embedded into a matrix of basaltic origin (Middlemost, 1972). The dominant rock type found on La Palma is olivine and augite–titanaugite porphyric basalt, resulting from rapidly rising magma from the upper mantle (Middlemost, 1970). In contrast, there were times when a sizeable magmatic chamber below the island enabled differentiation of magma and the removal of silica, thus yielding ultramafic, trachytic, and phonolitic rocks (Middlemost, 1970). Phonolite trachytes (showing the exhalation of gases during eruptions) occur on various volcanic islands such as La Palma, St Helena, Fernando de Noronha, Trinidade and Sal (Cabo Verde). On La Palma, their distribution is focused on the southern (young) part of the island. The major chemistry of phonolites on La Palma is comparable to that of “average phonolites”, as described by Nockolds (1954). The current volcanic activity and lava flow deriving from the Cumbre Vieja volcano (Pankhurst et al., 2021) are a demonstration of how phonolitic rocks became isolated by younger lava solidifying around the peaks of phonolitic rock. This event illustrates that the isolation of the investigated phonolitic habitats (e.g. Roque Teneguía) is far from being a singularity but rather a process that is highly likely to have happened repeatedly on oceanic islands in general.

Figure 1The endemic Cheirolophus junonianus (a, bottom left) and further plant species on a phonolite rock (© Severin Irl). Aerial image of rocks of phonolites isolated in a basaltic matrix in southern La Palma (b, © Google Earth 2020).
Volcanic activity with production of tephra and lava flows is a noticeably young phenomenon in the southern part of La Palma, with even present-day eruptions (Pankhurst et al., 2021). Thus, the remnant phonolitic rocks are the tips of a former land surface that are today embedded in a basaltic matrix of noticeably immature age (Garantje et al., 1998). As a consequence, weathering processes on phonolites were active on longer timescales compared to other surrounding rocks. In addition to petrography, differences in weathering between the rock types and resulting nutrient availability also infer different timescales of exposure.
A higher nutrient availability enables higher plant abundances and larger plant size. Porder et al. (2004) found comparable conditions at a catena of different rock ages on the Hawaiian Islands. Compared to basaltic lava outcrops, phonolites differ in their chemical composition and additionally in colour, texture, density, weathering, and formation fracturing (von Fragstein et al., 1988). Tafoni-weathering (Formoso et al., 1989) can be observed on phonolitic surfaces, indicating temperature and moisture gradients between the surface and the solid body of rocks (Brandmeier et al., 2011) that appear in combination with wind exposure. Circulating leachate reaches the rock's surface and evaporates, exposing its dissolved mineral content and enabling the development of secondary mineral assemblages (Spürgin et al., 2019). These can contribute to plant nutrient supply, which is also why ground phonolite rock powder is used as an effective fertiliser (Faccini et al., 2015). For phonolites, increased release of nutrients can be mediated by bio-weathering actions and plants receiving this fertiliser showed higher productivity, and increased accumulation of the macronutrient potassium in plants could be detected when applying phonolite rock powder (Tavares et al., 2018; Nogueira et al., 2021). Phonolites and the related larger-grained nepheline syenites contain significantly larger amounts of the essential nutrient potassium (see Table A1 in the Appendix for a literature overview). Even if quantitatively small, such processes are of particular importance at nutrient-poor sites. In contrast, the young basalts in the southern part of La Palma are barely weathered (Carracedo et al., 1999), appearing rough and friable with sharp spikes. We expect these petrographic and geochemical differences of parent material to affect vegetation cover and species occurrence.
Geologic outcrops, such as phonolites, increase microenvironmental heterogeneity, enhancing species richness at a landscape scale (Hjort et al., 2015). Increased speciation rates on isolated outcrops of scarce rocks are thought to lead to a higher percentage of endemic species than the surrounding matrix (Ricketts, 2001). Geodiversity may thus promote both species richness and endemism. However, relatively little is known about the extent to which phonolites promote species diversity in general and particularly endemism. To approach this topic theoretically, phonolitic outcrops could be considered as small habitat islands within a basaltic matrix (Fig. 1b). The established species–area relationship (SAR) and the species–isolation relationship (SIR; MacArthur and Wilson, 1967; Rosenzweig, 1995; Giladi et al., 2014) predict a smaller number of species on these small and isolated phonolitic rocks in comparison with basaltic rock outcrops in their surroundings. From the beginning of these concepts (MacArthur and Wilson, 1967), they were not only meant for real islands but instead took “islands” as examples of isolated habitats (or habitat islands) within a terrestrial landscape matrix. However, the expected higher availability of nutrients would give such habitats more favourable conditions for plant growth. In addition, it is by no means certain that the phonolitic rocks were permanently separated from each other in southern La Palma's geological evolution. Possibly, a historically much larger phonolitic rock is today largely buried by basaltic eruptions (Garantje et al., 1998). Thus, a few phonolite outcrops may serve as refugia for remnant populations (Eriksson, 1996) of species specialised to phonolitic rocks.
La Palma hosts 159 vascular plant species that are endemic to the archipelago and 47 single-island endemics (hereafter SIEs; Beierkuhnlein et al., 2021). The endemic plant species Cheirolophus junonianus (Svent.) Holub, comprising its var. junonianus and var. isoplexiphyllus (Svent.) G. Kunkel (Vitales et al., 2014a, b, Beierkuhnlein et al., 2021), occurs within a range of only 3500 m2, solely on La Palma (Bañares et al., 2004). Within this small range, individuals of this species occur only on a few outcrops, which are almost exclusively phonolitic rocks with a chemical composition different from most of the surrounding substrates. Therefore, the species is very restricted in its range size to just a few small locations (Muer et al., 2016; Atlantis, 2021) and appears to be restricted to phonolites (Fig. 1a). This example species evokes the question of whether or not phonolites are of special importance for endemic species on the Canary Islands.
We aim to investigate plant species richness, abundance and performance on phonolites compared to surrounding basaltic lavas. Therefore, we investigated the occurrences and traits of plant species in a comparative study matching basaltic and phonolitic rock formations on La Palma of comparable size, shape, and extent to test the following hypotheses:
- i.
Species diversity: we expect plant species richness on phonolitic rocks to be higher than on basaltic rocks because phonolites offer more favourable plant growth conditions.
- ii.
Plant performance: we expect plant species populations on phonolites show a larger abundance of individuals that are taller and have greater canopy diameter than neighbouring basalts due to their advantages in resource availability and porosity. We used plant performance as a surrogate for plant fitness.
- iii.
Island endemism: we expect phonolitic rocks to host more endemic plant species than basaltic rocks because of their high degree of spatial isolation, in combination with the older age of the phonolitic bedrock than the basaltic matrix.
2.1 Study site and data sampling
We sampled four phonolitic and four adjacent basaltic rocks in the southern part of La Palma in spring 2018 (Walentowitz et al., 2021; Fig. 2). Locations were identified in the field based on Middlemost (1972). The sampled phonolitic rocks represented most of the overall extent of this habitat on the island, covering a large gradient of microclimate, aspect (“northernness” and “easternness”), and inclination. Local climate data are not available for individual plots, nor for the sites. Interpolated modelled climate data (Karger et al., 2017) show only small variations in temperature and precipitation values for our study sites (Table A2). We chose comparable neighbouring pairs of phonolite and basalt consisting of one cohesive rock formation each. Outcrop pairs were chosen to match the size and microclimatic conditions (aspect, slope). For each selected phonolitic and basaltic rock, we recorded plant species composition and abiotic parameters within 15 plots of 2 m × 2 m that were randomly selected within the range of accessibility on phonolite and basalt. This resulted in a total of 120 plots sampled across the four pairs of phonolitic and basaltic rocks (60 plots on phonolite and 60 plots on basalt).
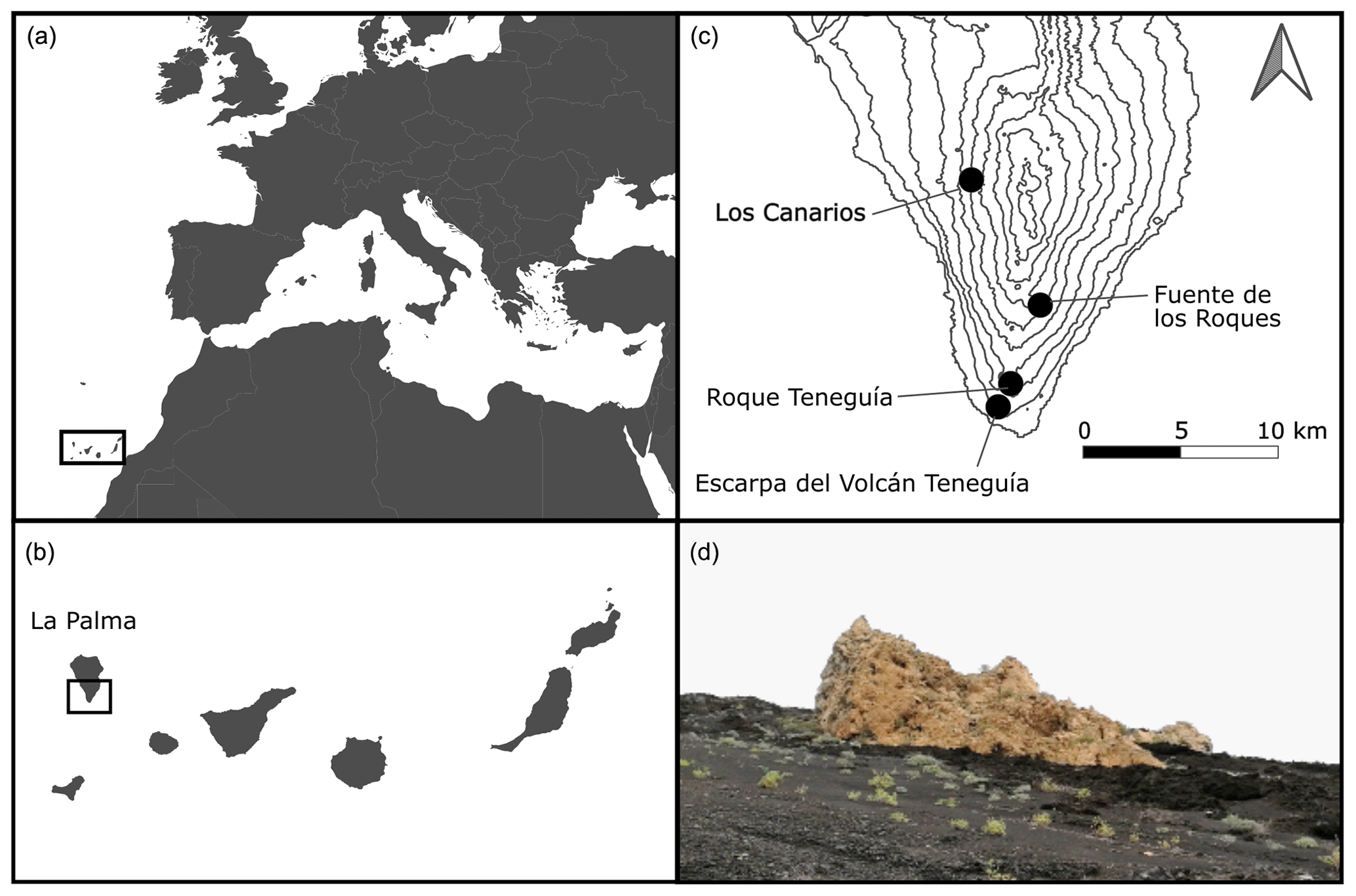
Figure 2Location of the Canary Islands (a) and La Palma (b). Southern La Palma with our four study sites and contour lines at 200 m intervals (c). Phonolite rock on southern La Palma downwards slopes of the “Roque Teneguía”, located in a basaltic matrix (d, © Anna Walentowitz).
Within each plot, we recorded coordinates, aspect, and slope inclination. Then, we estimated rock surface rugosity using thread transects spanning the two plot diagonals: we measured the transect length along the 3D rock surface (Walentowitz et al., 2021). Larger values of rugosity indicate higher levels of microtopography (cracks, hollows, uneven slope), while low values indicate smooth, even surfaces.
All vascular plant species within each plot, including ferns, were identified following the taxonomy of Beierkuhnlein et al. (2021) that use Plants of the World Online (POWO, 2019) as a taxonomic backbone. The biogeographic status of each species (SIE; multi-island endemic, MIE; non-endemic native; and introduced) is based on Muer et al. (2016; see extensive plant list in Appendix A3). The number of individuals per species and plot was counted.
Plant height (length from base of the stem to the tip) and canopy diameter (widest part of the plant parallel to the ground) of all single individuals found were measured as traits. Height, diameter, and species abundances were measured for all vascular plant species. As plant communities were dominated by perennial species, we can expect that vegetational differences evolved through long-term processes and did not reflect the short-term variability of environmental conditions. We are aware that there is a serious debate on the trade-off between different functional traits and their effect on plant growth responses. However, we assume that height and diameter are good proxies for different components such as survival and reproduction that contribute to plant fitness (Laughlin et al., 2020). We furthermore know that numbers of flowers and seeds might be more accurate to measure and monitor over the course of an entire reproductive cycle, but we chose plant height and width as proxies as these can be measured at the same time.
Lichen cover, which is abundant on the basalt, was estimated as the percent cover of each plot. Moss cover was negligible in all the plots.
2.2 Statistical analysis
Differences in total plant species number and the number of SIEs and MIEs were analysed using Pearson's Chi-squared tests. Percentages of abundance, plant height, diameter, and SIE percentage between plots on phonolites and basalt were analysed using Mann–Whitney U tests. We conducted detrended correspondence analysis (DCA) to investigate the multidimensional aspects of vegetation composition and identify potential fundamental underlying drivers (Fig. A1). Afterwards, we applied a post hoc permutation test (10 000 repetitions) between the environmental variables (substrate, inclination, aspect, and relief) and the DCA ordination axes (Table A3). We tested for differences in aspect, inclination, rock surface rugosity, and lichen cover between phonolite and basalt using Mann–Whitney U tests.
We recorded 68 species of vascular plants (pteridophytes and spermatophytes) overall. Of these species, nine were SIEs restricted to the island of La Palma, 16 were MIEs co-occurring on other islands in the archipelago, 39 were non-endemic natives, and 4 were introduced. The SIE Cheirolophus junonianus was only found on phonolite, and most individuals of var. junonianus occurred on one isolated outcrop (Roque Teneguía) and individuals of var. isoplexiphyllus on another one (Escarpa del Volcán Teneguía, Fig. 2c).
We found higher plant species richness on phonolitic rocks. While 22 species were encountered on both phonolite and basalt, only 11 species were restricted to basalt, and 34 were recorded only on phonolite (Table A4). Endemism groupings showed similar patterns (SIEs – phonolite: 9, basalt: 5; MIEs – phonolite: 15, basalt: 6). Besides the total number of plant species per rock type, we also found higher species richness on phonolite at the plot scale (p=0.0164, Fig. 3a), and higher diversity of SIEs (p=0.00151, Fig. 3b) and MIEs (p=0.00727, Fig. 3d). The percentage of SIEs (p=0.1928, Fig. 3c) and MIEs (p=0.05346, Fig. 3e) relative to total species number did not differ significantly at this scale between phonolitic and basaltic rocks.
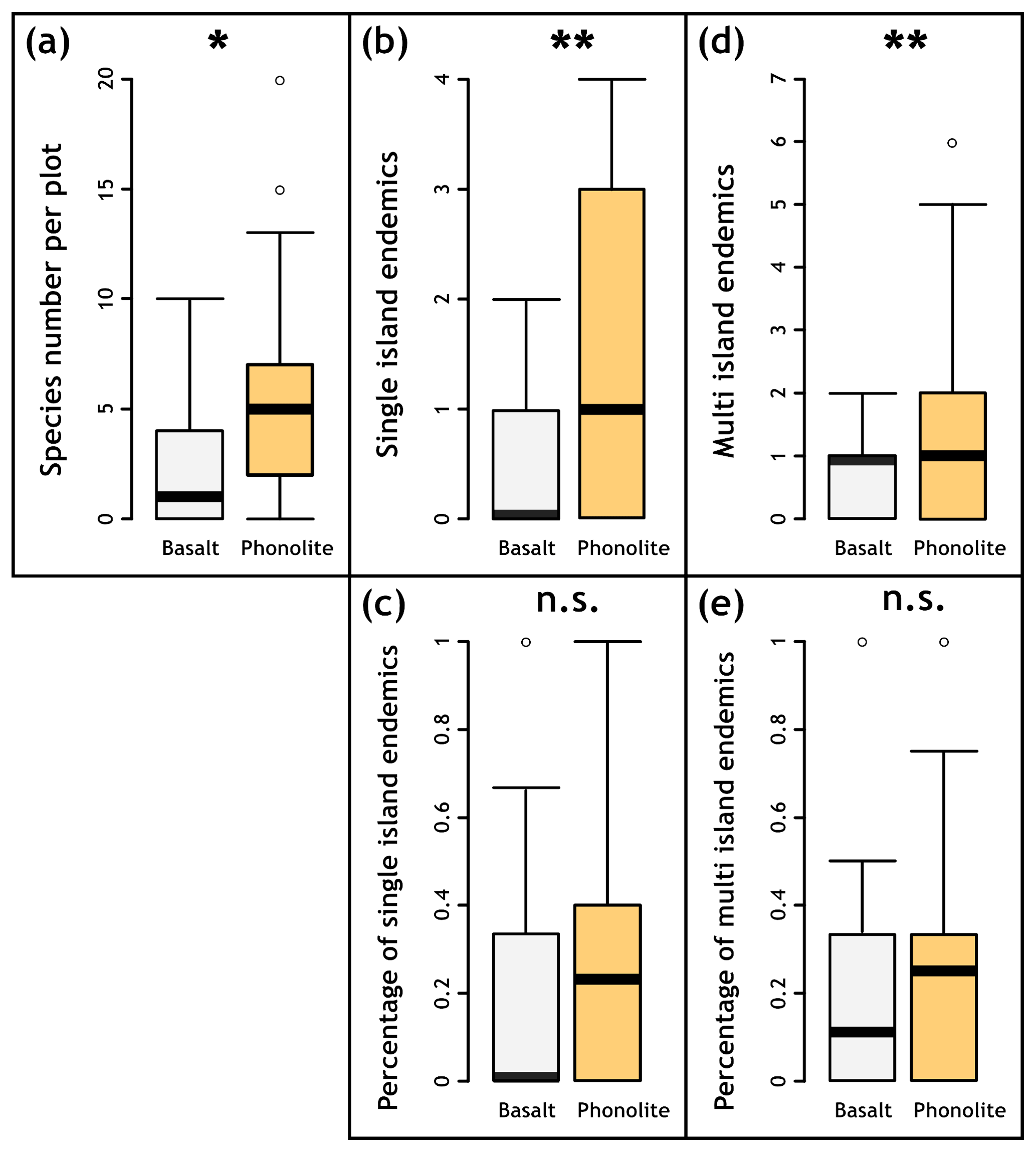
Figure 3Perennial species per 2 × 2 m plot for basaltic (n=60) and phonolite substrates (n=60). (a) The number of species per phonolite plot is significantly larger than for basaltic plots. (b) Phonolites have significantly more SIEs and (d) MIEs. However, the numbers of endemic species relative to the total number of species do not differ significantly between substrates (c, e). All analyses were conducted with Pearson's Chi-squared test (a, b, c) and the Mann–Whitney U test (c, e).
On phonolitic rocks, we did not find higher total plant abundance (p=0.169, Fig. 4a). Moreover, there was no significant difference in abundance when only considering the 23 species found in plots on both substrates (p=0.179, Fig. 4b).
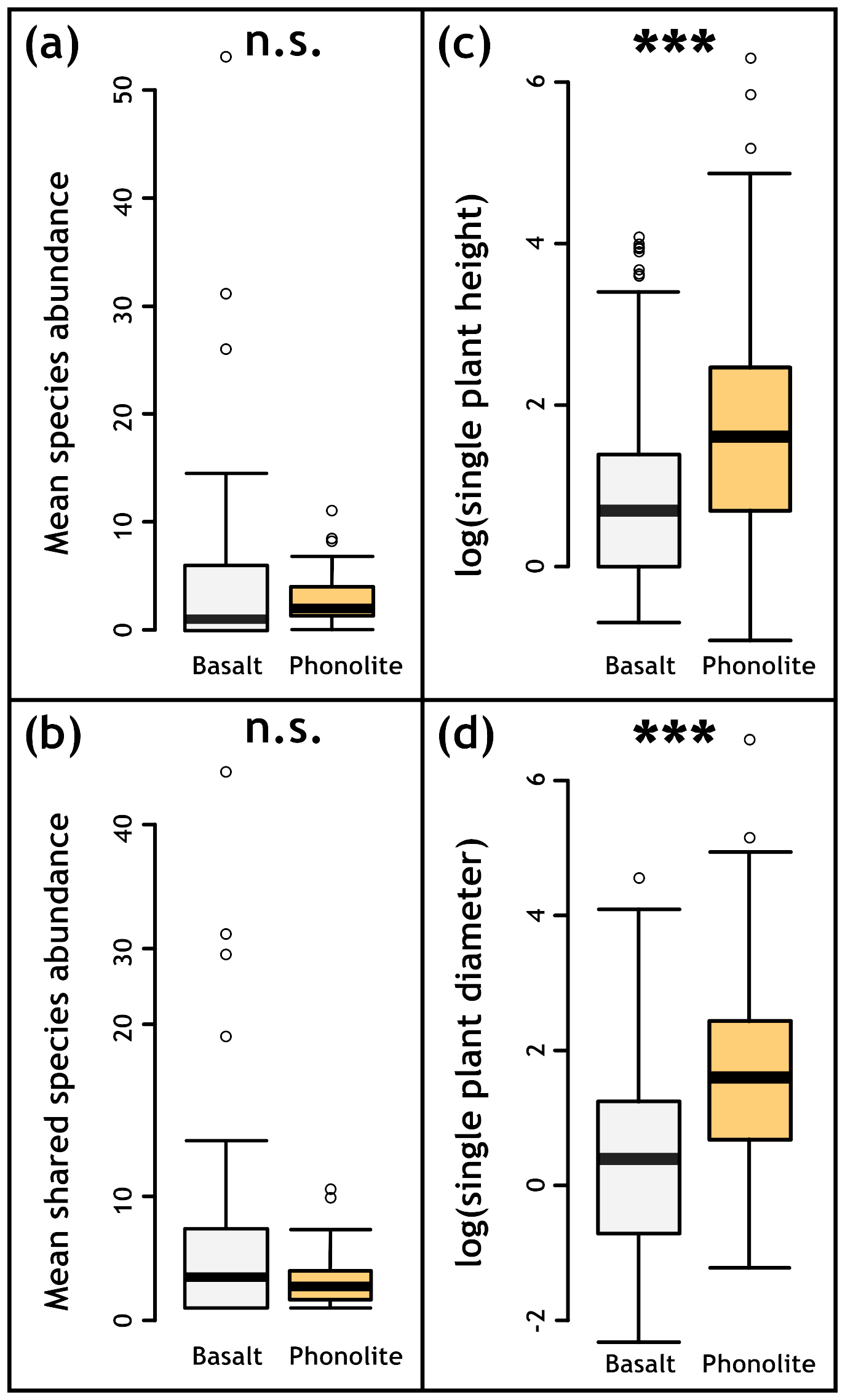
Figure 4(a) Mean abundance differs significantly between basaltic (n=60) and phonolite substrates (n=60), but (b) considering only shared species on both substrates resulted in no significant difference between basalt (n=39) and phonolite (n=51). (c, d) plant height and diameter (both log10-transformed) are significantly larger on phonolite plots (n=1560) than on basaltic plots (n=1173). Plot size: 2×2 m. All analyses were conducted with the Mann–Whitney U test.
For plant species recorded on both rock types, individuals were on average taller and had wider canopies (Fig. 4c–d) on phonolitic than on basaltic rocks (Fig. 4c–d). Plant cover only (excluding lichens) was also significantly higher (p<0.0001, Fig. A2a) on phonolites than on basalt. Lichen cover did not show a significant difference (p=0548, Fig. A2b).
The ordination did not show any difference in the species composition, indicating no aspects of beta-diversity at all (Fig. A1, Table A3). Topographic characteristics of basalt and phonolite plots showed no differences in surface rugosity, aspect (northernness, easternness), or differences in slope inclination (Table A5).
The vegetation on phonolitic rocks differs compared to equivalent neighbouring basaltic rocks by exhibiting higher species richness and higher performance of plant individuals and higher total plant cover. Larger numbers of SIEs and MIEs on phonolites also reflect augmented total species numbers on this rock type.
Phonolite rocks on La Palma and other oceanic islands are arranged as habitat islands in a basaltic matrix (Fig. 1b). On La Palma, the total area of phonolite outcrops is tiny compared to the area of the basaltic matrix. Thus, encountering more species on phonolites than on basalt aligns with our hypotheses but defies the area effect on species richness, which predicts species number to be lower on phonolites. Differences in species numbers might be attributed to lower-than-expected species numbers growing on basalt, higher-than-expected species numbers on phonolites, or both. Our findings are congruent with studies that did not find a species–area relationship or relationships with a less steep slope on habitat islands (Matthews et al., 2016; Deák et al., 2018). The existence of unrealised niches due to unsaturated evolutionary dynamics in a young system could explain lower-than expected species numbers on basalt. A possible extinction debt that might lead to the disappearance of species in the future might explain higher-than expected species numbers on phonolitic outcrops. Only the continuous monitoring of populations on both rock types can help to verify these assumptions. Environmental filters (Liu et al., 2020) enhancing growth conditions on phonolite outcrops may also exist, consistent with our findings that plants are larger on phonolites.
The rock types phonolite and basalt differ in their chemical composition resulting in different nutrient availability, which explains our observation of increased plant performance on phonolites. Phonolites consist of the potassium-rich nepheline, which dissolves much faster than other potassium sources (Manning, 2010). Various studies indicate that phonolites and the related nepheline syenite contain a higher proportion of potassium than basalt (Manning, 2010; Roqueto do Reis, 2021). Therefore, ground phonolite rock powder has traditionally been used as an inorganic fertiliser (von Wilpert and Lukes, 1998; Ramos et al., 2006; Schoen et al., 2016). Basaltic rock powder has also been used as fertiliser but is considered a less important source of potassium than phonolite (Manning, 2010). The usage of ground basalt as fertiliser can also be explained by unclear assignments (potassium-rich trachyte is often assigned to basalt; see Maning, 2010). In addition to geochemical differences, the duration of rock weathering is a decisive factor in providing nutrients. The phonolitic outcrops in southern La Palma are substantially older than the surrounding basalt, which stems from very young volcanic eruptions (Carracedo et al., 1999). The youngest nearby eruption of the Teneguía volcano took place only 50 years ago, in 1971.
The age of geological formations influences plant diversity and species compositions (Whittaker et al., 2008; Hulshof and Spasojevic, 2020). As noted in previous studies (Carracedo et al., 1999), the Cumbre Vieja rift on La Palma has evolved throughout several eruptions and therefore contains lava formations from different ages as well as slightly different mineralogical compositions. The current volcanic activity and lava flow at the Cumbre Vieja are a live example of this geological process (Pankhurst et al., 2021). The known phonolite rocks on La Palma are located in the geologically young southern part of the island. As a consequence, the species pool in the surrounding basaltic matrix of these rocks is poor. Under the arid conditions of southern La Palma, only very few early successional species establish on these young basaltic outcrops with not more than initial soil formation (Irl et al., 2019). The few rocky outcrops of phonolite are embedded in this species-poor matrix of young basalt. We observed partly buried phonolites on which the survival of plants or seedlings during volcanic events was improbable (Garantje et al., 1998). Carracedo et al. (1999) showed that the last phonolite formation occurred in 1585, while basaltic eruptions continue until modern times (Pankhurst et al., 2021). 19 plant species, including Cheirolophus junonianus, can solely be encountered on phonolitic rocks (Irl et al., 2015; Muer et al., 2016). This confirms that habitat diversity on islands contributes to their total species richness (Hortal et al., 2009).
Besides petro-chemical characteristics and rock age, the surface structure and colour of phonolites might be suspected to drive plant patterns on such rocks. We observed deeper fractures in phonolitic rocks than in other volcanites on La Palma. In addition, phonolitic rocks show a much smoother surface roughness than their surrounding matrix. Basaltic rocks seem to possess a more dynamic relief, mainly attributed to their origin in congealed lava flows, typically found on oceanic islands. Nevertheless, when testing rock surface rugosity, there were no significant differences between phonolitic and basaltic rocks. Hence, we argue that surface characteristics do not play a role in higher plant growth response, richness, and abundance observed on phonolites. Besides fractures, another visual observation was that phonolites are of lighter colour than their surrounding basaltic matrix. We expected that phonolites possess a higher albedo than surrounding rocks and therefore expected them to have a reduced surface temperature compared to volcanic outcrops with darker colouring, such as basalt. However, in an experiment with differently coloured bricks, Hall et al. (2005) showed that the albedo of white surfaces only leads to significantly lower temperature of the material when the surface temperature falls below air temperature. With monthly temperatures between 17 and 25 ∘C within large areas of oceanic islands (Harter et al., 2015), no major temperature differences between basalt and phonolite surfaces can be expected. We therefore consider that this effect has no major impact on plants' habitat suitability.
While a diversity of rocks with different chemical characteristics and at different ages supports species richness on volcanic islands, such rock characteristics do not necessarily contribute to higher percentages of endemic plants or distinct compositional vegetation on individual rocky outcrops. The overall percentage of SIEs on phonolites was not significantly enhanced, refuting our expectations. Thus, the differential geology of phonolites itself does not result in a specialised flora. Obviously, the small outcrops of phonolite on La Palma do not suffice to evolve and maintain a substantial set of endemic species, which contrasts with general assumptions that patterns caused by differing topography or discontinuous parent material can be explained by island biogeographic theory (e.g. Kruckerberg 1991). Consequently, lessons learnt from other outcrops (Kruckerberg, 1991) cannot be adapted to the phonolitic rocks on La Palma, and the functioning of phonolites as islands of speciation within a matrix of basalt does not seem to apply.
Despite the limited spatial extent of phonolites on La Palma, they contribute to insular habitat heterogeneity, which translates into increased species richness and abundance as well as higher plant performance. These phenomena are facilitated by specific characteristics of phonolite rock, like high nutrient availability fortified by longer geological timeframes for erosion and nutrient release compared to basalt. We are not aware of other studies conducted in locations where phonolites can be encountered that explore their potential role as exceptional plant habitat islands, even though phonolites can be found all over the world (Garcia et al., 1986; Ackerman et al., 2015; Hagos et al., 2017). Therefore, further studies are needed to investigate whether the patterns encountered on La Palma may also be found on comparable phonolitic rocks in other areas of the world. Their benefits for biodiversity found in this study need to be recognised and valued. Especially for isolated areas such as islands, phonolites can contribute to small-scale biodiversity hotspots and our findings suggest that they should be conserved.
Phonolites provide unique habitat conditions for plants on oceanic islands compared with surrounding areas. Higher species numbers and abundances as well as higher plant performance underline the importance of these rocks for the vegetation on oceanic islands. Despite the small total area covered by phonolites, they play a significant role in enhancing plant biodiversity on the island of La Palma. Our results suggest that exceptional rock outcrops like phonolites contribute to a better understanding of the formation of plant diversity on volcanic islands. As oceanic islands have always been formed through volcanic activity on the oceanic crusts, the combination of basaltic and phonolitic rocks is highly likely a regular pattern in Earth history.
Table A1References of selected chemical components of basaltic and phonolitic substrates. (B) indicates basaltic, (P) phonolitic, and (N) nepheline syenite (phonolite equivalent with larger grain size) substrates or treatments.
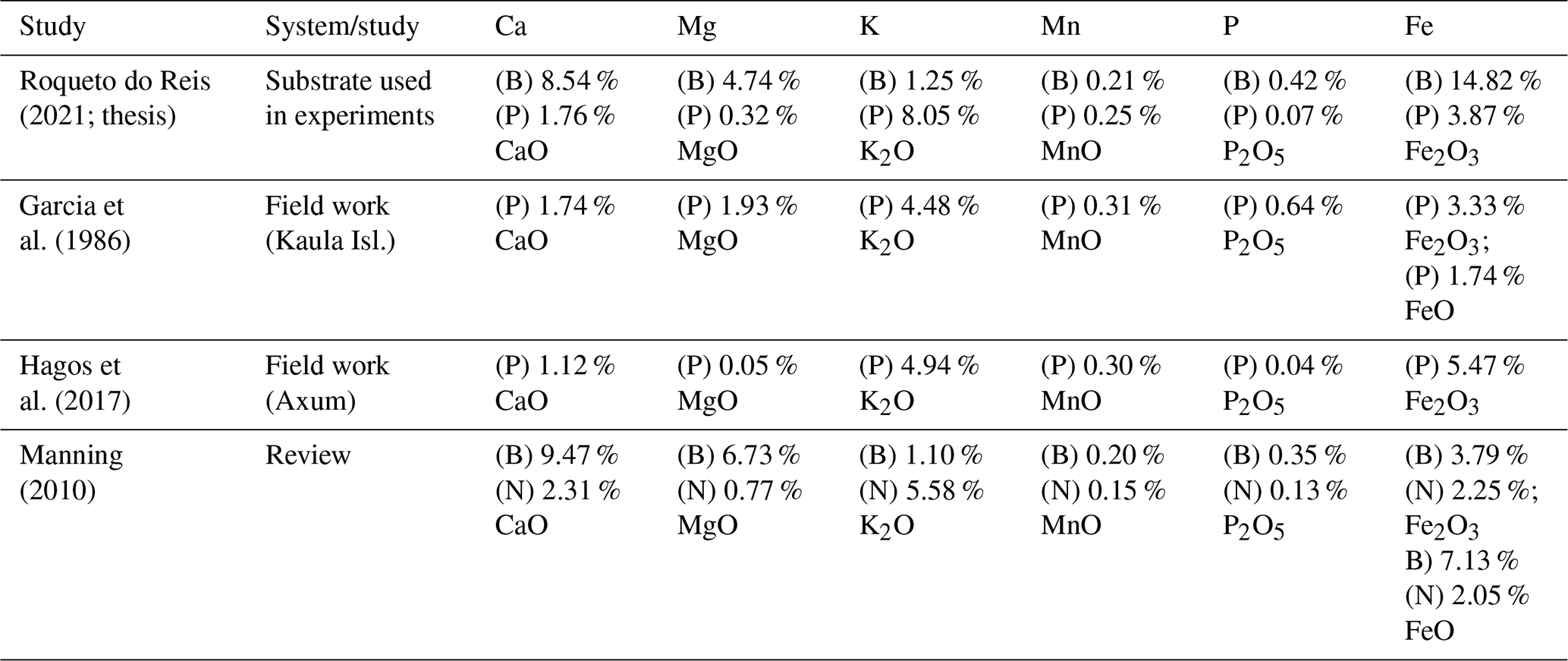
Table A2Interpolated data from climate models for our research sites based on CHELSA Climate Data (Karger et al., 2017).
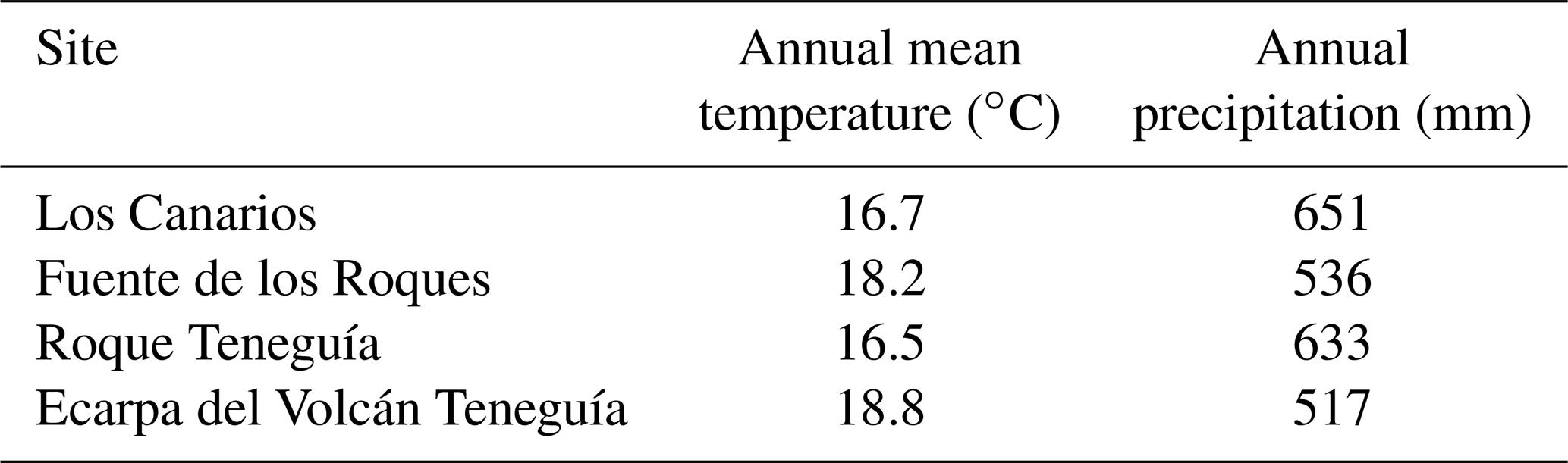
Table A3A post hoc permutation test (10 000 repetitions) between the DCA ordination axes and the environmental variables (substrate, inclination, aspect, and relief) showed no significant differences between phonolite and basalt. Obviously, the variation shown in the DCA does not depend on the substrate (but there is a relationship between northernness and the fourth dimension DCA4).

Table A4Complete list of all study species encountered on phonolites (P) and basalt (B) including their status as SIE, MIE, native (nat.), and introduced (intr.). The taxonomy follows the standards of Plants of the World Online (POWO 2019) updated and adapted to the FloCan Checklist (Beierkuhnlein et al., 2021).
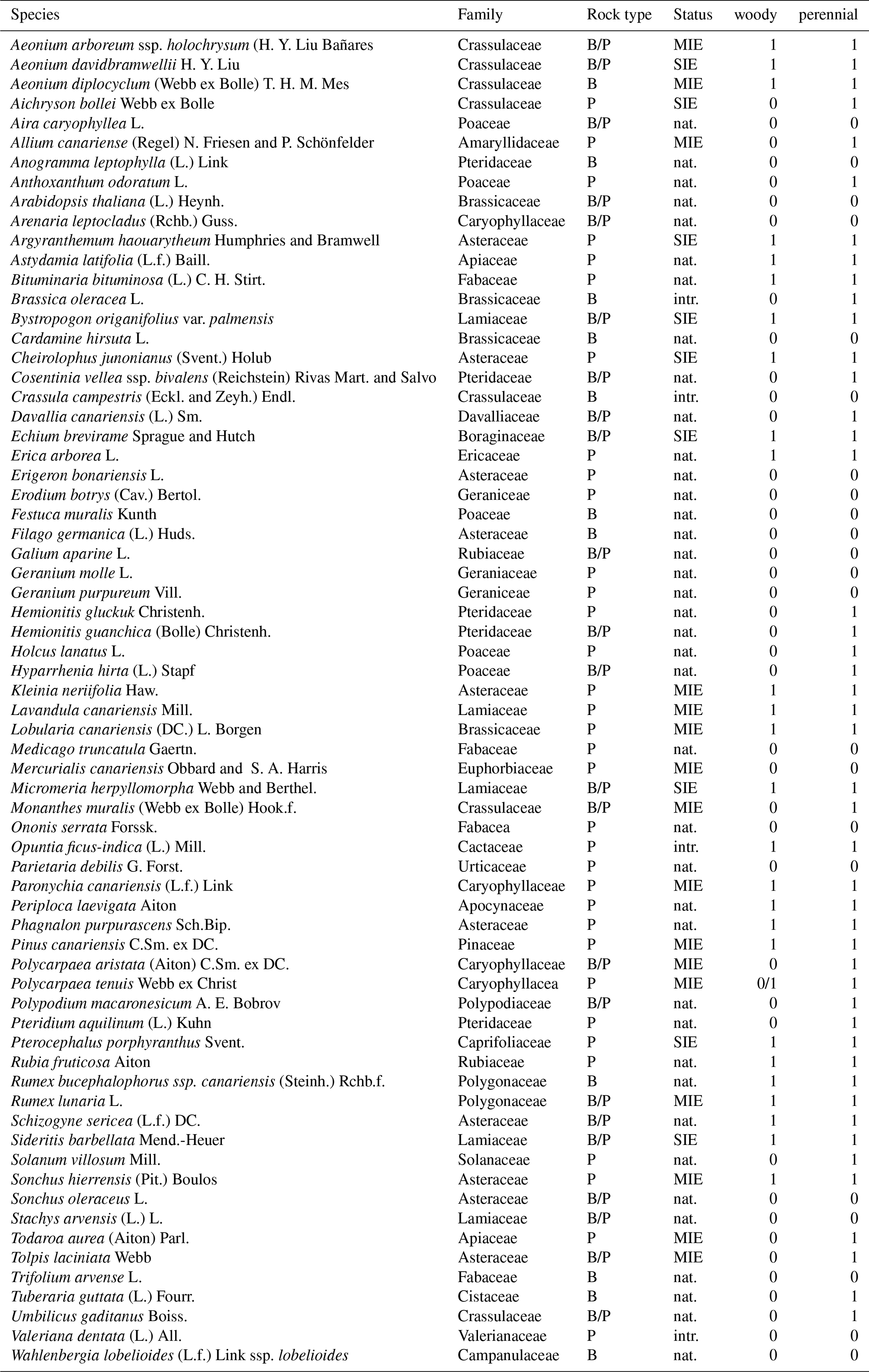
Table A5Environmental plot characteristics. Inclination on phonolites was (despite efforts to sample similar environments) significantly higher than on basalt. Components of exposition (northernness and easternness) and rugosity showed no significant differences (unpaired Whitney test).
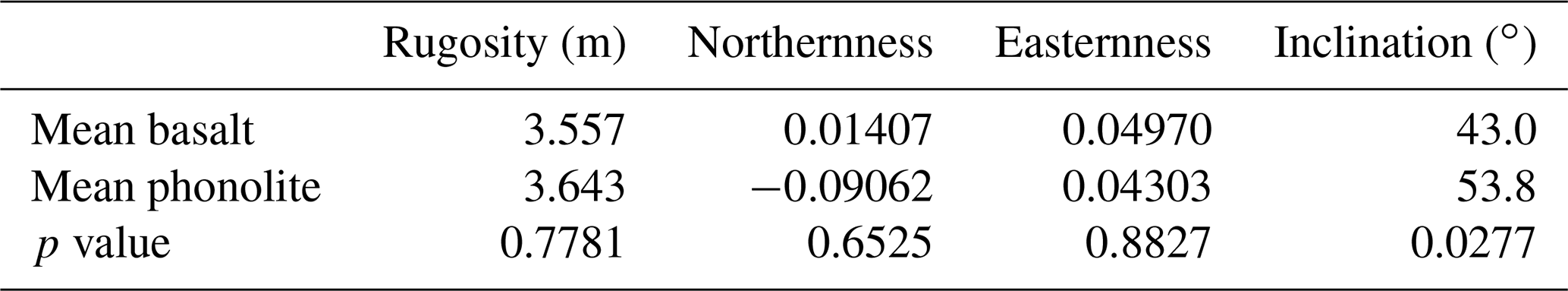
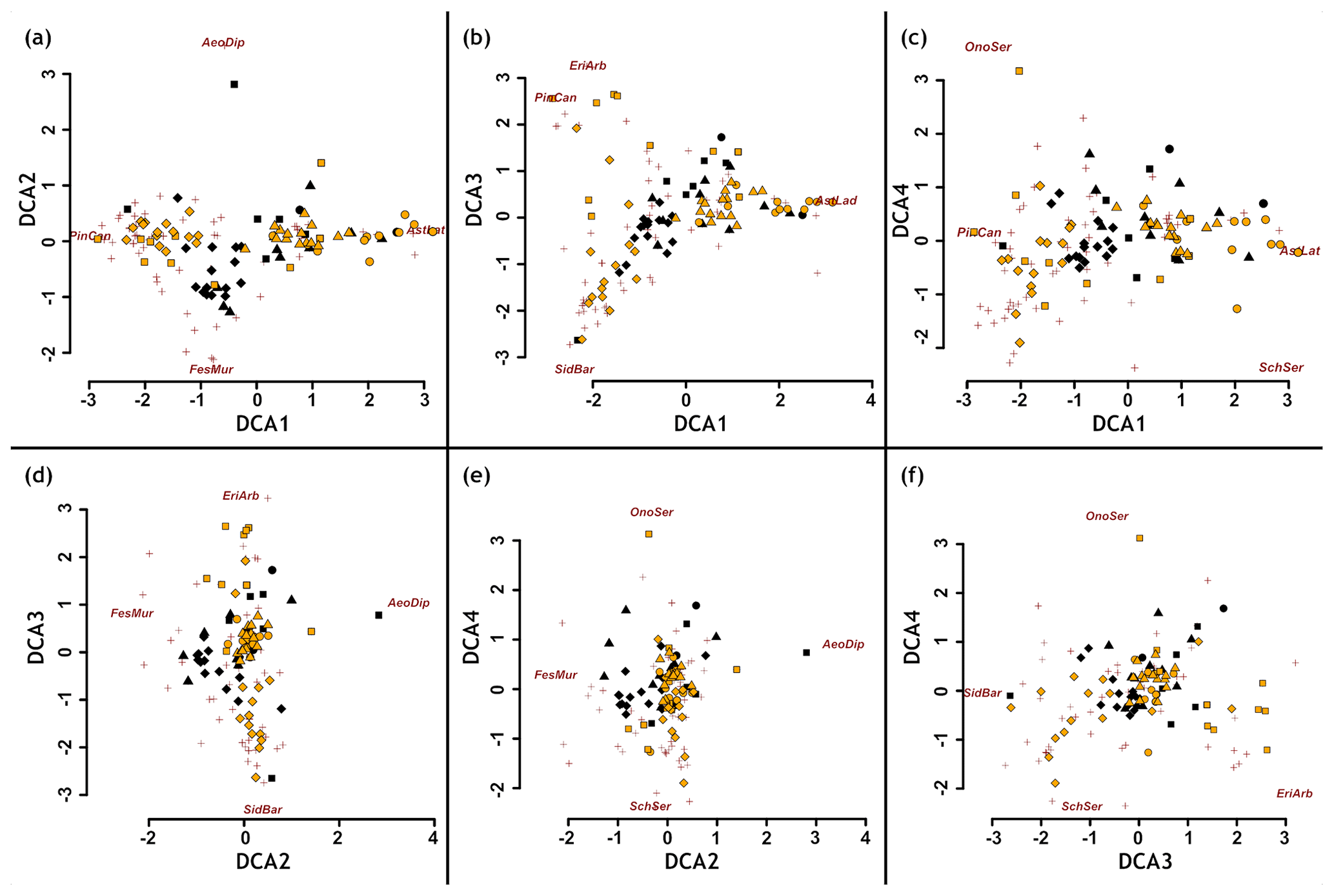
Figure A1Detrended correspondence analysis (DCA) shows no clear difference between phonolite and basaltic rock vegetation. Yellow dots show phonolite plots, black dots basalt plots, and dark red a subset of species centroids. Species names chosen based on the most extreme values along the gradients.
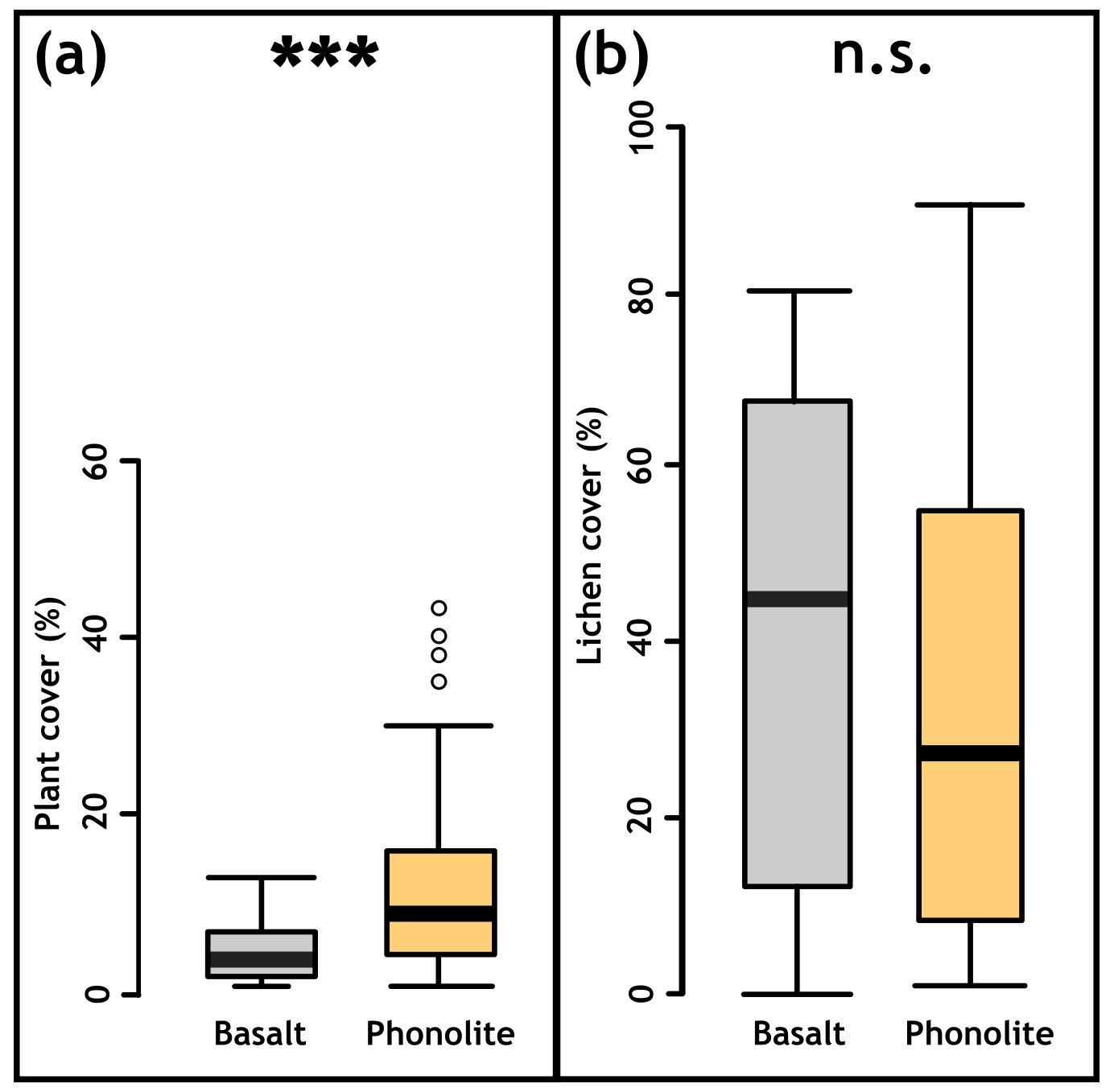
Figure A2Vegetation cover on basaltic and phonolite plots. (a) Plant cover showed significant differences between the substrates (p<0.001, Mann–Whitney U test) and (b) lichen cover showed no significant differences between the substrates (p>0.05, Mann–Whitney U test).
Only standard tests and plotting commands in R were used for data analysis. The code is available on request from the corresponding author.
Any data supporting the findings of this study are available within the Supplement of this article and were taken from Walentowitz et al. (2021; https://doi.org/10.1016/j.dib.2021.107229).
CB, SDHI, DK, LS, and AW developed the research idea, and DK, LS, and AW conducted the field work, analysed the data, and led the writing process. All authors developed the methods, discussed the results, and contributed to the manuscript.
The contact author has declared that neither they nor their co-authors have any competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
We would like to kindly thank the technical staff of the Biogeography Department of the University of Bayreuth for supporting this study and being of immense help in the implementation of field work. The Caldera de Taburiente National Park Directorate and especially Felix Medina from the Consejería de Medio Ambiente, Cabildo de La Palma, are thanked for permitting investigations in protected areas on La Palma and for their expertise on the local flora.
This research has been supported by Horizon 2020 (e-shape (grant no. 820852) and ECOPOTENTIAL (grant no. 641762)) and the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation (project no. 491183248)).
This open-access publication was funded by the University of Bayreuth.
This paper was edited by Frank Hagedorn and reviewed by two anonymous referees.
Abratis, M., Viereck, L., Pfänder, J. A., and Hentschel, R.: Geochemical composition, petrography and 40Ar/39Ar age of the Heldburg phonolite: implications on magma mixing and mingling, Int. J. Earth. Sci., 104, 2033–2055, https://doi.org/10.1007/s00531-015-1207-x, 2015.
Ackerman, L., Ulrych, J., Randa, Z., Erban, V., Hegner, E., Magna, T., Balogh, K., Frána, J., Milos, L., and Novák, J. K.: Geochemical characteristics and petrogenesis of phonolites and trachytic rocks from the České Středohoří Volcanic Complex, the Ohře Rift, Bohemian Massif, Elsevier Lithos, 224, 256–271, https://doi.org/10.1016/j.lithos.2015.03.014, 2015.
Alahuhta, J., Toivanen, M., and Hjort, J.: Geodiversity–biodiversity relationship needs more empirical evidence, Nat. Ecol. Evol., 4, 2–3, https://doi.org/10.1038/s41559-019-1051-7, 2020.
Atlantis 2021: Biodiversity Data Bank of the Canary Islands, Gobierno de las Canarias, 2021.
Bailey, J. J., Boyd, D. S., Hjort, J., Lavers, C. P., and Field, R.: Modelling native and alien vascular plant species richness: at which scales is geodiversity most relevant?, Glob. Ecol. Biogeogr., 26, 763–776, https://doi.org/10.1111/geb.12574, 2017.
Bañares, A., Blanca, G., Güemes, J., Moreno, J. C., and Ortiz, S. (Eds): Atlas y libro rojo de la flora vascular amenazada de España, Ministerio de Medio Ambiente, Madrid, ISBN 978-84-491-1071-9, 2004.
Barajas-Barbosa, M. P., Weigelt, P., Booregaard, M. K., Keppel, G., and Kreft H.: Environmental heterogeneity dynamics drive plant diversity on oceanic islands, J. Biogeogr., 47, 2248–2260, https://doi.org/10.1111/jbi.13925, 2020.
Beierkuhnlein, C., Walentowitz, A., and Welss, W.: FloCan – A revised checklist for the flora of the Canary Islands, Diversity, 13, 480, https://doi.org/10.3390/d13100480, 2021.
Brandmeier, M., Kuhlemann, J., Krumrei, I., Kappler, A., and Kubik, P. W.: New challenges for tafoni research. A new approach to understand processes and weathering rates, Earth Surf. Proc. Land., 36, 839–852, https://doi.org/10.1002/esp.2112, 2011.
Carracedo, J. C., Day, S. J., Guillou, H., and Gravestock, P.: Later stages of volcanic evolution of La Palma, Canary Islands: Rift evolution, giant landslides, and the genesis of the Caldera de Taburiente, GSA Bull., 111, 755–768, https://doi.org/10.1130/0016-7606(1999)111<0755:LSOVEO>2.3.CO;2, 1999.
Chiarucci, A., Robinson, B. H., Bonini, I., Petit, D., Brooks, R. R., and De Dominicis, V.: Vegetation of tuscan ultramafic soils in relation to edaphic and physical factors, Folia Geobot., 33, 113–131, https://doi.org/10.1007/BF02913340, 1998.
Chiarucci, A.: Vegetation ecology and conservation on Tuscan ultramafic soils, Bot. Rev., 69, 252–268, https://doi.org/10.1663/0006-8101(2003)069[0252:VEACOT]2.0.CO;2 , 2003.
Deák, B., Valkó, O., Török, P., Kelemen, A., Bede, A., Csathó, A. I., and Tóthmérész, B.: Landscape and habitat filters jointly drive richness and abundance of specialist plants in terrestrial habitat islands, Landscape Ecol., 33, 1117–1132, https://doi.org/10.1007/s10980-018-0660-x, 2018.
Eriksson, O.: Regional dynamics of plants: a review of evidence for remnant, source-sink and metapopulations, Oikos, 77, 248–258, https://doi.org/10.2307/3546063, 1996.
Eriksson, O.: Functional roles of remnant plant populations in communities and ecosystems, Glob. Ecol. Biogeogr., 9, 443–449, https://doi.org/10.1046/j.1365-2699.2000.00215.x , 2000.
Faccini, B., Di Giuseppe, D., Malferrari, D., Coltorti, M., Abbondanzi, F., Campisi, T., Laurora, A., and Passaglia, E.: Ammonium-exchanged zeolitite preparation for agricultural uses: from laboratory tests to large-scale application in ZeoLIFE project prototype, Periodico di Mineralogia, 84, 303–321, https://doi.org/10.2451/2015PM0015, 2015.
Field, R., Hawkins, B. A., Cornell, H. V., Currie, D. J., Diniz-Filho, J. A. F., Guégan, J. F., Kaufman, D. M., Kerr, J. T., Mittelbach, G. G., Oberdorff, T., O'Brian, E. M., and Turner, J. R. G.: Spatial species-richness gradients across scales: a meta-analysis, J. Biogeogr., 36, 132–147, https://doi.org/10.1111/j.1365-2699.2008.01963.x, 2009.
Formoso, M. L. L., Retzmann, K., and Valeton, I.: Fractionation of rare earth elements in weathering profiles on phonolites in the area of Lages, Santa Catarina, Brazil, Geochim. Brasil., 3, 51–61, 1989. Garnatje, T., Susanna, A., and Messeguer, R.: Isozyme studies in the genus Cheirolophus (Asteraceae: Cardueae-Centaureinae) in the Iberian Peninsula, North Africa and the Canary Islands, Pl. Syst. Evol., 213, 57–70, https://doi.org/10.1007/BF00988908, 1998.
Garcia, M., Fre, F., and Grooms, D.: Petrology of volcanic rocks from Kaula Island, Hawaii, Contrib. Mineral. Petr., 94, 461–471, https://doi.org/10.1007/BF00376339, 1986.
Gaston, K. J.: Global patterns in biodiversity, Nature, 405, 220–227, https://doi.org/10.1038/35012228, 2000.
Giladi, I., May, F., Ristow, M., Jeltsch, F., and Ziv, Y.: Scale-dependent species–area and species–isolation relationships: a review and a test study from a fragmented semi-arid agro-ecosystem, J. Biogeogr., 41, 1055–1069, https://doi.org/10.1111/jbi.12299, 2014.
Gillespie, R. G. and Roderick, G. K.: Geology and climate drive diversification, Nature, 509, 297–298, https://doi.org/10.1038/509297a, 2014.
Gray, M.: Geodiversity, Valuing and Conserving Abiotic Nature, John Wiley & Sons Ltd, Chichester, ISBN 0-470-84895-2, 2004.
Gray, M.: Comment, Other nature: geodiversity and geosystem services, Environ. Conserv., 38, 271–274, https://doi.org/10.1017/S0376892911000117, 2011.
Hagos, M., Koeberl, C., and Jourdan, F.: Geochemistry and geochronology of phonolitic and trachytic source rocks of the Axum obelisks and other stone artifacts Axum, Ethiopia, Geoheritage, 9, 479–494, https://doi.org/10.1007/s12371-016-0199-7, 2017.
Hall, K., Lindgren, B. S., and Jackson, R.: Rock albedo and monitoring of thermal conditions in respect of weathering: some expected and some unexpected results, Earth Surf. Proc. Land., 30, 801–811, https://doi.org/10.1002/esp.1189, 2005.
Harrison, S., Safford, H. D., Grace, J. B., Viers, J. H., and Davies, K. F.: Regional and local species richness in an insular environment: serpentine plants in California, Ecol. Monogr., 76, 41–56, https://doi.org/10.1890/05-0910, 2006.
Harrison, S. P. and Rajakaruna, N.: Serpentine: The Evolution and Ecology of a Model System, University of California Press, Berkeley, ISBN 978-0-520-26835-7, 2011.
Harter, D. E. V., Irl, S. D., Seo, B., Steinbauer, M. J., Gillespie, R., Triantis, K. A., Fernández-Palacios, J.-M., and Beierkuhnlein, C.: Impacts of global climate change on the floras of oceanic islands–Projections, implications and current knowledge, Perspect. Plant Ecol. Evol. Syst., 17, 160–183, https://doi.org/10.1016/j.ppees.2015.01.003, 2015.
Hjort, J., Gordon, J. E., Gray, M., and Hunter, M. L.: Why geodiversity matters in valuing nature's stage, Conserv. Biol., 29, 630–639, https://doi.org/10.1111/cobi.12510, 2015.
Hortal, J., Triantis, K. A., Meiri, S., Thébault, E., and Sfenthourakis, S.: Island species richness increases with habitat diversity, Am. Nat., 174, 207–217, https://doi.org/10.1086/645085, 2009.
Hulshof, C. M. and Spasojevic, M. J.: The edaphic control of plant diversity, Global Ecol. Biogeogr., 29, 1634–1650, https://doi.org/10.1111/geb.13151, 2020.
Irl, S. D. H., Harter, D. E. V., Steinbauer, M. J., Gallego Puyol, D., Fernández-Palacios, J. M., Jentsch, A., and Beierkuhnlein, C.: Climate vs. topography–spatial patterns of plant species diversity and endemism on a high-elevation island, J. Ecol., 103, 1621–1633, 2015.
Irl, S. D. H., Schweiger, A., Hoffmann, S., Beierkuhnlein, H., Pickel, T., and Jentsch, A.: Spatiotemporal dynamics of plant diversity and endemism during primary succession on an oceanic volcanic island, J. Veg. Sci., 30, 587–598, 2019.
Karger, D. N., Conrad, O., Böhner, J., Kawohl, T., Kreft, H., Soria-Auza, R. W., Zimmermann, N. E., Linder, P., and Kessler, M.: Climatologies at high resolution for the Earth land surface areas, Sci. Data, 4, 170122, https://doi.org/10.1038/sdata.2017.122, 2017.
Kazakou, E., Adamidis, G. C., Baker, A. J., Reeves, R. D., Godino, M., and Dimitrakopoulos, P. G.: Species adaptation in serpentine soils in Lesbos Island (Greece): metal hyperaccumulation and tolerance, Plant Soil, 332, 369–385, https://doi.org/10.1007/s11104-010-0302-9, 2010.
Kier, G., Kreft, H., Lee, T. M., Jetz, W., Ibisch, P. L., Nowicki, C., Mutke, J., and Barthlott, W.: A global assessment of endemism and species richness across island and mainland regions, P. Natl. Acad. Sci. USA, 106, 9322–9327, https://doi.org/10.1073/pnas.0810306106, 2009.
Kruckerberg, A. R.: An essay: Geoedaphics and island biogeography for vascular plants, Aliso, 13, 225–238, 1991.
Lawler, J. J., Ackerly, D. D., Albano, C. M., Anderson, M. G., Dobrowski, S. Z., Gill, J. L., Heller, N. E., Pressey, R. L., Sanderson, E. W., and Weiss, S. B.: The theory behind, and the challenges of, conserving nature's stage in a time of rapid change, Conserv Biol., 29, 618–629, https://doi.org/10.1111/cobi.12505, 2015.
Laughlin, D. C., Gremer, J. R., Adler, P. B., Mitchell, R. M., and Moore M., M.: The net effect of functional traits on fitness, Trends Ecol. Evol., 35, 1037–1047, https://doi.org/10.1016/j.tree.2020.07.010, 2020.
Liu, J., Möller, M., Gao, L., Poudel, R. C., and Li, D.: Geological and ecological factors drive cryptic speciation of yews in a biodiversity hotspot, New Phytol., 199, 1093–1108, https://doi.org/10.1111/nph.12336, 2013.
Liu, J., Matthews, T. J., Zhong, L., Liu, J., Wu, D., and Yu, M.: Environmental filtering underpins the island species-area relationship in a subtroppical anthropogenic archipelago, J. Ecol., 108, 424–432, https://doi.org/10.1111/1365-2745.13272, 2020.
MacArthur, R. H. and Wilson, E. O: The Theory of Island Biogeography, Monographs in Population Biology no. 1, Princeton University Press, Princeton, NJ, ISBN 978-1-4008-8137-6, 1967.
Manning, D. A. C.: Mineral sources of potassium for plant nutrition, A review, Agron. Sustain. Dev., 30, 281–294, https://doi.org/10.1051/agro/2009023, 2010.
Matthews, T. J., Guilhaumon, F., Triantes, K. A., Borregaard, M. K., and Whittaker, R. J.: On the form of species-area relationships in habitat islands and true islands, Global. Ecol. Biogeogr., 25, 847–858, https://doi.org/10.1111/geb.12269, 2016.
Middlemost, E. A. K.: San Miguel de La Palma – A volcanic island in section, Bull. Volcanol., 34, 216–239, https://doi.org/10.1007/BF02597787, 1970.
Middlemost, E. A. K.: Evolution of La Palma, Canary Archipelago, Contr. Mineral. Petrol., 36, 33–48, https://doi.org/10.1007/BF00372833, 1972.
Muer, T., Sauerbier, H., and Cabrera Calixto, F.: Die Farn- und Blütenpflanzen der Kanarischen Inseln, Margraf Publishers, Weikersheim, ISBN 978-3-8236-1721-1, 2016.
Nockolds, S. R.: Average chemical composition of some igneous rocks, Bull. Geol. Soc. Am., 65, 1007–1032, 1954.
Nogueira, T. A. R., Miranda, B. G., Jalal, A., Lessa, L. G. F., Filho, M. C. M. T., Marcante, N. C., Abreu-Junior, C. H., Jani, A. D., Capra, G. F., Moreira, A., and Martins, É. D. S.: Nepheline syenite and phonolite as alternative potassium sources for maize, Agronomy, 11, 1385, https://doi.org/10.3390/agronomy11071385, 2021.
Pankhurst, M. J., Scarrow, J. H., Barbee, O. A., Hickey, J., Coldwell, B. C., Rollinson, G., Rodriguez-Losada, J. A., Martín-Lorenzo, A., Rodríguez, F., Hernández, W., Hernández, P. A., and Pérez, N. M.: Petrology of the opening eruptive phase of the 2021 Cumbre Vieja eruption, La Palma, Canary Islands, https://doi.org/10.21203/rs.3.rs-963593/v1, 2021.
Paulay, M.: Biodiversity on oceanic islands: Its origin and extinction, Am. Zool., 34, 134–144, https://doi.org/10.1093/icb/34.1.134, 1994.
Pausas, J. G., Carreras, J., Ferré, A., and Font, X.: Coarse-scale plant species richness in relation to environmental heterogeneity, J. Veg. Sci., 14, 661–668, https://doi.org/10.1111/j.1654-1103.2003.tb02198.x, 2003.
Porder, S., Paytan, A., and Vitousek, P. M.: Erosion and landscape development affect plant nutrient status in the Hawaiian Islands, Oecologia, 142, 440–449, https://doi.org/10.1007/s00442-004-1743-8, 2004.
POWO: Plants of the World Online, Facilitated by the Royal Botanic Gardens, Kew, 2019: http://www.plantsoftheworldonline.org/, last access: 6 January 2021.
Ramos, L. A., Nolla, A., Korndörfer, G. H., Pereira, H. S., and Camargo, M. S.: Reatividade de corretivos da acidez e condicionadores de solo em colunas de lixiviação, Rev. Bras. Cienc. Solo., 30, 849–857, 2006.
Ricketts, T. H.: The matrix matters: Effective isolation in fragmented landscapes, Am. Nat., 158, 87–99, https://doi.org/10.1086/320863, 2001.
Rosenzweig, M.: Species Diversity in Space and Time, Cambridge: Cambridge University Press, https://doi.org/10.1017/CBO9780511623387, 1995.
Roqueto de Reis, B.: Rock powders in the soil-plant system: mineralogy and microbiome response, Master Thesis, University of São Paulo, https://doi.org/10.11606/D.11.2020.tde-31032021-161123, 2021.
Schoen, C., Aumond, J. J., and Stuermer, S. L.: Efficiency of the on-Farm mycorrhizal inoculant and phonolite rock on growth and nutrition of Schinus terebinthifolius and Eucalyptus saligna, Rev. Bras. Cien. Solo., 40, 1–14, https://doi.org/10.1590/18069657rbcs20150440, 2016.
Spürgin, S., Weisenberger, T. B., and Marković, M.: Zeolite-group minerals in phonolite-hosted deposits of the Kaiserstuhl Volcanic Complex, Germany, Am. Mineral., 104, 659–670, https://doi.org/10.2138/am-2019-6831, 2019.
Tavares, L. F., de Carvalho, A. M. X., Camargo, L. G. B., de Fátima Pereira, S. G., and Cardoso, I. M.: Nutrients release from powder phonolite mediated by bioweathering actions, Int. J. Recycl. Org. Waste. Agr., 7, 89–98, https://doi.org/10.1007/s40093-018-0194-x, 2018.
Tukiainen, H., Bailey, J. J., Field, R., Kangas, K., and Hjort, J.: Combining geodiversity with climate and topography to account for threatened species richness, Conserv. Biol., 31, 364–375, https://doi.org/10.1111/cobi.12799, 2016.
Vitales, D., Garnatje, T., Pellicer, J., Vallès, J., Santos-Guerra, A., and Sanmartín, I.: The explosive radiation of Cheirolophus (Asteraceae, Cardueae) in Macaronesia, BMC Evol. Biol., 14, 1–15, https://doi.org/10.1186/1471-2148-14-118, 2014a.
Vitales, D., García-Fernández, A., Pellicer, J., Vallè, J., Santos-Guerra, A., Cowan, R. S., Fay, M. F., Hidalgo, O., and Garnatje, T.: Key processes for Cheirolophus (Asteraceae) diversification on oceanic islands inferred from AFLP Data, Plos ONE, 9, e113207, https://doi.org/10.1371/journal.pone.0113207, 2014b.
von Fragstein, P., Pertl, W., and Vogtmann, H.: Artificial weathering of silicate rock powders, Z. Pflanz. Bodenkunde, 151, 141–146, https://doi.org/10.1002/jpln.19881510214, 1988.
von Wilpert, K. and Lukes, M.: Ecochemical effects of phonolite rock powder, dolomite and potassium sulfate in a spruce stand on an acidified glacial loam, Nutr. Cycl. Agroecosys., 65, 115–127, https://doi.org/10.1023/A:1022103325310, 1998.
Walentowitz, A., Kienle, D., Sungur, L., and Beierkuhnlein, C.: Vegetation plot and trait data from phonolitic and basaltic rocks on La Palma (Canary Islands, Spain), [data set], 37, 107229, https://doi.org/10.1016/j.dib.2021.107229, 2021.
Whittaker, R. J. and Fernández-Palacios, J.-M.: Island Biogeography: Ecology, Evolution, and Conservation, 2 Edn., Oxford University Press, Oxford, ISBN 0-19-856611-5, 2007.