the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 30 Aug 2022
| 30 Aug 2022
Modeling the effects of alternative crop–livestock management scenarios on important ecosystem services for smallholder farming from a landscape perspective
Mirjam Pfeiffer
Munir P. Hoffmann
Simon Scheiter
William Nelson
Johannes Isselstein
Kingsley Ayisi
Jude J. Odhiambo
Reimund Rötter
Smallholder farming systems in southern Africa are characterized by low-input management and integrated livestock and crop production. Low yields and dry-season feed shortages are common. To meet growing food demands, sustainable intensification (SI) of these systems is an important policy goal. While mixed crop–livestock farming may offer greater productivity, it implies trade-offs between feed supply, soil nutrient replenishment, soil carbon accumulation, and other ecosystem functions (ESFs) and ecosystem services (ESSs). Such settings require a detailed system understanding to assess the performance of prevalent management practices and identify potential SI strategies. Models can evaluate different management scenarios on extensive spatiotemporal scales and help identify suitable management strategies. Here, we linked the process-based models APSIM (Agricultural Production Systems sIMulator) for cropland and aDGVM2 (Adaptive Dynamic Global Vegetation Model) for rangeland to investigate the effects of (i) current management practices (minimum input crop–livestock agriculture), (ii) an SI scenario for crop production (with dry-season cropland grazing), and (iii) a scenario with separated rangeland and cropland management (livestock exclusion from cropland) in two representative villages of the Limpopo Province, South Africa, for the period from 2000 to 2010. We focused on the following ESFs and ESSs provided by cropland and rangeland: yield and feed provision, soil carbon storage, cropland leaf area index (LAI), and soil water. Village surveys informed the models of farming practices, livelihood conditions, and environmental circumstances. We found that modest SI measures (small fertilizer quantities, weeding, and crop rotation) led to moderate yield increases of between a factor of 1.2 and 1.6 and reduced soil carbon loss, but they sometimes caused increased growing-season water limitation effects. Thus, SI effects strongly varied between years. Dry-season crop residue grazing reduced feed deficits by approximately a factor of 2 compared with the rangeland-only scenario, but it could not fully compensate for the deficits during the dry-to-wet season transition. We expect that targeted deficit irrigation or measures to improve water retention and the soil water holding capacity may enhance SI efforts. Off-field residue feeding during the dry-to-wet season transition could further reduce feed deficits and decrease rangeland grazing pressure during the early growing season. We argue that integrative modeling frameworks are needed to evaluate landscape-level interactions between ecosystem components, evaluate the climate resilience of landscape-level ecosystem services, and identify effective mitigation and adaptation strategies.
- Article
(6465 KB) - Full-text XML
-
Supplement
(3292 KB) - BibTeX
- EndNote





Smallholder farms occupy approximately 62 % of Africa's farmland area. Family-driven labor generates household income and food security (FAO, 2014) and heavily relies on natural ecosystem functions (ESFs) and ecosystem services (ESSs), including the provision of yield and biomass as services provided by nature that benefit farmers and their livelihoods (Costanza et al., 1997; Millennium Ecosystem Assessment, 2005). Low income and education levels, poor access to markets and credits, lack of technology, and strong dependence on external support from governments and non-governmental organizations in the form of safety nets, advisory support, off-farm income such as pensions, or donations, are challenges that endanger food security, welfare, and the well-being of smallholders (FAO, 2015). These obstacles impede the capacity to mitigate and adapt to climate change (Harvey et al., 2013). Livestock husbandry and subsistence cropping characterize smallholder farming in Africa (Thornton and Herrero, 2015; Descheemaeker et al., 2016). Livestock provide milk, meat, and leather; convey prestige; and contribute to economic diversification. Livestock often feed on crop residues during the dry period, which allows rangeland resting, accelerates nutrient recycling, and links both land use types. Although integrated crop–livestock farms may offer greater farming efficiency and sustainability (Sumberg, 2003; Herrero et al., 2009; Tarawali et al., 2011), they often lead to various trade-offs (Herrero et al., 2009; Erenstein, 2002). Harvest residues as fodder reduce feed gaps, whereas leaving residues on-field allows for nutrient replenishment and enhances soil fertility (Castellanos-Navarrete et al., 2014; Valbuena et al., 2012). However, freshly excreted nutrients from livestock are not used during dry-season fallow, and considerable nutrient losses can occur (Hack-ten Broeke et al., 1996).
Climatic variability creates additional challenges. Southern Africa experiences high interannual rainfall variability due to the El Niño–Southern Oscillation phenomenon (Conway et al., 2015). Increased stocking density during high-rainfall years results in forage shortages in subsequent dry years. Over-stocking without feed supplementation then results in severe pressure on the drought-afflicted vegetation and leads to animal malnutrition, increased livestock mortality, economic losses, and threats to the ESSs provided by rangelands (Müller et al., 2015). Overgrazing may lead to rangeland degradation, species loss, reduced carbon transfer to soils, and habitat deterioration. Higher bare-ground fractions increase runoff, soil erosion, and evaporation.
Focusing on the multi-functionality and complex interactions of mixed cropland–rangeland systems is necessary to evaluate the performance of prevalent cropping and livestock husbandry practices and identify possible site-specific sustainable intensification strategies (Giller et al., 2006; Rusinamhodzi et al., 2011). Observational assessments of different management options require numerous field experiments, which are often limited by time and resource constraints. In contrast, models can systematically explore different management scenarios on various spatiotemporal scales and help identify suitable management strategies once they have been evaluated satisfactorily (Kersebaum et al., 2015). Crop simulation models (CSMs) simulate the effects of different management strategies on biomass, yield, water use, and nutrient uptake for numerous combinations of genotype and environment interactions (see, e.g., Rötter and Van Keulen, 1997; Whitbread et al., 2010). Livestock models that simulate animal productivity (e.g., meat and milk) depending on age, gender, and status (e.g., juvenile, pregnant, or lactating; see van de Ven et al., 2003) can integrate the output of such CSMs. For example, Descheemaeker et al. (2018) used the APSIM (Agricultural Production Systems sIMulator) crop model (Holzworth et al., 2014) and the LIVSIM (LIVestock SIMulator) livestock model (Rufino et al., 2015) within the Agricultural Model Intercomparison and Improvement Project (AgMIP) framework (Rosenzweig et al., 2013) to investigate climate change effects on forage availability as well as livestock and crop productivity. Dynamic vegetation models (DVMs) simulate natural vegetation dynamics, carbon sequestration, energy and water fluxes, and biomass production in response to environmental drivers and disturbances. Recent developments have increasingly focused on anthropogenic influences and aim to include management. For example, the aDGVM (Adaptive Dynamic Global Vegetation Model), a DVM developed for African savanna ecosystems (Scheiter and Higgins, 2009), has been expanded to simulate fuelwood harvesting and grazing in order to determine how climate change influences the economic value of ESSs in southern African rangelands (Scheiter et al., 2019). New routines in the trait-based aDGVM2 allow for an improved representation of grass-layer diversity and simulation of selective grazing (Pfeiffer et al., 2019). Currently, livestock in aDGVM2 are not represented as an interactive agent, i.e., the model simulates livestock impact on vegetation but cannot simulate herd-related aspects such as decision-based animal movement, growth, metabolism, reproduction, or nutrition status in the manner of an agent-based model such as RaMDry (Rangeland Model in Drylands; Fust and Schlecht, 2018).
While assessments of ESFs in crop–livestock systems are crucial for smallholder farming communities in sub-Saharan Africa (Descheemaeker et al., 2016; Waha et al., 2018), CSMs and DVMs commonly consider cropland and rangeland independently. While such applications improve the individual system understanding, only a coupled framework that combines CSMs and DVMs can capture landscape-scale interactions. In this study, we linked APSIM with aDGVM2, two models that are both well tested for southern Africa (Hoffmann et al., 2018a, b, 2020; Pfeiffer et al., 2019), in order to address the following research questions for a landscape-scale study including two villages in South Africa's Limpopo Province:
-
Do village-specific sustainable intensification measures improve ecosystem services and functions compared with status quo land use practice and, if so, which functions and services are affected?
-
Does sustainable intensification lead to stronger positive effects on environmentally constrained sites compared with environmentally more favorable sites?
-
Do feed gaps occur and are there village-specific differences?
-
Can joint management of cropland and rangeland reduce or eliminate feed gaps?
-
Can integrated modeling of cropland and rangeland identify management strategies that result in a sustained provision of landscape-level ecosystem functions and services?
Simulations of cropland and rangeland dynamics for the two villages were coupled off-line to test the impacts of different management scenarios on those landscape-scale ESSs that are (a) particularly relevant for smallholder farmers in the region and (b) can be quantified by the coupled modeling framework. These include crop yield, provision of biomass for animal consumption from rangeland and cropland, carbon input to cropland soils and cropland soil organic carbon (SOC) formation and sequestration, soil water availability for crops, and leaf area index (LAI) as a measure to describe soil cover and, implicitly, protection against erosion and potential photosynthetic capacity at the stand level. The explicit evaluation of additional ESFs such as runoff, evapotranspiration, or species diversity was not part of the current study, although we acknowledge them as relevant. We focused on the following scenarios: the current status quo (minimum input integrated crop and livestock agriculture), a minimum sustainable intensification scenario, and separated vs. combined cropland–rangeland management. The APSIM model delivered results on yield, biomass productivity, carbon sequestration, and water use. We then used this output to compare feed demand with simulated harvest residues to determine whether cropland can cover the feed demand during dry seasons. The aDGVM2 simulated rangeland vegetation dynamics and biomass consumption during periods when livestock had no cropland access. The aDGVM2 output showed whether rangeland could satisfy animal demand during those periods. Moreover, seasonal and interannual high-risk times for feed deficits were determined. Such an integrative analysis combines the strengths of crop and rangeland models and is the first attempt in this form.
2.1 Study region
South Africa's Limpopo Province has a high share of smallholder farmers who often experience food insecurity. Maize is a staple crop, accompanied by legumes, including peanut, bambara nut, and cowpea, and some tubers. Cropping systems are low input with limited or no fertilizer application, and farmers typically broadcast seeds. Climate conditions range from a warm desert climate in the west to a warm semiarid to humid climate in the east (Engelbrecht and Engelbrecht, 2016). Approximately 80 % of annual precipitation falls during the cropping season from October to April/May, followed by a May–October dry season. We selected two villages representing the climate extremes for arable farming in the province: Gabaza and Selwana (802 and 585 mm mean annual precipitation, MAP, respectively, during the 2000–2010 period; Ruane et al., 2015). Average daily maximum temperatures are 30.6 and 32.4 ∘C in January and 24.3 and 25.9 ∘C in July for Gabaza and Selwana, respectively (Table 1). Both villages are part of research projects (SALLnet, Rötter et al., 2021, https://www.uni-goettingen.de/en/592566.html, last access: 24 August 2022) and have been under survey since 2013 (Table 1).
Table 1Site characteristics of the two selected study villages and cropland usage (% of cropland area) for the most common crop types according to village surveys conducted in 2019.
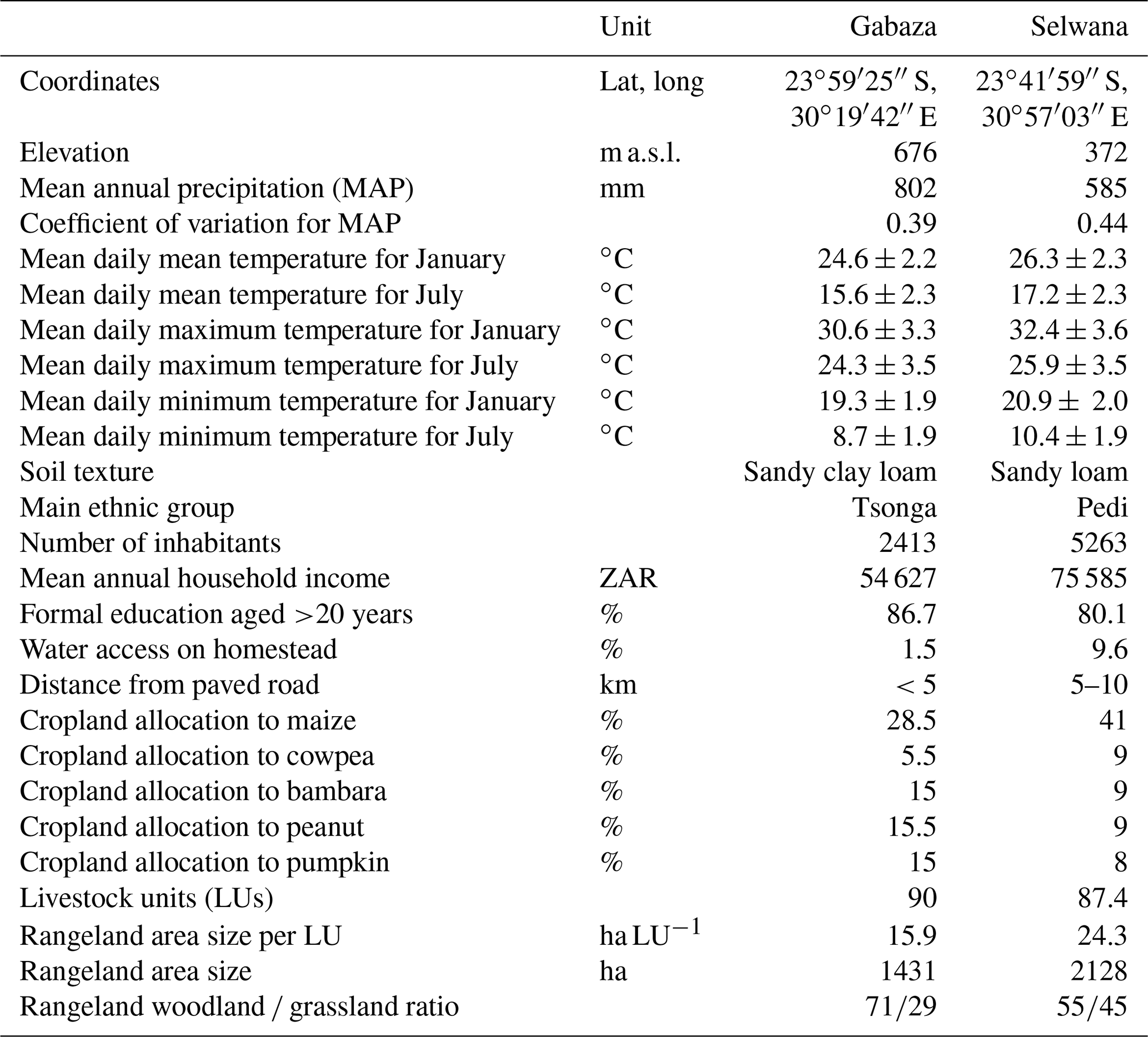
2.2 Current smallholder farming practices vs. a minimum sustainable intensification scenario
Farmers in Limpopo have adapted their crop–livestock practices to the strong climatic seasonality. Cattle graze on the communal rangeland during the cropping season. After crop harvest, when rangeland provides little feed, livestock frequently consume the remaining crop residues on-site (Bennett et al., 2009). Sometimes herders destroy fences to give cattle access to fields, potentially causing conflicts with crop farmers. We characterized village-specific cropping patterns based on a survey and ground-truthing campaign conducted in April and May 2019 and on background information based on working with smallholders in the region for >20 years. Table 1 summarizes the cropping patterns. More than 90 % of the farmers individually cultivate <1 ha and typically receive land usage rights from the community (permission to occupy, PTO). Maize is the prevailing regional staple (Table 1) and is complemented by legumes (e.g., peanuts and cowpea). Smallholders often recycle maize seeds from the previous harvest. Seeds are broadcast, and planting density is low, frequently ranging between 1 and 5 plants m−2. While mineral fertilizers are uncommon, cattle feeding on harvest residues accelerates nutrient cycling and concentrates nutrients in dropped manure. Weeds are widespread and are sporadically removed by hand. Farmers do not use agricultural chemicals nor machinery. Maize yields are often <1 t ha−1 (where “t” denotes metric tons), with yields of 2–3 t ha−1 in good years (manure input and regular weeding). We termed this “status quo” of cropland cultivation the “SQ scenario”.
For the minimum sustainable intensification scenario, termed “SI scenario”, we prescribed 50 kg ha−1 yr−1 of N-fertilizer application at sowing to increase the nutrient supply, but we did not consider any phosphorus or potassium application. Additionally, we implemented weeding during the crop growth period as well as crop rotation (maize followed by a legume, either peanut or cowpea) in order to avoid soil exhaustion. APSIM has been tested for this regionally common crop rotation practice (Hoffmann et al., 2020). The frequency of a crop within the rotation was determined by the land allocation of the crop as observed in the field. Cattle had access to the cropland in the dry season (postharvest) in both the SQ and SI scenarios, and they provided dung input, which was parameterized as a daily constant input value, that was considered in APSIM simulations as a nutrient input source. Dung collection on rangeland and the transfer of dung from rangeland to cropland was not included in simulations, as this practice is not common in either study village. This represents a low level of intensification, although it is an improvement on the current status quo. We deem it realistic that our assumptions for sustainable intensification are feasible for smallholder farmers in the target villages, even under current resource constraints, as farmers interviewed in surveys indicated that similar efforts are already made in neighboring villages.
2.3 Cattle and rangeland management according to village surveys
During the 2019 survey, local guides approached herders to assess cattle number, age, gender, and breed in order to establish village-specific feed demand and rangeland usage habits. At Selwana, the rangeland consisted of well-fenced “herder camps” where herders and cattle lived semipermanently and animals stay on the rangeland overnight. In contrast, in Gabaza, animals grazed on rangeland during the daytime and returned to the homestead at night to reduce the risk of theft. In Gabaza, brief interviews with herders were conducted in the morning before they sent livestock to the rangeland, which also allowed for the visual assessment of the herds. As tracking the herds with GPS collars was not possible, interviewers asked herders to delineate grazing areas on maps. For the parameterization of the rangeland simulations, we determined the daily village-level dry matter demand based on the livestock units (LUs), assuming a temporally constant daily dry matter demand of 12.5 kg per LU because aDGVM2 cannot currently quantify temporal changes in biomass quality. For APSIM simulations, we additionally calculated the monthly energy and protein demand of herds based on the surveys to estimate the nutrition-based input of manure to cropland. For this, we parameterized dry matter content, metabolizable energy, crude protein, and dry matter digestibility using the values established by Descheemaeker et al. (2018, see their Table 1).
Differing stocking density between villages is represented in aDGVM2 simulations via the village-specific daily biomass demand of herds. The differing spatiotemporal distribution of grazing pressure between both villages due to the subdivision of the rangeland area at Gabaza into four spatially separated sub-areas vs. the contiguous rangeland area at Selwana is also considered in aDGVM2 simulations. While we simulated continuous grazing of the rangeland area at Selwana during cattle presence, we explicitly simulated grazing on sub-areas in Gabaza by subdividing the total rangeland grazing time into four periods. However, due to the lack of sub-daily resolution in the grazing module of aDGVM2, we could not account for differing daytime–nighttime livestock handling between villages.
2.4 Coupling APSIM and aDGVM2
We coupled the APSIM crop growth model and the aDGVM2 vegetation model off-line. Both models conduct simulations at a daily resolution, and we used the same environmental input data to drive both models. APSIM simulations were conducted prior to the aDGVM2 simulations to determine (1) the crop residue quantities available for livestock during the dry season and (2) the timing of sowing and harvest to establish the times when cattle could access cropland in the RC scenario. The cropland grazing time windows established through the APSIM simulations were then used to exclude livestock from rangeland in aDGVM2 (no grazing in aDGVM2 simulated during these time windows) in the RC scenario.
2.5 Crop growth simulation using APSIM
We simulated crop growth and development using the Agricultural Production Systems sIMulator (APSIM v7.9, https://www.apsim.info, last access: 24 August 2022), which is partly based on early modeling work conducted in Kenya (Keating et al., 2003). Crop growth and development in APSIM are calculated on a daily time step and are affected by temperature, radiation, and water and nutrient supply. APSIM also calculates the dynamics of soil water, nitrogen, and phosphorus on a daily basis. Model outputs include cardinal physiological stages, such as duration of flowering and physiological maturity; total above- and belowground biomass; yield; water balance components, such as evapotranspiration and soil moisture at different depths of the root zone; nitrogen uptake; and nitrate leaching (Probert et al., 1998; Wang et al., 2014; Whitbread et al., 2017). APSIM has previously been calibrated and validated for sites and target crops in the study region; see, for example, Hoffmann et al. (2018b) and Nelson et al. (2022). These evaluations include a wide range of crops and crop rotations. While Hoffmann et al. (2018b) looked specifically at peanut crops, Nelson et al. (2022) focused on the calibration of maize for the study sites and two additional villages in the Mopani District, Limpopo Province. A number of complex crop rotations were tested at sites in South Africa by Hoffmann et al. (2020). For a general overview of APSIM applications in Africa, see Whitbread et al. (2010). For information on APSIM performance regarding different crops and functional aspects, see the studies listed in Table S1.
2.6 The APSIM simulation setup
We ran APSIM for the period from 1 October 1998 to 1 October 2010 using AgMERRA climate data (Ruane et al., 2015). Sowing took place in a time window between November and December once cumulative rainfall reached 20 mm within a 5 d period. Crops were harvested in April/May (2 weeks prior to the onset of the animal presence period on cropland in the RC scenario; see Fig. 1e and f and Fig. 2). We simulated soil water, soil organic carbon (SOC), N, and P continuously between cropping cycles. SOC was halved every 30 cm (see Dagliesh et al., 2016) and simulated to a soil depth of 180 cm, with constant soil texture below the root zone, based on SOC starting values measured during a 2015 field campaign. Simulated crops included maize (sc501), peanut (kangwana), and cowpea (banjo), the most common local types of each crop (see, e.g., Hoffmann et al., 2018b; Rapholo et al., 2019; Hoffmann et al., 2020). We used a generic summer grass and a winter dicot to simulate weeds. Weeds in the SI scenario were removed at a weed biomass threshold of 2 t ha−1 but at a maximum of 30 d (up to three times during the cropping season) to emulate findings from the ground-truthing campaign (see Sect. 2.2). We simulated two scenarios: (i) the average farmers' practice observed in the villages (i.e., “status quo”, termed SQ scenario) and (ii) a minimum sustainable intensification scenario (termed SI scenario), according to the specifications defined in Sect. 2.2.
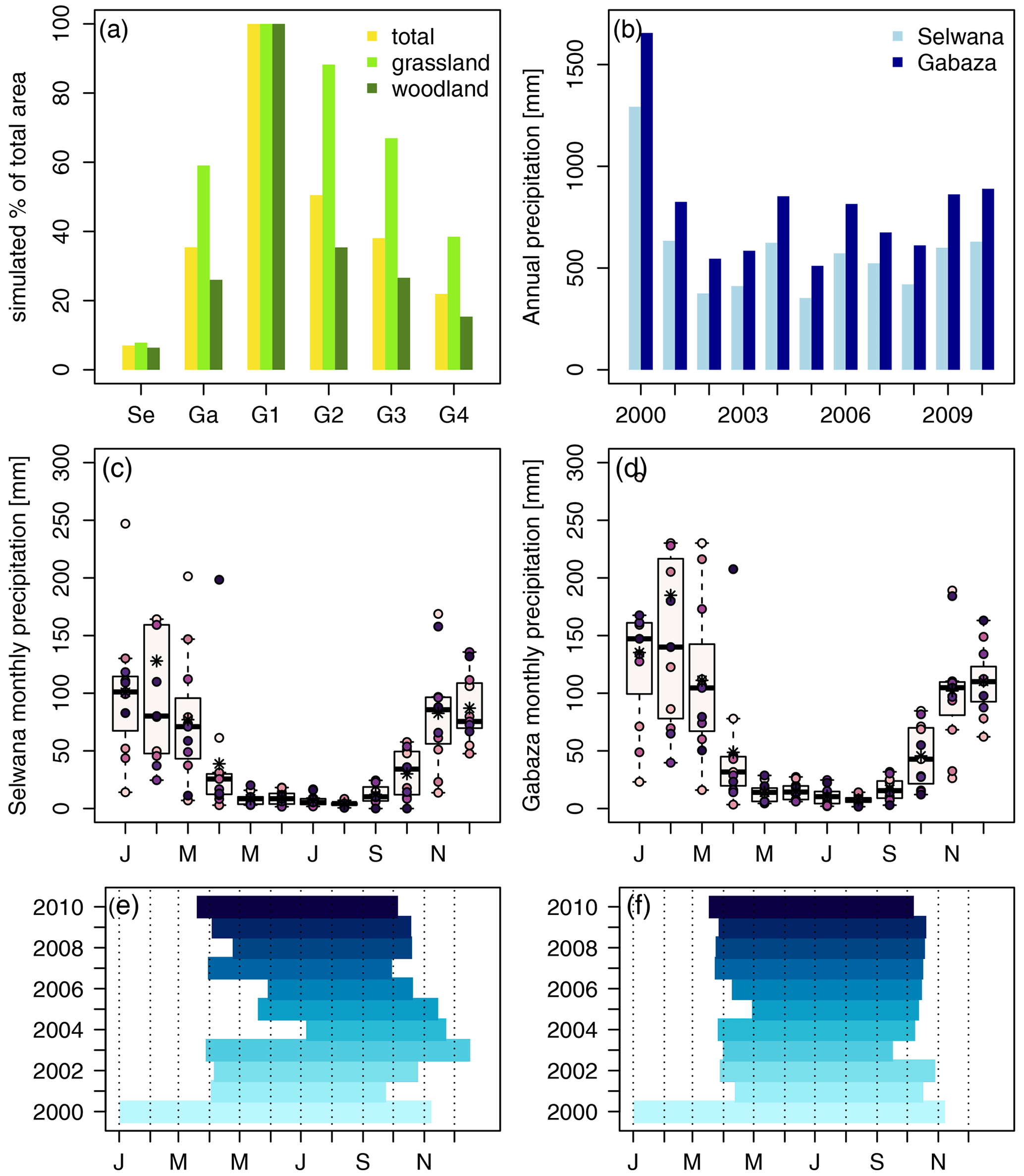
Figure 1Panel (a) shows the simulated area percentages for the different sites and (sub-)areas, and panel (b) presents the annual precipitation for the years 2000–2010 for Selwana and Gabaza from the AgMERRA climatology used to drive the APSIM and aDGVM2 simulations. Panels (c) and (d) show monthly precipitation at Selwana and Gabaza, respectively; stars indicate the 2000–2010 average, and the dots represent the individual annual values. Panels (e) and (f) show the annual time lines of animal presence on cropland in the RC scenario. A maximum of 150 ha (75 ha of grassland and 75 ha of woodland) was simulated per (sub-)area, amounting to the shown simulated percentages of total area. The abbreviations used in the figure are as follows: SE – Selwana and Ga – Gabaza. G1–G4 indicate sub-areas for Gabaza. For sub-area G1 (total size of 57 ha), we simulated all individual hectares of the sub-area.
2.7 Modeling rangeland dynamics
The aDGVM2 simulates the daily growth and state of individual plants on representative 1 ha stands (Scheiter et al., 2013; Langan et al., 2017). Trait sets describe each individual's growth form (grass, tree, shrub, perennial, or annual grass), leaf characteristics (specific leaf area and photosynthetic pathway), carbon allocation to plant compartments, plant architecture (roots and crown shape), response to fire, reproduction, and mortality. Plant performance emerges from trait characteristics and environmental filtering (Scheiter et al., 2013; Langan et al., 2017; Pfeiffer et al., 2019). The model has been specifically developed and tested for conditions in southern Africa (Pfeiffer et al., 2019). To represent grass functional diversity and grazing impacts, the model simulates annual and perennial grasses and accounts for preferential grazing (see the model description in Pfeiffer et al., 2019).
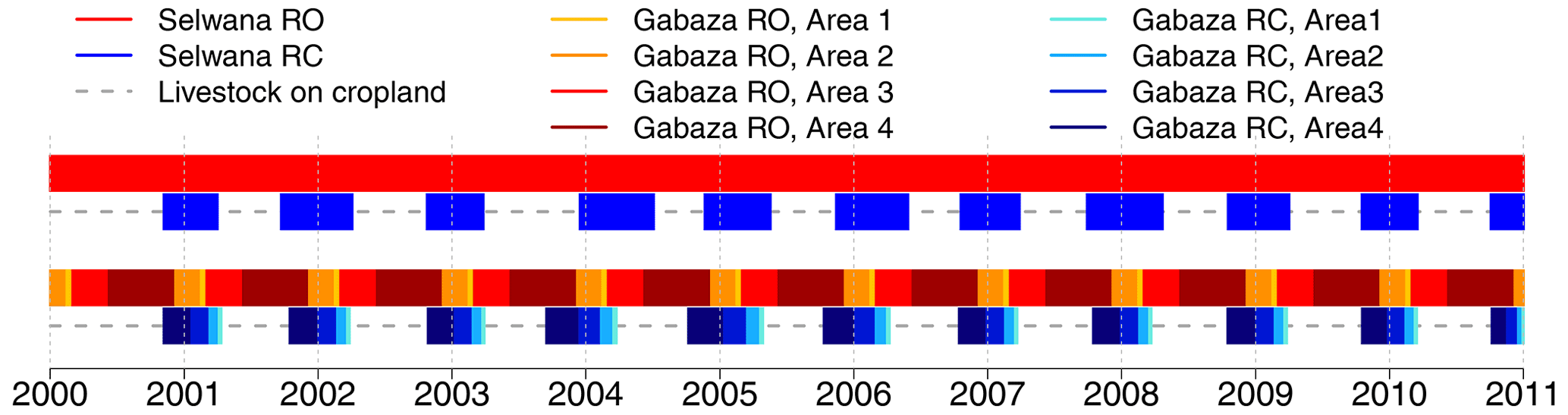
Figure 2Periods of livestock presence on communal rangeland. Bars with a red hue illustrate periods of rangeland presence in the RO scenario, bars with a blue hue show the rangeland presence/crop growth period in the RC and SQ scenarios, and dashed gray lines indicate animal presence on cropland. For Gabaza (bottom pair of bar sequences), the different hues show animal presence in the four sub-areas. The period of animal presence in sub-areas is proportional to the sub-area size.
2.7.1 The aDGVM2 simulation setup
The aDGVM2 required animal presence times, the number of livestock units (LUs), and the dry matter demand per LU to determine daily hectare-based biomass removal. We assumed that animals move from cropland to rangeland 2 weeks before planting and return 2 weeks after the completion of the crop harvest to feed on crop residues during the dry season. SI or SQ crop management did not influence the timing of rangeland grazing simulations, as the timing of sowing and harvest maximum varied by a few days.
We simulated three different scenarios: (1) a rangeland-only scenario (RO) in which animals exclusively graze on rangeland; (2) a rangeland–cropland scenario (RC) in which animals are on rangeland during the cropping season and on cropland during the dry season; and (3) a control scenario (CO) with no cattle presence and very low-intensity background grazing on random days (frequency equal to the mean annual frequency of the RC scenario, animal density equal to 25 % of the mean animal density of the RC scenario), following the scheme described in Pfeiffer et al. (2019), to ensure the development of a grazing-adapted plant community. Based on test runs, we conducted a 310-year spin-up with a randomized climate data sequence from 1980 to 2010 for all scenarios, followed by a 31-year transient simulation from 1980 to 2010. In the RO and RC scenarios, we prescribed grazing between 2000 and 2010 and kept the low-level background grazing from the spin-up for the CO scenario. Natural fire as part of the local rangeland dynamics was allowed during spin-up and transient simulations. Each individual replicate simulation within a scenario was initialized with a unique random seed, creating unique fire occurrence sequences for all simulated hectares. This approach implies small fires that do not fully burn an entire grazing area at a given time and reflects the commonly observed predominance of low-intensity grass layer fires in the region. We used the same set of replicate initializations for all three scenarios, implying that fire event sequences between scenarios were identical up to the start of the grazing treatments but then deviated because the grazing module also uses random numbers and, therefore, alters the sequences between different grazing scenarios. As grazing and fire also interact in the field (e.g., via effects of grazing on fuel availability), we consider this variation as an imitation of the naturally occurring effect.
2.7.2 Rangeland specifics and animal numbers
Based on the herder surveys (see Sect. 2.3), we set livestock to a total of 90 LUs for Gabaza and 87.4 LUs for Selwana. The rangeland area was 1431 ha at Gabaza (15.9 ha per LU) and 2128 ha (24.3 ha per LU) at Selwana. At Gabaza, 71 % of the rangeland was woodland and 29 % was grassland, whereas 55 % was woodland and 45 % was grassland at Selwana (Table 1). While rangeland at Selwana is one contiguous area, Gabaza's rangeland includes four sub-areas (A1 to A4) with area sizes of 57, 279, 394, and 683 ha. Lacking detailed information, we assumed equal woodland–grassland partitioning for all sub-areas.
2.7.3 Spatiotemporal distribution of livestock
The aDGVM2 required daily information on the LUs visiting a given hectare. The aDGVM2 models vegetation on individual 1 ha stands with no information exchange between different hectares. Spatiotemporal sequences mimicking animal movement on grazed areas were created (see the following subsections for details) to determine the animal effects on each individual hectare on any given day, as explicit animal movement was not tracked. At Gabaza, we established an additional temporal subdivision to determine the duration of animal presence on each sub-area. We assumed active herd relocation between sub-areas (and between rangeland and cropland) and prescribed the use of only one sub-area at a time. Additionally, we partitioned the period of animal presence proportionally to the sub-area size. Thus, due to this size-proportional time split between sub-areas, all rangeland hectares experience approximately the same annual demand, independent of their location in a small or large sub-area, but resting periods are longer for hectares located in the smaller sub-areas. Therefore, differences in the feed deficit size between different sub-areas can be attributed to seasonality, as the grazing load is distributed equally between sub-areas. The grazing periods were prescribed as input to the aDGVM2 grazing routine. Figure 2 illustrates presence/absence on rangeland for both villages and scenarios (RO and RC). The proportional reduction in the period of animal presence in the RC scenario compared with the RO scenario is illustrated in the Supplement (Fig. S1).
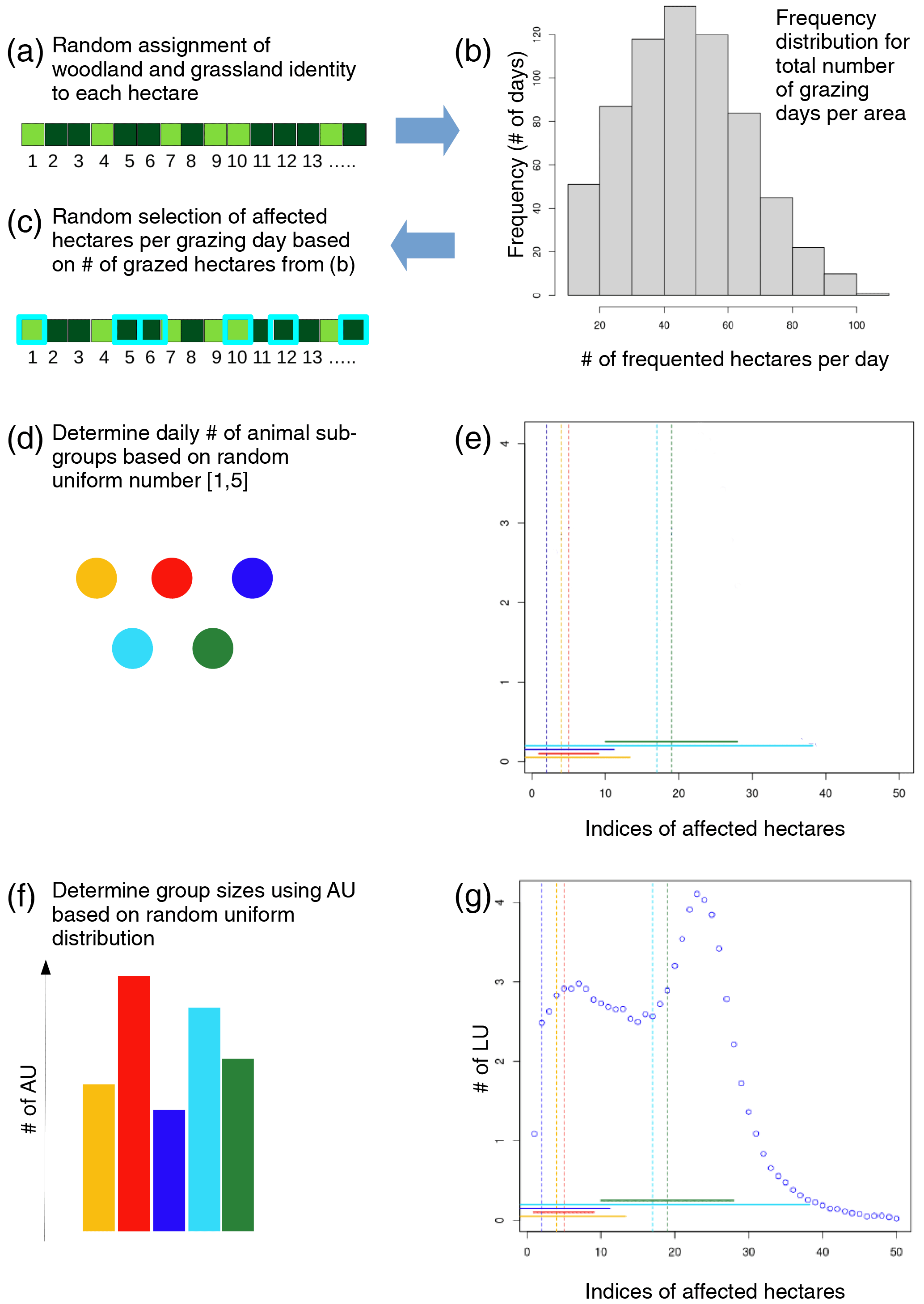
Figure 3Illustration of the steps involved in creating the hectare-specific grazing sequences.
2.7.4 Generation of hectare-specific grazing sequences: the daily choice of affected hectares
We assigned an index to each hectare in a given area and attributed it to woodland or grassland based on the respective percentages of both vegetation types (Fig. 3a). We then determined the grazing-affected hectares on a given day by (1) the number of affected hectares (Naff) (“how many?”) and (2) the indices of affected hectares (“which hectares?”). Naff depends on the herd walking range. Cattle typically move between 1 and 13 km d−1 (Schrader, 2007). To link walking distance with the Naff, we assumed that animals moving between hectares walk a minimum of 100 m (the lateral length of a hectare). Based on the typical range of walking distance combined with the spatial scatter of the herd and lacking more detailed information, we defined a mean daily Naff of 50, with a standard deviation of ± 20 ha, and a minimum daily Naff of 10. If the available grazing area was smaller than Naff, animals grazed all hectares in the area. We drew a random number from a normal distribution using the 50 ha mean and 20 ha standard deviation to determine the daily Naff during periods of animal presence in a given area. If a random number was <10, we replaced it with a random uniform number between 10 and 100 ha to ensure the minimum daily Naff of 10. In this way, we created a frequency distribution for the number of visited hectares per day akin to the one shown in Fig. 3b. Knowing Naff for each day in the 11-year simulation period, we determined the daily indices of affected hectares (Iaff) via daily random uniform subsampling from all available hectare indices of the considered area (Fig. 3c), i.e., by randomly choosing Naff random indices.
2.7.5 Daily animal distribution across affected hectares
To distribute the LUs across affected hectares using the Iaff, we converted LUs to daily animal units (AUs) by multiplying the minutes per day by the number of LUs (i.e., 1440×90 LUs = 129 000 AUs for Gabaza, and 1440×87.4 LUs = 125 856 AUs for Selwana). This minute resolution considers that animals may spend only part of a day on an individual hectare. For each day, we randomly created between one and five animal subgroups () with varying sizes and spatial densities (Fig. 3d). We placed the group centers randomly on the Naff, as shown in Fig. 3e using an example with five sub-groups. For each group, we drew the standard deviation around the group center from a random uniform distribution ranging between 0.5 and 1 times the Naff (the colored horizontal lines in Fig. 3e illustrate the standard deviation, and the vertical lines represent the locations of the individual group centers). We chose this spread around the group center somewhat arbitrarily to describe how tightly the animals within a group stay together. The scatter also influences how strongly subgroups intermingle on the number of affected hectares. We assigned the AUs to the different groups by creating fractional group sizes by first drawing (Ng−1) random uniform numbers between 0 and 1 and then sorting these by size to establish the group breaks. Fractional group sizes were calculated as the differences between the breaks (including 0 and 1 as lower and upper edges) and were multiplied by the number of AUs to obtain the daily AU per group (Fig. 3f). For each AU group, we applied a random normal distribution using the group's mean and standard deviation (the group center and scatter, as shown in Fig. 3e) to determine how many AUs are assigned to the hectares around the group mean. We iteratively redistributed AUs whose drawn numbers were <1 or >Naff.
Generating individual distributions for varying numbers of animal groups of changing size allows flexible animal distribution across the affected hectares. The superordinate animal distribution of the herd emerges from superimposing the individual normal distributions (Fig. 3g) and allows the creation of flexible multi-modal livestock distributions. It imitates the daily livestock group dynamics in a pseudo-explicit manner that does not require exact knowledge of spatial relationships between affected hectares while still creating average long-term characteristics of herd behavior and rangeland usage. When assigning the AUs per hectare to the affected hectares, we sorted the AUs per hectare in descending order and randomly assigned the lower range of the ordered list to the affected woodland hectares to account for the higher feed availability on grassland compared with woodland. For use in aDGVM2, we reconverted the daily assigned AUs for each affected hectare to daily LUs per hectare (Fig. 3g).
2.7.6 Simulation of representative hectares
Although we created grazing sequences for all hectares (see the Supplement), computational constraints precluded simulating the total rangeland. Therefore, we simulated different grazing intensity levels based on the total LU visits per hectare during the experiment. For each rangeland (sub-)area, we conducted 150 simulations (75 ha of grassland and 75 ha of woodland), with 15 simulations per area and vegetation type ranging around the minimum, maximum, median, 25th percentile, and 75th percentile of the cumulative LU numbers per hectare over 11 years, respectively. For sub-area A1 at Gabaza, 57 ha (i.e., the complete sub-area) was simulated. For simulated percentages of (sub-)areas, see Fig. 1a. We conducted 657 simulations for each scenario (CO, RO, and RC), i.e., 1971 simulations in total.
2.7.7 Feed gap analysis
Consumable grass biomass includes living leaves and stalks as well as standing dead and reproductive biomass. A minimum amount of each pool is unavailable to grazers as they cannot graze completely down to the ground. For cattle, we defined a limit of 30 g m−2 of living and dead biomass, respectively, that needs to remain as well as 10 g m−2 of seed biomass (see Pfeiffer et al., 2019). Feed gaps occur when demand exceeds the available biomass. We calculated the average annual demand, consumption, and deficit across all simulated hectares by vegetation type (i.e., for woodland and grassland hectares) for both villages and for (sub-)areas. In addition, we summed the annual deficits of all simulated hectares per village and divided this sum by the summed demand from all simulated hectares. This ratio characterizes the demand-specific deficit severity and allows comparisons between different years. In the same way, we calculated the ratio between the number of days with a grazing deficit and the sum of rangeland grazing days. As animals in the RC scenario only spent part of the year on the rangeland, we scaled both ratios by the fraction of the year that animals were present on the rangeland for this scenario. We also created hectare-scale overviews for biomass consumption and demand to assess variability between hectares, which allowed for comparisons between both villages as well as sub-areas at Gabaza.
2.8 Performance indicators
For the evaluation of the cropland simulations, we analyzed the temporal grain and straw yield (dry matter) dynamics at both villages and determined site-specific differences in yield patterns and between SQ and SI scenarios. Aside from yields, we analyzed the SOC status, cropland vegetation cover, and soil water dynamics. For the rangeland simulations, we considered metrics that describe animal-related aspects and grazing pressure on vegetation, such as grazing frequency and biomass demand relative to consumption. We identified when feed gaps occur and their severity compared with demand. Additionally, we examined the seasonal and interannual dynamics of grass biomass and productivity (net primary productivity, NPP, and gross primary productivity, GPP) to determine the between-village difference between the RO and RC scenarios.
3.1 Cropland simulation results
3.1.1 Yield comparison between the two villages
Maize grain yield
Village-scale grain yields showed considerable interannual variability (Fig. 4a, b; see Fig. S2 for an individual depiction per crop type). With a larger arable area, Selwana had higher total yields than Gabaza but lower yields per hectare due to the drier conditions (Fig. 4c, d). The total yield at Gabaza in the SQ scenario varied between 396 t in 2005 and 1253 t in 2006 (mean of 910 ± 251 t). Yields per hectare ranged between 1.2 t ha−1 and 3.7 t ha−1 (mean of 2.7 ± 0.8 t ha−1). SI led to a moderate, non-significant, factor of 1.2 ± 0.1 average yield increase (Fig. 4f). Straw yield was moderately, but at Selwana not significantly, increased by SI (Fig. 4e). Maize production at Selwana varied greatly, from 367 t in 2001 to 5739 t in 2004 (mean of 2620 ± 1603 t). Maize grain yields per hectare at Selwana were 53 ± 23 % lower than at Gabaza, ranging between 0.2 and 2.9 t ha−1 (mean of 1.3 ± 0.8 t ha−1). Compared with Gabaza, SI at Selwana led to a slightly higher mean maize yield increase (by a factor of 1.2 ± 0.3, although statistically non-significant), but the response varied considerably between years (Fig. 4f). The highest yield increase at Selwana (factor of 1.8 increase) occurred in 2005. However, SI reduced maize yield by 11 % compared with the SQ scenario in 2004 and to a lesser extent also in 2002 and 2006. At Gabaza, SI consistently increased maize yields.
Cowpea grain yield
The mean cowpea yield in the SQ scenario at Gabaza was 62 ± 24 t, with a minimum of 30 t in 2004 and a maximum of 96 t in 2007, and a mean yield per hectare of 0.7 ± 0.4 t ha−1 (Fig. 4). In the SI scenario, grain yield increased by a factor of 1.6 ± 0.0 on average (statistically significant increase at p<0.05; Fig. 4f). The total mean village cowpea grain yield in the SQ scenario at Selwana was 169 ± 92 t, with a minimum of 36 t in 2005 and a maximum of 328 t in 2008. By hectare, the mean yield was 0.4 ± 0.2 t ha−1 (minimum of 0.1 t ha−1 and maximum of 0.7 t ha−1). SI led to a mean, non-significant, factor of 1.3 ± 0.2 yield increase.
Peanut grain yield
Gabaza had a village-scale peanut yield of 161 ± 27 t, with a minimum of 125 t in 2005 and a maximum of 214 t in 2000 (Fig. 4b). The mean yield per hectare was 0.9 ± 0.2 t ha−1 (minimum of 0.7 t ha−1 and maximum of 1.2 t ha−1; Fig. 4d). SI increased the yield by an average factor of 1.2 ± 0.0 (range from 1.1 to 1.3, increase significant at p<0.05; Fig. 4f). At Selwana, the village-scale peanut yield was 199 ± 74 t, ranging between 63 t in 2003 and 309 t in 2000 (Fig. 4a). By hectare, the mean yield was 0.5 ± 0.2 t ha−1 (minimum of 0.1 t ha−1 and maximum of 0.7 t ha−1; Fig. 4c). On average, SI increased peanut yields at Selwana by a factor of 1.3 ± 0.3 (statistically non-significant), with the strongest positive effect in 2001 (factor of 1.7 increase; Fig. 4f). However, in 2005, yield at Selwana was reduced by more than 50 % compared with the SQ scenario.
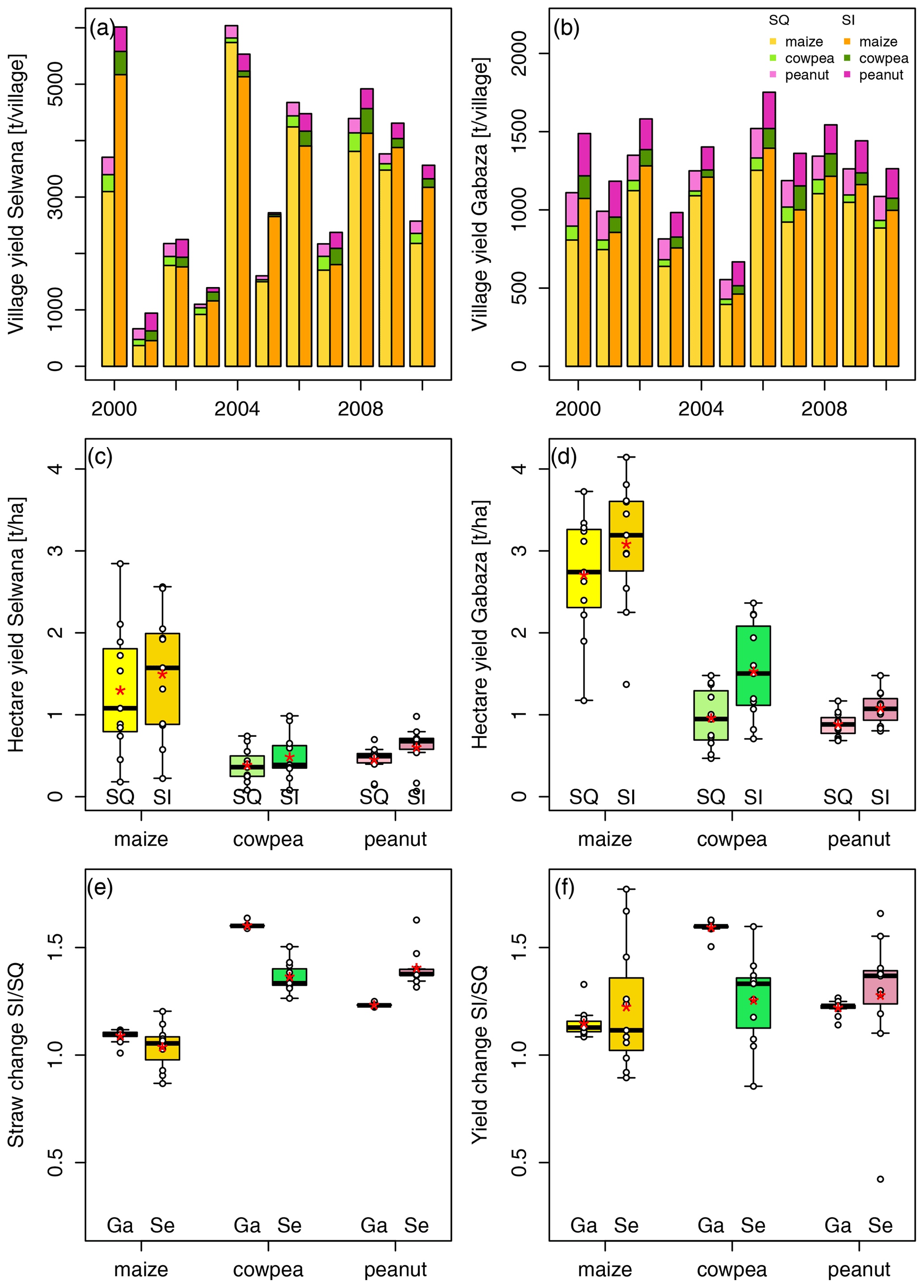
Figure 4Time series of village-scale crop grain yields for Gabaza (a) and Selwana (b) under status quo (SQ) management and sustainable intensification (SI), and hectare-specific grain yields at Gabaza (c) and Selwana (d). Panels (e) and (f) summarize the change in grain yield and straw quantity between the SI and SQ scenarios at Gabaza (Ga) and Selwana (Se) for maize, cowpea, and peanut, respectively. The white dots in panels (c) to (f) depict the values of individual years, and red asterisks represent the mean value.
Effect of SI measures on grain yields – summary comparison
For cowpea, SI had a stronger positive effect at Gabaza with respect to relative and hectare-specific increases. Cowpea benefited the most with a factorial increase of 1.6 ± 0.0 (absolute yield increase of 0.6 ± 0.2 t ha−1). With the highest hectare-specific yields of all crops, the moderate factor of 1.2 ± 0.1 increase led to an overall increase of 0.4 t ha−1 for maize, while the relative factor of 1.2 ± 0.0 increase for peanut corresponded to a yield increase of 0.2 ± 0.1 t ha−1. At Selwana, the hectare-specific yield increase was highest for maize (0.2 ± 0.4 t ha−1; factor of 1.2 ± 0.3 relative increase), followed by peanut (1.2 ± 0.1 t ha−1; factor of 1.3 ± 0.3 relative increase) and cowpea (0.1 ± 0.1 t ha−1; factor of 1.3 ± 0.2 relative increase). The crop-type-specific responses also caused a slight shift in the relative contribution of each crop type to the total village-level yield (see Fig. S3).
SI led to a statistically significant increase in village-scale yield at Gabaza for cowpea and peanut, although not for maize (Welch two-sample t test, p<0.05). No crop type at Selwana showed a significant increase. Moreover, the interannual variability in SI-related yield effects was more pronounced and even negative in some years.
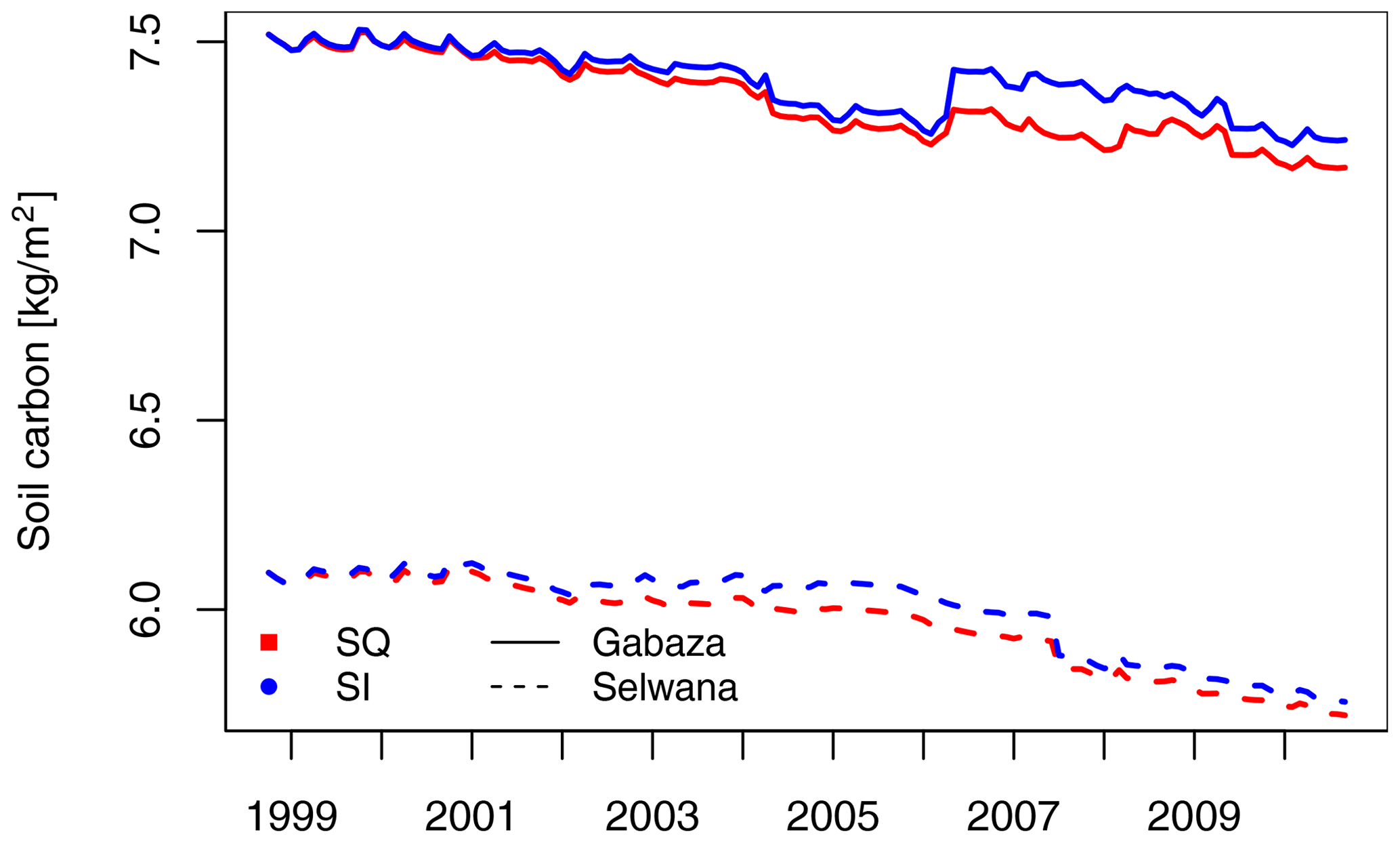
Figure 5Time series of the cropland soil organic matter (SOC) content at Gabaza and Selwana.

Figure 6Monthly cropland leaf area index (LAI) at Selwana (a) and Gabaza (b) as well as monthly extractable soil water at Selwana (c) and Gabaza (d). The abbreviations used in the figure are as follows: SQ – status quo management, SI – minimum sustainable intensification, and n – non-significant difference between SQ and SI. Red asterisks represent mean values.
3.1.2 Soil organic carbon, cropland vegetation cover, and soil water
Cropland soil organic carbon (SOC) gradually decreased irrespective of management (Fig. 5), although somewhat less under SI. At Gabaza, SOC declined by 4.7 %, from 7.5 to 7.2 kg C m−2, in the SQ scenario and by 3.7 % in the SI scenario (7.3 kg C m−2 left in 2010). On average, cropland soil at Selwana stored 1.4 ± 0.1 kg C m−2 less than at Gabaza, and C loss was 6.2 % in the SQ scenario (from 6.1 kg C m−2 in 1998 to 5.7 kg C m−2 in 2010). SI only had a minor effect on SOC loss at Selwana (5.6 % loss, 5.8 kg C m−2 in 2010). At the village scale, cropland soils under SI had 427 t (Gabaza) and 1030 t (Selwana) more carbon stored in 2010 than under SQ management conditions.
The mean green leaf area index (LAI) for cropland varied both seasonally and interannually (Fig. 6). Monthly values at Selwana (Fig. 6a) were generally lower than at Gabaza (Fig. 6b). SI had a moderately positive but statistically non-significant effect on the LAI, and the highest canopy-cover values occurred during the growing period from December to March. After crop harvest, weed infestation drove the cropland LAI during the dry season.
Extractable soil water (ESW) was generally higher at Gabaza (Fig. 6c, d). Minimum values occurred in August and September and increased throughout the wet season before gradually declining again. At both locations, SI tended to reduce ESW due to the higher vegetation cover and biomass simulated in the scenario. However, the difference between the SQ and SI scenarios was non-significant.
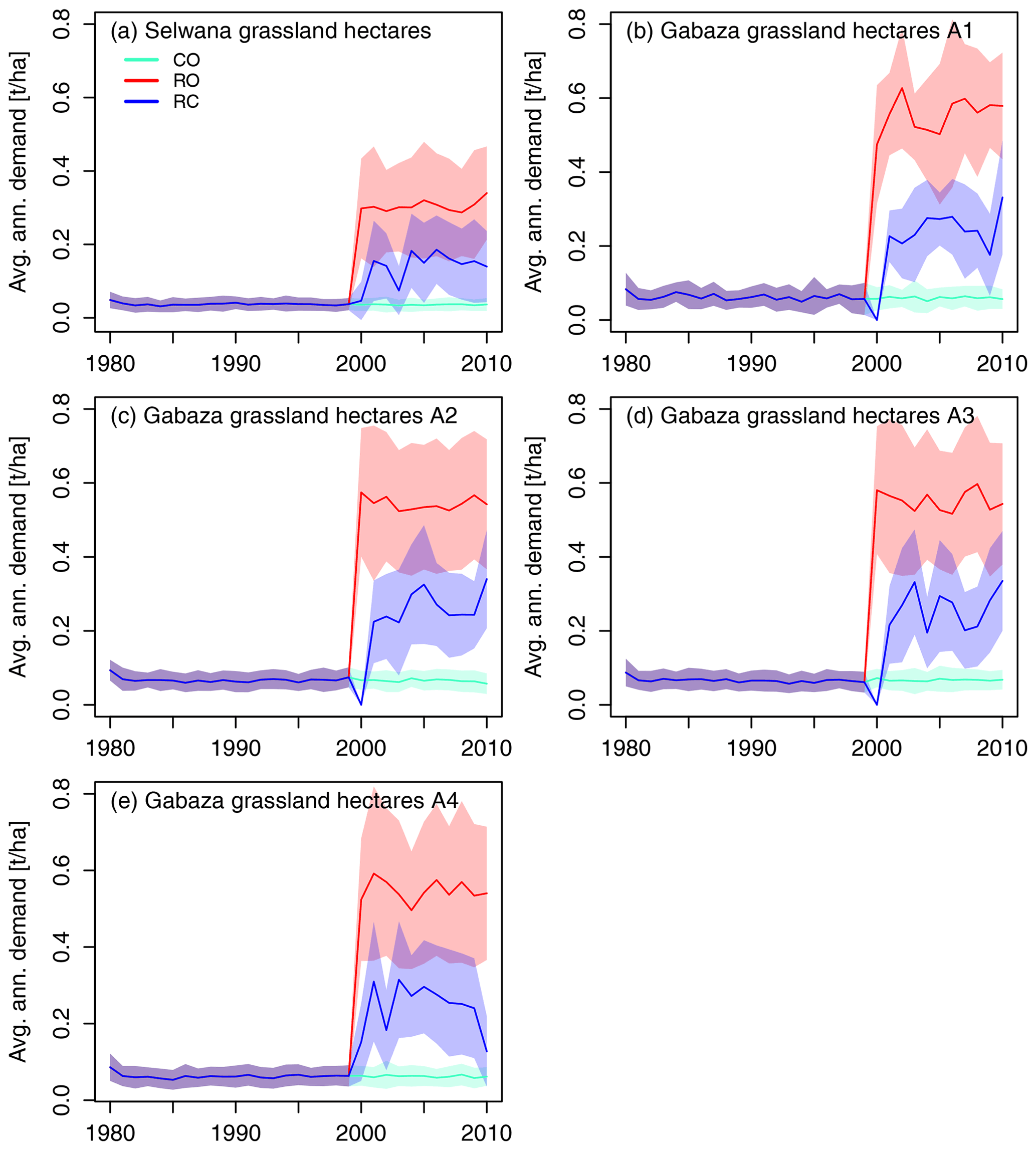
Figure 7The mean per-hectare annual biomass demand across simulated grassland hectares at Selwana (a) and the four sub-areas at Gabaza (b–e). Non-zero biomass demand in the CO scenario is due to the prescribed low-intensity background grazing caused by small game when cattle are completely excluded.
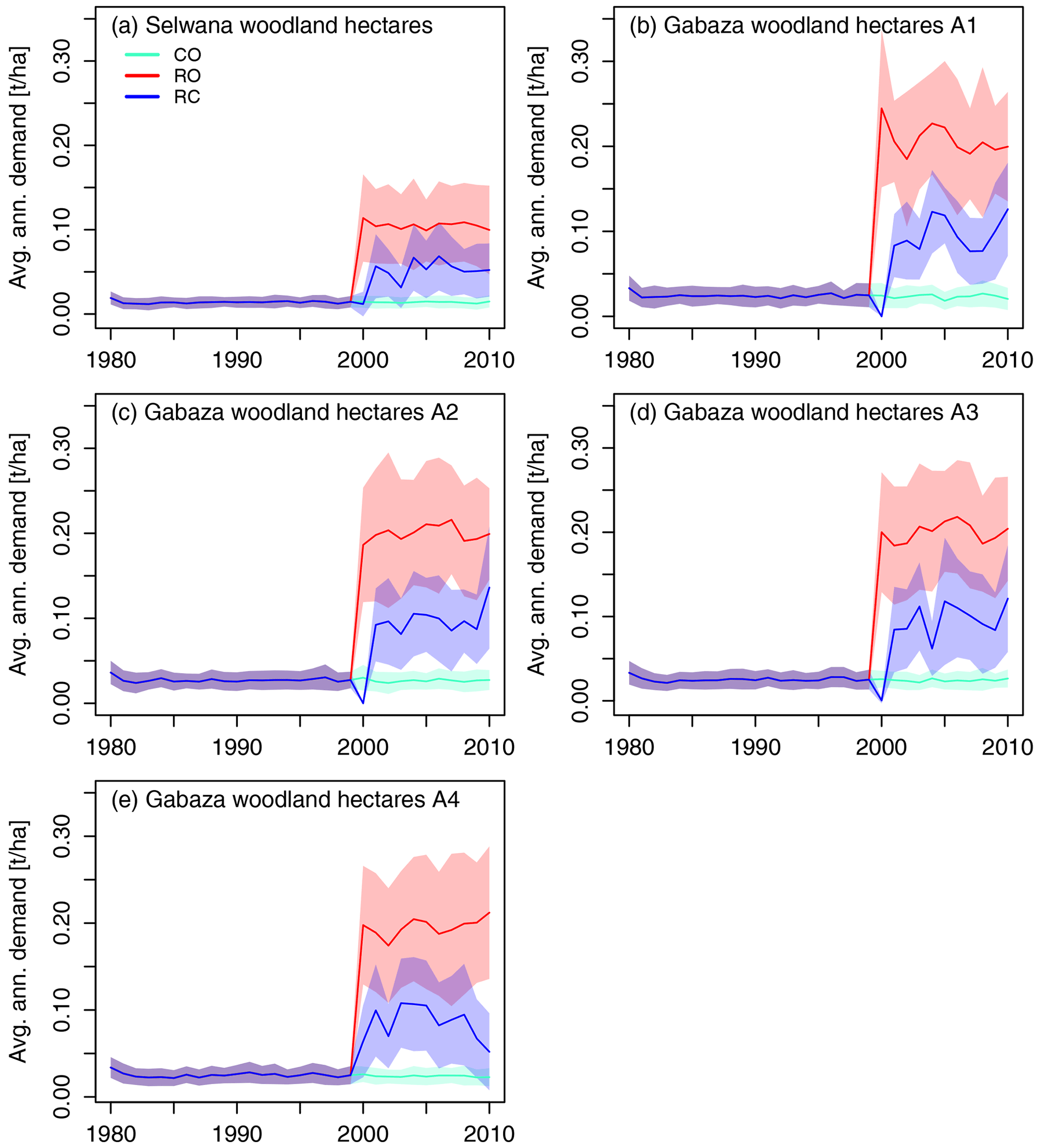
Figure 8The mean per-hectare annual biomass demand across simulated woodland hectares at Selwana (a) and the four sub-areas at Gabaza (b–e). Non-zero biomass demand in the CO scenario is due to the prescribed low-intensity background grazing caused by small game when cattle are completely excluded.
3.2 Rangeland simulation results
3.2.1 Biomass demand, grazing frequency, and consumption
The hectare-specific mean annual grass biomass demand was lower at Selwana (Figs. 7, 8; for detailed numbers, see Table S2a). There, animals required approximately 300 kg ha−1 yr−1 on average from grassland hectares in the RO scenario but only between 70 and 190 kg ha−1 yr−1 in the RC scenario. The biomass demand on woodland hectares was lower in both scenarios due to animals preferentially visiting grassland hectares with higher biomass availability. Due to the higher stocking density, the annual hectare-specific demand at Gabaza was consistently higher than at Selwana (see panels b–e in Figs. 7 and 8), roughly ranging between 470 and 630 kg ha−1 yr−1 in the RO scenario and between 130 and 340 kg ha−1 yr−1 in the RC scenario on grassland hectares. Due to the fact that the presence period scaling is proportional to the size of the sub-areas at Gabaza, the hectare-specific demand is approximately equal for all sub-areas. Interannual variability in average demand results from the variability in the visit pattern generated according to the description in Sect. 2.7 and the fact that only a sample of the total number of rangeland hectares was simulated at each site.
On average, grazers visited each hectare at Selwana on 8–9 d yr−1 in the RO scenario (Table S2b) and on to 2–6 d yr−1 in the RC scenario. On average, at Gabaza, LUs frequented each hectare on 11–14 d yr−1 in the RO scenario and on 3–8 d yr−1 in the RC scenario, except for the smallest sub-area A1 where the average visit frequency was between 7 and 11 d yr−1 in the RO scenario (annual presence on A1 was 15 d yr−1) and between 3 and 6 d yr−1 in the RC scenario (presence on A1 was 3–8 d yr−1, depending on the year).
Despite a similar visit frequency, the average grass biomass consumption on woodland was lower than on grassland (Table S2c) due to the lower number of animals assigned to woodland hectares (see Sect. 2.7.5). The mean daily grass biomass consumption on affected hectares was comparable between the RO scenario and RC scenario, as the same number of animals per area were present on a per-day basis during both scenarios. Differences in annual grass biomass consumption between scenarios only emerged due to the shortened presence periods per area in the RC scenario (Table S2d).
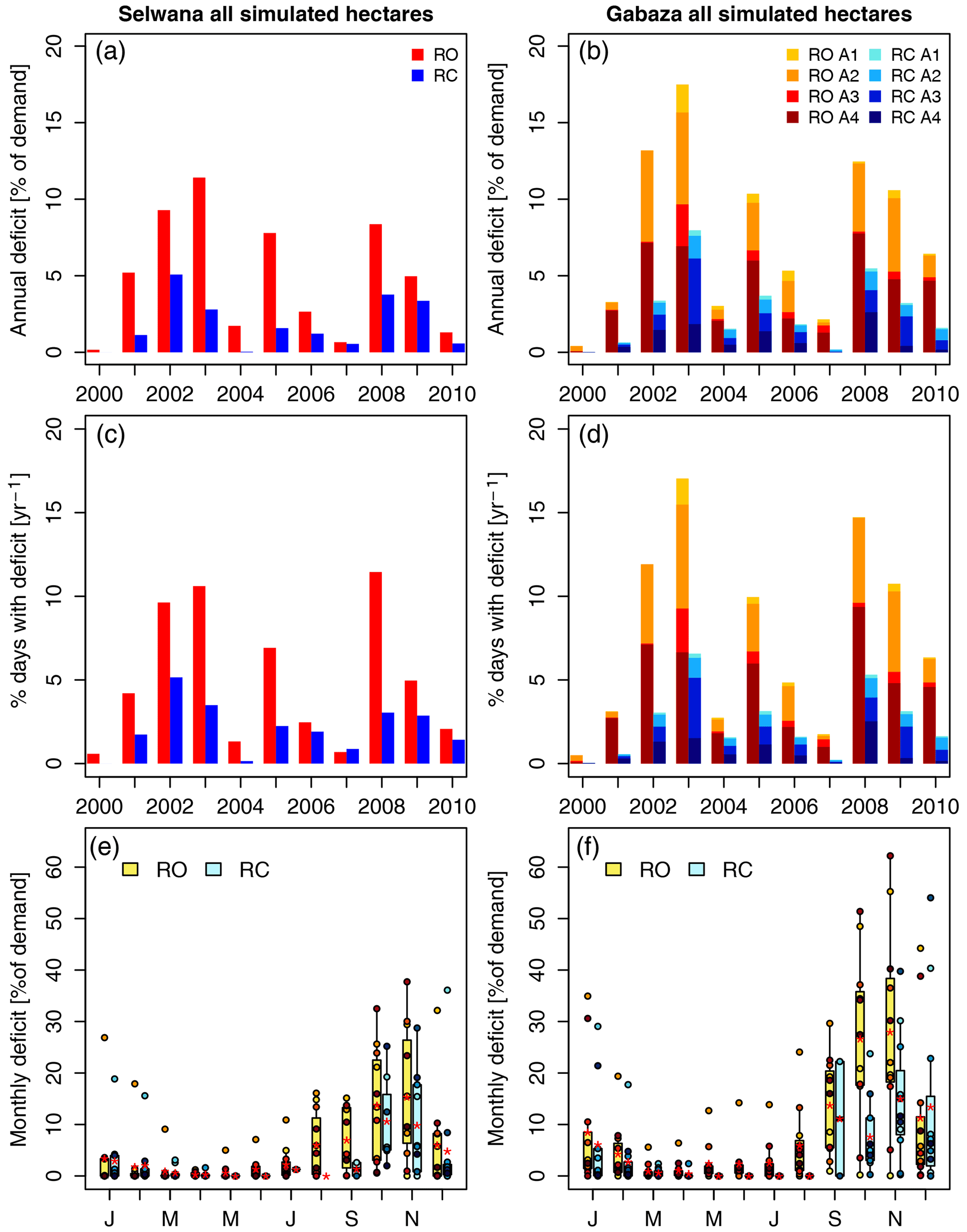
Figure 9The demand-specific annual deficit across all simulated hectares, and the percentage of grazing days integrated across all simulated hectares that had a deficit (irrespective of the size of the deficit), relative to the total grazing days within a year. Panel (a) shows the relative deficits for Selwana, and panel (b) presents the relative deficits for Gabaza. Panel (c) shows the percentage of grazing days with a deficit for Selwana, and panel (d) displays the percentage of grazing days with a deficit for Gabaza. The subdivisions of bars in panels (b) and (d) indicate the relative contribution of each sub-area to the site-scale annual deficit and days with deficit, respectively. Panels (e) and (f) show the monthly demand-specific deficits across all simulated hectares per village for Gabaza and Selwana, respectively. Red asterisks denote mean values, and white dots denote annual values.
3.2.2 Feed gaps on rangeland
Across all simulated hectares, the annual demand-specific deficit (gap between supply and demand) at Selwana ranged from 0.2 % to 11.4 % in the RO scenario (Fig. 9a). In the RC scenario, the deficit declined to values between 0 % and 5.1 %, partially due to the shorter herd presence on rangeland. Gabaza, where the average cattle density per hectare was 1.5 times higher, had higher relative deficits. Moreover, Gabaza has a higher relative woodland proportion (Table 1). Demand-specific deficits at Gabaza were between 0.4 % and 17.5 % in the RO scenario (Fig. 9b) and declined to values between 0 % and 8.0 % in the RC scenario. Years with higher deficits, such as 2002, 2003, 2005, and 2008, coincide with low annual precipitation at both sites (Fig. 1b). The maximum deficits occurred in 2003, which was the second dry year in a row after 2002. For the respective relative deficits for grassland and woodland hectares, see Fig. S4.
Integrated across all simulated hectares, the time line of the percentage of grazing days with a deficit resembles that of the relative deficits (compare panels c and a and panels d and b in Fig. 9). Between 0.6 % and 11.5 % of grazing days had a deficit in the RO scenario at Selwana. The RC scenario reduced this to a range from 0.0 % to 5.2 % of total days. At Gabaza, the higher grazing pressure caused more deficit days. Here, the RO scenario resulted in a range from 0.5 % to 17.0 % of grazing days, with a reduction to 0.0 %–6.6 % in the RC scenario. For a presentation by vegetation type, see Fig. S5. Deficit timing showed a clear seasonal pattern (Fig. 9e, f). Monthly deficits in the RO scenario were lowest between March and June and highest in October/November before declining towards December. While both villages had a similar seasonal pattern, the demand-specific monthly deficits were higher at Gabaza, particularly from September to November. Cropland pasturing between approximately April to October (see Fig. 1e, f) avoided deficits during these months. However, it also reduced the deficit size during rangeland presence (compare the RO and RC scenarios in Fig. 9e, f). Deficits showed a delay of 2–3 months relative to precipitation (Fig. 1c, d), i.e., the maximum was delayed and occurred after the end of the dry season, when precipitation increased again. Similarly, the lowest deficits in March/April occurred approximately 2 months after peak precipitation.

Figure 10Temporal dynamics of average consumable grass biomass on simulated grassland hectares (living and dead standing grass leaf biomass, and reproductive biomass, reduced by the minimum amount that is not available to grazers, i.e., 0.3 t h−1 for living and dead grass biomass, respectively, and 0.1 t ha−1 of reproductive biomass). Lines denote the mean across all simulated hectares, and shaded areas show the standard deviation. The horizontal lines at the bottom of the panels denote the respective animal presence times for the RO and RC scenarios.
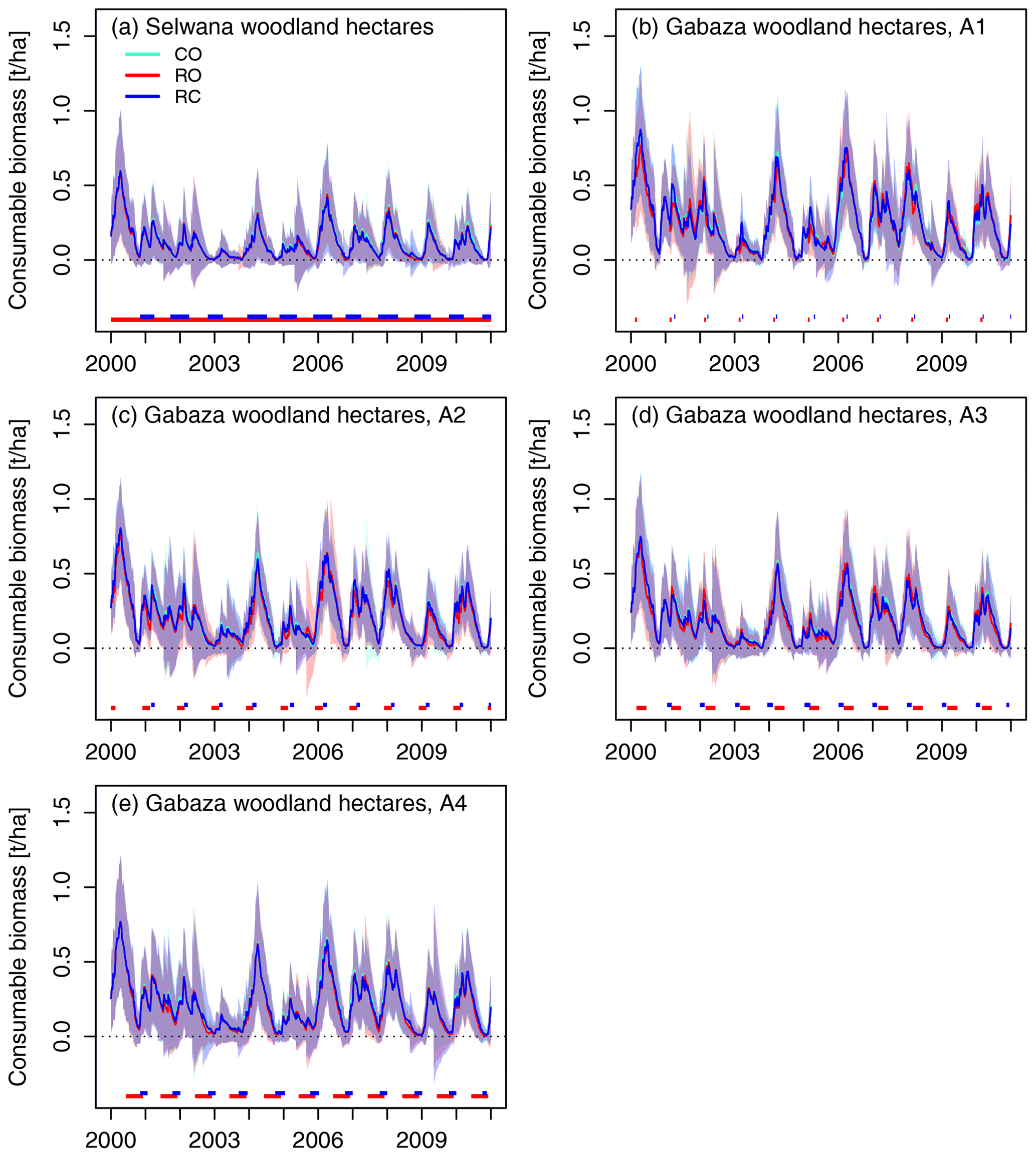
Figure 11Temporal dynamics of average consumable grass biomass on simulated woodland hectares (living and dead standing grass leaf biomass, and reproductive biomass, reduced by the minimum amount that is not available to grazers, i.e., 0.3 t h−1 for living and dead grass biomass, respectively, and 0.1 t ha−1 of reproductive biomass). Lines denote the mean across all simulated hectares, and shaded areas show the standard deviation. The horizontal lines at the bottom of the panels denote the respective animal presence times for the RO and RC scenarios.
3.2.3 Temporal dynamics of grass biomass
Consumable grass biomass provision was distinctively seasonal (Figs. 10, 11). Peak biomass occurred between February and March, and the minimum availability developed towards the end of the dry season in October/November. Overall biomass availability was lower at Selwana than at Gabaza, and woodland hectares produced less consumable biomass than grassland hectares. Consumable biomass also greatly varied between years. As the second of 2 consecutive dry years, 2003 had the lowest biomass of the 11 years. Grazing had a minor effect on the across-hectare mean consumable biomass (compare the scenario averages and standard deviations in Figs. 10 and 11). Differences in fire occurrence between grazing scenarios also caused biomass differences (see Figs. S6 and S7).
3.2.4 Rangeland productivity (NPP and GPP)
For Selwana grassland, the average annual GPP in the control ranged between 3.1 ± 1.5 and 11.2 ± 4.1 t ha−1, whereas Gabaza had values between 3.7 ± 1.9 and 15.8 ± 6.5 t ha−1 (Table S3, Fig. S8). On woodland, the hectare-specific grass-layer GPP was lower and ranged between 1.2 ± 0.6 and 3.6 ± 1.7 t ha−1 at Selwana and between 1.3 ± 0.7 and 4.7 ± 1.6 t ha−1 at Gabaza (Table S3, Fig. S9). Differences between scenarios were statistically non-significant (two-sided t test, p<0.05).
Annual grassland NPP at Selwana varied considerably and ranged between 1.2 ± 0.5 and 6.0 ± 2.3 t ha−1 in the control scenario. Annual grassland NPP at Gabaza was higher, with values between 1.4 ± 0.6 and 8.7 ± 3.8 t ha−1 (Table S4, Fig. S10). We also simulated this pattern for woodland hectares, where the mean annual NPP ranged between 0.4 ± 0.3 and 1.9 ± 0.9 t ha−1 at Selwana and between 0.5 ± 0.3 and 3.0 ± 0.9 t ha−1 at Gabaza (Table S4, Fig. S11). The RO and RC scenarios did not differ significantly (two-sided t test, p<0.05) from the control, except for the years 2005 and 2007 in the RC scenario for Selwana grassland hectares and the year 2008 in the RO scenario for the woodland hectares of sub-area A1 at Gabaza.
The GPP normalized for living leaf biomass (LLBM) had annual values of approximately between 7 and 13 g C g−1 (see Table S5 and Figs. S12 and S13), and the NPP normalized for LLBM had values of between 2 and 8 g C g−1 (see Table S6 and Figs. S14 and S15). Values of biomass-specific GPP and NPP were comparable between grassland and woodland, i.e., grass on woodland was as productive as grass on grassland. In contrast to the hectare-specific GPP and NPP values, grazing frequently caused significantly (two-sided t test with p<0.05) higher average biomass-normalized GPP and NPP values relative to the control (see Figs. S12, S13, S14, and S15). This effect was usually stronger in the RO scenario. Based on the sensitivity study in Pfeiffer et al. (2019), we suspect that an optimum grazing frequency and intensity likely exists where simulated biomass-normalized productivity is maximized, but we did not explicitly test for this optimum in either the cited or present study. Plot-level GPP and NPP showed pronounced seasonality, and monthly values varied strongly between years (Figs. S16, S17, S18, and S19).
We saw similar seasonality for biomass-specific monthly GPP and NPP (Figs. S20, S21, S22, S23) with minimum values in June and a gradual increase to the December maximum. Values gradually declined from January to March and then rapidly declined towards June. Integrated across all simulated hectares, annual biomass consumption relative to NPP varied between years and was lower at Selwana (Fig. 12). The highest ratios occurred in the dry years 2002, 2005, and 2008 (Figs. S10, S11). The annual consumption / NPP ratio was approximately halved in the RC scenario compared with the RO scenario (reduction by a factor of 2.1 ± 0.6 at Selwana and by a factor of 2.1 ± 0.2 at Gabaza).
The UN Sustainable Development Goals (SDGs; UN General Assembly, 2015) include alleviating poverty (SDG1), reducing hunger and enhancing food production (SDG2), and reducing habitat loss and degradation to preserve terrestrial ecosystems and biodiversity (SDG13). These are among the most desirable goals for rural areas in southern Africa. Agricultural scientists emphasize SI as a way of improving cropland productivity (e.g., Mueller et al., 2012; Cassman and Grassini, 2020) while reducing environmental impacts and maintaining ESSs and ESFs (Tilman et al., 2011; Tscharntke et al., 2012). Given that smallholder farming provides more than 80 % of the food supply in sub-Saharan Africa and Asia (Walpole et al., 2013), SI measure adaptation in smallholder farming systems needs a special focus and an integrated system approach with a range of possible management interventions (Vanlauwe and Dobermann, 2020). With more than 500 million smallholder farms worldwide sustaining the livelihoods of more than 2 billion people (Walpole et al., 2013), smallholder farming needs explicit consideration when discussing SI measures. Landscape-based integrative adaptations linking agricultural and natural systems are required to ensure the continued provision of ESSs and to help smallholders adapt to climate change (Harvey et al., 2013; Vignola et al., 2015). In our study, we present an example that illustrates how linked crop and rangeland modeling can address research questions on the sustainable management of smallholder farming systems at the landscape level. We focused on research questions revolving around (i) the effect of minimum levels of intensification on crop production and (ii) the trade-offs and opportunities of the combined vs. separated management of cropland and rangeland for cattle production.
4.1 Impact of minimum SI measures on cropland ESSs
The minimum intensification measures implemented in our simulations had minor to moderate effects on cropland ESSs. Carbon sequestration increased moderately due to higher yields and SOC input. While SI measures increased SOC sequestration compared with the SQ scenario at the end of the 11-year simulation period, these measures could not reverse the decreasing trend, and cropland soils remained a carbon source, indicating the overly low current input of organic material to the soil. Due to the rough parameterization of soil SOC in APSIM, where C contents are halved with every additional 30 cm of soil depth, SOC losses (in kg m−2) over time may be approximative but should, nonetheless, capture the relative trends and differences between scenarios. Although SI had a negative effect on the amount of plant-available soil water, the interannual variability was larger than the difference caused by SI.
We expected a stronger relative improvement through SI at Selwana, as SQ produced lower yields per hectare at this location compared with Gabaza; therefore, the potential for improvement seemed particularly promising. However, a general prediction of the SI effect strength turned out to be challenging due to the variety of influencing factors that may enforce, although in most cases counter, SI effects. In particular, more pronounced water availability constraints at Selwana under SI caused great interannual response variability. Moreover, the crop-type-specific response intensity was not consistent between sites. With an average significant increase of 59 %, cowpea yields at Gabaza reacted most strongly to SI. Weeding, fertilizer, and manure input moderately improved productivity when soil water was not limiting, with the potential for higher increases with yet more nitrogen and additional phosphorus input (as the latter is often a major limiting factor in the province’s smallholder fields). While N limitation may be less critical for N-fixing legumes such as peanut and cowpea than for maize, both crops nonetheless responded positively to N input, showing that the reduced energy expense required for N fixation also improved legume growth. However, at Selwana, SI measures could result in reduced yields in some years compared with SQ due to enhanced crop growth causing more severe water deficits during crucial times of the growing season. Improved nutrient provision under SI boosts early-stage crop development and results in increased biomass accumulation compared with SQ without fertilizer input. During later stages, the augmented biomass requires more soil water, which can reduce yields and even cause crop failure in years during which water becomes limited during later crop development stages. Without irrigation, crops cannot benefit from the additional nutrients from soil amendments under water limitation. Targeted deficit irrigation could make SI measures more effective but is often unavailable to smallholders. From our survey work, we found that 2 out of 140 smallholders irrigated. However, some initiatives show promise in making irrigation feasible for smallholder farmers – for example, through public investments in South Africa, public–private partnerships in Zimbabwe, and SI business models in Tanzania (Hanjra and Williams, 2020). Whether irrigation is feasible also depends on regional water availability. Recent investigations in our project area (e.g., Lam et al., 2022) have shown a decline in available surface and groundwater resources in some of the catchments. However, very restricted deficit irritation is still often possible if water is collected from rainwater harvesting and if irrigation is realized as drip irrigation from available boreholes and surface water without exhausting water resources, as reported in Magomeyi et al. (2018) and Parry et al. (2020). According to Parry et al. (2020), a combination of training and experiential learning of farmers in the context of irrigation can lead to significant change, including water use reduction, improved nutrient retention, and greater yields.
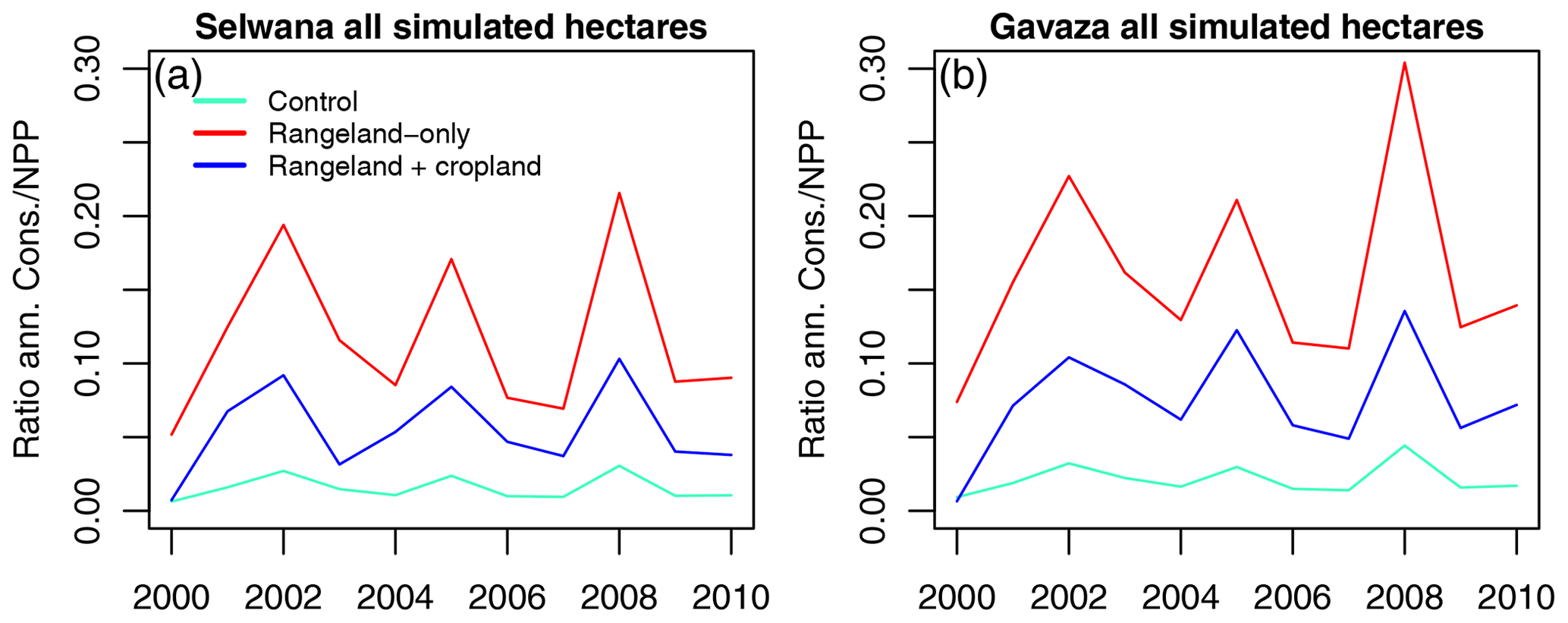
Figure 12Temporal dynamics of the consumption NPP ratio, integrated across all simulated hectares per site.
Plant water availability also depends on the soil water holding capacity (SWC), which is determined by soil texture, SOC, soil flora and fauna, and soil structure. Particularly on sandy soils, organic matter substantially improves the SWC. In addition, loosening compacted soils is an effective way to increase infiltration, create a favorable structure, and increase the SWC. The sandy clay loam at Gabaza and the sandy loam at Selwana may specifically benefit from loosening where the clay and silt components make the substrate prone to slake, hard-setting, and surface crusting, which can cause poor water and air filtration and increase the erosion risk. Although farmers have little influence on soil texture, they can improve soil structure with considerable SWC-enhancing effects (Suzuki et al., 2007). Another measure to amend soil water infiltration and storage is to increase soil organic matter (SOM) by adding plant or animal material, which in turn also reduces soil erosion (Mohler and Johnson, 2009). Moreover, utilizing certain aspects of conservation agriculture (CA) practices, such as diversified crop rotations and crop residue retention can also enhance SOM development, crop yields, and climate resilience (Franzluebbers, 2002; Lehman et al., 2017; Williams et al., 2018; Hoffmann et al., 2020). Participatory research with farmers could help identify realistic pathways for SOM-enhancing interventions to be included in future simulations that explore SI for smallholder systems.
4.2 Feed gaps at village-level and village-specific differences
Feed gaps occurred in both villages. At Gabaza, a higher MAP yielded higher productivity and biomass availability for grazers than the arid rangeland at Selwana. However, higher cattle density at Gabaza compared with Selwana implied higher grazing pressure, which counteracted more favorable environmental conditions and caused higher average deficits at Gabaza.
On average, a given rangeland hectare was grazed on very few days a year (Table S2b). This implies that there was ample time for biomass recovery. Additionally, average hectare-specific consumption was low compared with productivity and had no severe impact on biomass, GPP, or NPP (Figs. 10, 11, S8, S9, S10, S11). These indicators do not hint at a general overgrazing problem, where we would expect a drastic reduction in biomass and rangeland productivity. Moderate rangeland grazing stimulated biomass-specific grass NPP by triggering biomass regrowth and growth overcompensation. Grazing is also beneficial because it removes dead biomass, reduces the LAI of living grasses and self-shading within the grass layer, and increases leaf-area-specific productivity. Regrowth can also indirectly increase leaf-specific productivity because young leaf tissue tends to be high in nitrogen and photosynthetically more active than old leaves (Kitajima et al., 1997; Mediavilla and Escudero, 2003). However, aDGVM2 does not currently capture this latter effect. The effect of grazing intensity and frequency on pasture regeneration was also not explicitly investigated in this study, but it is subject of a separate study on the effects of drought and grazing that is currently in preparation (Behn et al., unpublished).
Due to the lack of explicit spatial movement patterns and information on preferred cattle resting and grazing places, we likely underestimated imbalances in rangeland usage, i.e., we may not have captured overgrazing on specific areas (e.g., next to resting or watering places). Here, a long-term monitoring of cattle movement using GPS collars (Bailey et al., 2018) could reveal such patterns and allow for their incorporation in rangeland simulations.
Substantial feed gaps despite little biomass and productivity reductions seem contradictory. Feed shortage timing explains how deficits occur despite moderate stocking densities. Agreeing with herder statements, we simulated the largest shortfalls at the dry-to-wet season transition (Fig. 9e, f). This also agrees with the findings of Lamega et al. (2021) that mixed crop–livestock farmers in the drier parts of Limpopo perceive spring as the time with the most pronounced feed gaps. In most years, the biomass shortage started in August and intensified until October/November before gradually declining. The middle to late wet season and early dry season usually experienced minimal deficits. Therefore, feed shortages occurred with 2–3 month delays relative to precipitation seasonality (i.e., they were most prominent when precipitation started increasing and were lowest in March/April, approximately 2 months after peak precipitation). This pattern likely emerges because vegetation growth starts shortly after the onset of the first rains, but peak biomass is reached with a delay of 2–3 months after the start of the wet season. During the time when biomass in-growth happens, grass biomass quantities are (a) not yet sufficient to fully supply the demand, (b) dead biomass has been largely consumed during the preceding dry season, and (c) grazing additionally slows the development of new grass biomass. Therefore, livestock are most prone to experience deficits during this critical time.
4.3 Closing feed gaps with the integrated management of cropland and rangeland
Our results show that dry-season residue grazing on cropland can reduce feed shortages. Dry-season access to cropland often more than halved the annual feed gap and shortened the feed scarcity period. Moreover, dead grass on the rangeland lasted longer into the early wet season, which reduced the feed gap between September and November. Eliminating feed gaps would require cattle to return to the rangeland at a later stage than simulated (i.e., when early regrowth of grasses has terminated). However, a later return would collide with crop planting dates. Thus, for sufficient feed supply during the transition period, we recommend storing part of the crop residues directly after harvest, given the availability of storage space and appropriate storage possibilities. This practice is common in other densely populated areas of Africa from Senegal to Ethiopia. The stored residues can then feed cattle until rangeland provides sufficient biomass. Controlled feeding also allows for the treatment of stover to improve nutrient and crude protein supply in order to increase feed intake and reduce dry-season weight loss in cattle (Smith, 2002). Digestibility, energy value, nutrients, and protein content also vary with crop residue type. Therefore, residue mixing can additionally improve livestock supply. Ideally, livestock farmers should provide licks and ensure gradual adaptation to residues and avoid typical problems associated with crop residue feeding (Hofmeyr, 2020). In this context, well-trained extension personnel who provide such information for farmers is crucial.
4.4 Identification of management strategies for the sustained provision of ESFs and ESSs at the landscape level
Linking cropland and rangeland modeling helped analyze individual management scenarios for both land use types and allowed for the identification of management strategies that maximize benefits at the landscape level. In this study, a comparison of village-level crop residue quantities with the feed gap from rangeland grazing revealed how much of the dry-season feed gap can be closed with residue grazing. However, the consideration of time-dependent aspects also showed that quantities are only a partial aspect that may lead to incomplete conclusions. The timing of the most severe feed deficit at the beginning of the wet season coincides with the start of the crop planting season. Therefore, a complete elimination of feed deficits requires off-field access to previously collected crop residue. Moreover, feed-quality-related aspects may be underestimated in our study, as aDGVM2 only crudely accounts for quality by assigning dead biomass a nutrition value of 66 % compared with living grass biomass, but it does not track seasonal changes in living grass biomass.
Dry-season crop residue grazing can reduce feed gaps and accelerate nutrient turnover via manure dropping. The simulated village-level crop residues provided feed supply and additionally allowed SOM formation on cropland. However, neither the SQ nor the SI scenario had enough carbon input to reverse the decreasing SOC trend on cropland, although SI measures reduced SOC loss rates. While we did not conduct crop growth simulations without cattle residue grazing, the comparably low yields and nutrient input levels suggest that avoiding SOC loss may be challenging even without cattle presence. The implementation of context-specific conservation agriculture measures may improve cropland SOC formation and soil water availability.
Based on our results, we propose the following strategies to ensure the sustained provision of landscape-level ESFs and ESSs: (1) apply the minimal SI measures prescribed in our simulations and considered feasible for smallholders at our study villages to moderately increase yields of the staple crops. To avoid the negative effects of SI measures due to water limitation, the adoption of deficit irrigation would be ideal, combined with water conservation measures, runoff prevention, rainwater harvesting, and soil amendments to increase soil water storage capacity, in particular where irrigation is not possible. (2) To reduce dry-season grazing deficits, we propose giving cattle access to cropland and allowing crop residue grazing. At the beginning of the crop planting season, crop residues collected postharvest can be offered off-field to avoid feed deficits and allow for a quick build-up of new grass biomass by alleviating grazing pressure on the rangeland.
Integrated crop–livestock management of highly diverse smallholder farming systems in semiarid ecosystems is crucial to ensure the continued provision of ESFs and ESSs. Here, the presented linked cropland–rangeland simulations can identify potential pathways towards the SI of crop production and the reduction of livestock feed gaps. We found that modest SI measures – deemed feasible for the smallholder farmers in question – can increase yields and SOC sequestration; however, water limitation during later-stage crop development can counteract SI measures without adequate irrigation or measures to conserve water and increase the soil water holding capacity. We found that dry-season cropland residue grazing can substantially reduce feed deficits. Nevertheless, the most severe rangeland feed gaps occurred at the beginning of the wet season when grass biomass regrowth was at an early stage. Both findings have marked implications at the policy level and call for appropriate actions. If crop residues are abundant, partial postharvest collection, storage, and provision of residues as feed during the dry-to-wet season transition period could reduce grazing pressure during early rangeland vegetation development. Research approaches that capture landscape-level interactions and synergies are crucial as climate change impacts and extreme events become more severe. The future climate resilience of ESFs and ESSs needs a landscape-scale evaluation to identify effective mitigation and adaptation strategies. Targeted experimental work and environmental monitoring allows for the evaluation of model components for new scenarios. Subsequently, an updated integrative modeling framework, as suggested by Rötter et al. (2021), can be applied to test different climate change and management scenarios. Further development should include an animal physiology model to simulate the dynamics of animal growth, reproduction, and health condition based on nutrition status. Furthermore, incorporating agent-based modeling could improve the representation of animal behavior on the rangeland and account for interactive decision processes made by human agents based on both economic and ecological criteria. Likewise, further management options could be included, such as the identification and sowing of suitable “dry-season cover crops” that are beneficial for animal nutrition and soil improvement.
The data analysis scripts used to conduct the data analysis and to create the figures shown in the paper and the Supplement are available at https://doi.org/10.17605/OSF.IO/R2TS7 (Pfeiffer et al., 2022). The aDGVM2 code contains currently unpublished parts not related to this publication and is available from the authors upon request.
The supplement related to this article is available online at: https://doi.org/10.5194/bg-19-3935-2022-supplement.
MP, MPH, RR, and SS conceived the study. MP and MPH conducted the aDGVM2 and APSIM simulations. MP, MPH, and WN conducted the analysis of the simulation results. MP led the writing of this article. MPH, WN, JI, KA, JJO, and RR were involved in and contributed to the field survey campaigns and data collection. All authors contributed to the writing of this article.
The contact author has declared that none of the authors has any competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This paper is dedicated to the memory of our esteemed colleague Marian Koch.
This research has been supported by the Bundesministerium für Bildung und Forschung (South African Limpopo Landscapes Network SPACES II “SALLnet”, grant nos. 01LL1802A and 01LL1802B).
This paper was edited by Paul Stoy and reviewed by Carsten Marohn and one anonymous referee.
Bailey, D. W., Trotter, M. G., Knight, C. W., and Thomas, M. G.: Use of GPS tracking collars and accelerometers for rangeland livestock production research, Trans. Animal Sci., 2, 81–88, https://doi.org/10.1093/tas/txx006, 2018. a
Bennett, J., Lent, P. C., and Harris, P. J. C.: Dry season foraging preferences of cattle and sheep in a communal area of South Africa, Afr. J. Range Forage Sci., 24, 109–121, https://doi.org/10.2989/AJRFS.2007.24.3.1.294, 2009. a
Cassman, K. G. and Grassini, P.: A global perspective on sustainable intensification research, Nature Sustain., 3, 262–268, https://doi.org/10.1038/s41893-020-0507-8, 2020. a
Castellanos-Navarrete, A., Tittonell, P., Rufino, M. C. C., and Giller, K. E.: Feeding, crop residue and manure management for integrated soil fertility management – A case study from Kenya, Agr. Syst., 134, 1–12, https://doi.org/10.1016/j.agsy.2014.03.001, 2014. a
Conway, D., van Garderen, E. A., Deryng, D., Dorling, S., Krueger, T., Landman, W., Lankford, B., Lebek, K., Osborn, T., Ringler, C., Thurlow, J., Zhu, T., and Dalin, C.: Climate and southern Africa's water–energy–food nexus, Nat. Clim. Change, 5, 837–846, https://doi.org/10.1038/Nclimate2735, 2015. a
Costanza, R., d'Arge, R., de Groot, R., Farber, S., Grasso, M., Hannon, B., Limburg, K., Naeem, S., O'Neill, R. V., Paruelo, J., Raskin, R. G., Sutton, P., and van den Belt, M.: The value of the world's ecosystem services and natural capital, Nature, 287, 253–260, https://doi.org/10.1038/387253a0, 1997. a
Dagliesh, N., Hochman, Z., Huth, N., and Holzworth, D.: Field Protocol to APSoil characterisations, Version 4, Tech. Rep., CSIRO Ecosystem Service, CSIRO Agriculture and Food, Canberra, http://hdl.handle.net/102.100.100/89627?index=1 (last accessed: 24 August 2022), 2016. a
Descheemaeker, K., Oosting, S. J., Homann-Kee Tui, S., P., M., Falconnier, G. N., and Giller, K. E.: Climate change adaptation and mitigation in smallholder crop-livestock systems in sub-Saharan Africa: a call for integrated assessments, Reg. Clim. Change, 16, 2331–2343, https://doi.org/10.1007/s10113-016-0957-8, 2016. a, b
Descheemaeker, K., Zijlstra, M., Masikati, P., Crespo, O., and Homann-Kee Tui, S.: Effects of climate change and adaptation on the livestock component of mixed farming systems: A modelling study from semi-arid Zimbabwe, Agr. Syst., 159, 282–295, https://doi.org/10.1016/j.agsy.2017.05.004, 2018. a, b
Engelbrecht, C. J. and Engelbrecht, F. A.: Shifts in Köppen-Geiger climate zones over southern Africa in relation to key global temperature goals, Theor. Appl. Climatol., 123, 247–261, https://doi.org/10.1007/s00704-014-1354-1, 2016. a
Erenstein, O.: Crop residue mulching in tropical and semi-tropical countries: An evaluation of residue availability and other technological implications, Soil Till. Res., 67, 115–133, https://doi.org/10.1016/S0167-1987(02)00062-4, 2002. a
FAO: Family farmers: Feeding the world, caring for the Earth, World Food Day Celebration Report, Bangkok, http://www.fao.org/3/i4621e/i4621e.pdf (last access: 24 August 2022), 2014. a
FAO: The economic lives of smallholder farmers – an analysis based on household data from nine countries, http://www.fao.org/3/i5251e/i5251e.pdf (last access: 24 August 2022), 2015. a
Franzluebbers, A. J.: Water infiltration and soil structure related to organic matter and its stratification with depth, Soil Till. Res., 66, 197–205, https://doi.org/10.1016/S0167-1987(02)00027-2, 2002. a
Fust, P. and Schlecht, E.: Integrating spatio-temporal variation in resource availability and herbivore movements into rangeland management: RaMDry – An agent-based model on livestock feeding ecology in a dynamic, heterogeneous, semi-arid environment, Ecol. Model., 369, 13–41, 2018. a
Giller, K. E., Rowe, E. C., de Ridder, N., and van Keulen, H.: Resource use dynamics and interactions in the tropics: Scaling up in space and time, Agr. Syst., 88, 8–27, https://doi.org/10.1016/j.agsy.2005.06.016, 2006. a
Hack-ten Broeke, M. D., De Groot, W. J. M., and Dijkstra, J. P.: Impact of excreted nitrogen by grazing cattle on nitrate leaching, Soil Use Manag., 12, 190–198, https://doi.org/10.1111/j.1475-2743.1996.tb00542.x, 1996. a
Hanjra, M. A. and Williams, T. O.: The Role of Smallholder Farms in Food and Nutrition Security, Global Change and Investments in Smallholder Irrigation for Food and Nutrition Security in Sub-Saharan Africa, 99–132, Springer, Cham, https://doi.org/10.1007/978-3-030-42148-9_6, 2020. a
Harvey, C. A., Chacón, M., Donatti, C. I., Garen, E., Hannah, L., Andrade, A., Bede, L., Brown, D., Calle, A., Chará, J., Clement, C., Gray, E., Hoang, M. H., Minang, P., Rodríguez, A. M., Seeberg-Elverfeldt, C., Semroc, B., Shames, S., Smukler, S., Somarriba, E., Torquebiau, E., van Etten, J., and Wollenberg, E.: Climate-smart landscapes: opportunities and challenges for integrating adaptation and mitigation in tropical agriculture, Conserv. Lett., 7, 77–90, https://doi.org/10.1111/conl.12066, 2013. a, b
Herrero, M., Thornton, P. K., Gerber, P., and Reid, R. S.: Livestock, livelihoods and the environment: understanding the trade-offs, Curr. Opin. Environ. Sustain., 1, 111–120, https://doi.org/10.1016/j.cosust.2009.10.003, 2009. a, b
Hoffmann, M. P., Isselstein, J., Rötter, R. P., and Kayser, M.: Nitrogen management in crop rotations after the break-up of grassland: Insights from modelling, Agr. Ecosyst. Environ., 259, 28–44, https://doi.org/10.1016/j.agee.2018.02.009, 2018a. a
Hoffmann, M. P., Odhiambo, J. J. O., Koch, M., Ayisi, K. K., Zhao, G., Soler, A. S., and Rötter, R. P.: Exploring adaptations of groundnut cropping to prevailing climate variability and extremes in Limpopo Province, South Africa, Field Crop. Res., 219, 1–13, https://doi.org/10.1016/j.fcr.2018.01.019, 2018b. a, b, c, d
Hoffmann, M. P., Swanepoel, C. M., Nelson, W. C. D., Beukes, D. J., van der Laan, M., Hargreaves, J. N. G., and Rötter, R. P.: Simulating medium-term effects of cropping system diversification on soil fertility and crop productivity in southern Africa, Eur. J. Agron., 119, 126089, https://doi.org/10.1016/j.eja.2020.126089, 2020. a, b, c, d, e
Hofmeyr, I.: Correct adaptation of livestock on crop residues in winter, Stockfarm, 10, 37–39, 2020. a
Holzworth, D. P., Huth, N. I., Peter, G., Zurcher, E. J., Herrmann, N. I., McLean, G., Chenu, K., van Oosterom, E. J., Snow, V., Murphy, C., Moore, A. D., H., B., Whish, J. P. M., Verrall, S., J., F., Bell, L. W., Peake, A. S., Poulton, P. L., Z., H., Thorburn, P. J., Gaydon, D. S., Dalgliesh, N. P., Rodriguez, D., Cox, H., Chapman, S., Doherty, A., Teixeira, E., Sharp, J., Cichota, R., I., V., Y., L. F., Wang, E., Hammer, G. L., Robertson, M. J., Dimes, J. P., Whitbread, A. M., J., H., van Rees, H., McClelland, T., Carberry, P. S., Hargreaves, J. N. G., Mac Leod, N., C., M., Harsdorf, J., Wedǵwood, S., and Keating, B. A.: APSIM – Evolution towards a new generation of agricultural systems simulation, Environ. Model. Softw., 62, 327–350, https://doi.org/10.1016/j.envsoft.2014.07.009, 2014. a
Keating, B. A., Carberry, P. S., Hammer, G. L., Probert, M. E., Robertson, M. J., Holzworth, D., Huth, N. I., Hargreaves, J. N. G., Meinke, H., Hochman, Z., McLean, G., Verburg, K., Snow, V., Dimes, J. P., Silburn, M., Wang, E., Brown, S., Bristow, K. L., Asseng, S., Chapman, S., McCown, R. L., Freebairn, D. M., and Smith, C. J.: An overview of APSIM, a model designed for farming systems simulation, Eur. J. Agron., 18, 267–288, https://doi.org/10.1016/S1161-0301(02)00108-9, 2003. a
Kersebaum, K. C., Boote, K. J., Jorgenson, J. S., Nendel, C., Bindi, M., Frühauf, C., Gaiser, T., Hoogenboom, G., Kollas, C., Olesen, J. E., Rötter, R. P., Ruget, F., Thorburn, P. J., Trnka, M., and Wegehenkel, M.: Analysis and classification of data sets for calibration and validation of agro-ecosystem models, Environ. Model. Softw., 72, 402–417, https://doi.org/10.1016/j.envsoft.2015.05.009, 2015. a
Kitajima, K., Mulkey, S. S., and Wright, S. J.: Decline of photosynthetic capacity with leaf age in relation to leaf longevity for five tropical canopy tree species, Am. J. Bot., 84, 702–708, https://doi.org/10.2307/2445906, 1997. a
Lam, Q. D., Rötter, R. P., Bracho-Mujica, G., Ferreira, N. C. R., Foord, S. H., Nelson, W. C. D., Odhiambo, J., and Ayisi, K.: Climate change impacts on the water resources of subhumid to semi-arid landscapes of Limpopo in southern Africa, in review, J. Agr. Water Manage., 2022. a
Lamega, S. A., Komainda, M., Hoffmann, M. P., Ayisi, K. K., Odhiambo, J. J., and Isselstein, J.: It depends on the rain: Smallholder farmers' perceptions on the seasonality of feed gaps and how it affects livestock in semi-arid and arid regions in Southern Africa, Clim. Risk Manag., 34, 100362, https://doi.org/10.1016/j.crm.2021.100362, 2021. a
Langan, L., Higgins, S., and Scheiter, S.: Climate-biomes, pedo-biomes or pyro-biomes: which world view explains the tropical forest – savanna boundary in South America?, J. Biogeogr., 44, 2319–2330, https://doi.org/10.1111/jbi.13018, 2017. a, b
Lehman, R. M., Osborne, S. L., and Duke, S. E.: Diversified no-till crop rotation reduces nitrous oxide emissions, increases soybean yields, and promotes soil carbon accrual, Soil Sci. Soc. Am. J., 81, 76–83, https://doi.org/10.2136/sssaj2016.01.0021, 2017. a
Magomeyi, M. S., Taigbenu, A. E., and Barron, J.: Effectiveness of agricultural water management technologies on rainfd cereal crop yield and runoff in semi-arid catchment: a meta-analysis, Int. J. Agr. Sustain., 16, 418–441, https://doi.org/10.1080/14735903.2018.1523828, 2018. a
Mediavilla, S. and Escudero, A.: Photosynthetic capacity, integrated over the lifetime of a leaf, is predicted to be independent of leaf longevity in some tree species, New Phytol., 159, 203–211, 2003. a
Millennium Ecosystem Assessment: Ecosystems and Human Well-being: Synthesis, Island Press, Washington, DC, ISBN 1-59726-040-1, 2005. a
Mohler, C. L. and Johnson, S. E.: Crop rotation on organic farms: a planning manual. Ithaca, NY: Natural Resource, Agriculture, and Engineering Service (NRAES) Cooperative Extension, ISBN 978-1-933395-21-0, https://doi.org/10.1046/j.0028-646x.2003.00798.x, 2009. a
Mueller, N., Gerber, J. S., Johnston, M., Ray, D. K., Ramankutty, N., and Foley, J. A.: Closing yield gaps through nutrient and water management, Nature, 490, 254–257, https://doi.org/10.1038/nature11420, 2012. a
Müller, B., Schulze, J., Kreuer, D., Linstädter, A., and Frank, K.: How to avoid unsustainable side effects of managing climate risk in drylands – The supplementary feeding controversy, Agr. Syst., 139, 153–65, https://doi.org/10.1016/j.agsy.2015.07.001, 2015. a
Nelson, W. C. D., Hoffmann, M. P., May, C., Mashao, F., Ayisi, K., Odhiambo, J., Bringhenti, T., Feil, J. H., Bakhsh, S. Y., and Abdulai, I.: Tackling climate risk to sustainably intensify smallholder maize farming systems in southern Africa, Environ. Res. Lett., 17, 075005, https://doi.org/10.1088/1748-9326/ac77a3, 2022. a, b
Parry, K., van Rooyen, A. F., Bjornlund, H., Kissoly, L., Moyo, M., and de Sousa, W.: The importance of learning processes in transitional small-scale irrigation schemes, Int. J. Water Resour. D., 36, 199–223, https://doi.org/10.1080/07900627.2020.1767542, 2020. a, b
Pfeiffer, M., Langan, L., Linstädter, A., Martens, C., Gaillard, C., Ruppert, J. C., Higgins, S. I., Mudongo, E. I., and Scheiter, S.: Grazing and aridity reduce perennial grass abundance in semi-arid rangelands – Insights from a trait-based dynamic vegetation model, Ecol. Model., 395, 11–22, 2019. a, b, c, d, e, f, g
Pfeiffer, M., Hoffmann, M. P., Scheiter, S., Nelson, W., Isselstein, J., Ayisi, K., Odhiambo, J. J., and Rötter, R. P.: Data and Data Processing Scripts for the Publication “Modeling the Effects of Alternative Crop-Livestock Management Scenarios on Important Ecosystem Services for Smallholder Farming from a Landscape Perspective”, Open Science Framework (OSF) [data set and code], https://doi.org/10.17605/OSF.IO/R2TS7, 2022. a
Probert, M. E., Dimes, J. P., Keating, B. A., Dalal, R. C., and Strong, W. M.: APSIM's water and nitrogen modules and simulation of the dynamics of water and nitrogen in fallow systems, Agr. Syst., 56, 1–28, https://doi.org/10.1016/S0308-521X(97)00028-0, 1998. a
Rapholo, E., Odhjambo, J. J. O., Nelson, W. C. D., Rötter, R. P., Ayisi, K., Koch, M., and Hoffmann, M. P.: Maize–lablab intercropping is promising in supporting the sustainable intensification of smallholder cropping systems under high climate risk in southern Africa, Exp. Agr., 56, 104–117, https://doi.org/10.1017/S0014479719000206, 2019. a
Rosenzweig, C., Jones, J. W., Hatfield, J. L., Ruane, A. C., Boote, K. J., Thorburn, P., Antle, J. M., Nelson, G. C., Porter, C., Janssen, S., Asseng, S., Basso, B., Ewert, F., Wallach, D., Baigorria, G., and Winter, J. M.: The Agricultural Model Intercomparison and Improvement Project (AgMIP): Protocols and pilot studies, Agr. Forest Meteorol., 170, 166–182, https://doi.org/10.1016/j.agrformet.2012.09.011, 2013. a
Rötter, R. P. and Van Keulen, H.: Variations in yield response to fertilizer application in the tropics: II. Risks and opportunities for smallholders cultivating maize on Kenya's arable land, Agr. Syst., 53, 69–95, https://doi.org/10.1016/S0308-521X(96)00037-6, 1997. a
Rötter, R. P., Scheiter, S., Hoffmann, M., Pfeiffer, P., Nelson, M., Ayisi, K., Taylor, P., Feil, J.-H., Bakhsh, S. Y., Isselstein, J., Linstädter, A., Behn, K., Westphal, K., Odhiambo, J., Wayne, T., Grass, I., Merante, P., Bracho-Mujica, G., Bringhenti, T., Lamega, S., Abdulai, I., Lam, Q. D., Anders, M., Linden, V., Weier, V., Foord, S., and Erasmus, B.: Modeling the multi-functionality of African savanna landscapes under global change, Land Degrad. Dev., 32, 2077–2081, https://doi.org/10.1002/ldr.3925, 2021. a, b
Ruane, A. C., Goldberg, R., and Chryssantacopoulos, J.: AgMIP climate forcing datasets for agricultural modeling: Merged products for gap-filling and historical climate series estimation, Agr. Forest Meteorol., 200, 233–284, https://doi.org/10.1016/j.agrformet.2014.09.016, 2015. a, b
Rufino, M. C., Tittonell, P., Reidsma, P., López-Ridaura, S., Hengsdijk, H., Giller, K. E., and Verhagen, A.: Network analysis of N flows and food self-sufficiency – a comparative study of crop-livestock systems of the highlands of East and southern Africa, Agr. Forest Meteorol., 200, 233–284, https://doi.org/10.1016/j.agrformet.2014.09.016, 2015. a
Rusinamhodzi, L., Corbeels, M., van Wijk, M. T., Rufino, M. C., Nyamangara, J., and Giller, K. E.: A meta-analysis of long-term effects of conservation agriculture on maize grain yield under rain-fed conditions, Agron. Sustain. Dev., 31, 657–673, https://doi.org/10.1007/s13593-011-0040-2, 2011. a
Scheiter, S. and Higgins, S. I.: Impacts of climate change on the vegetation of Africa: an adaptive vegetation modelling approach (aDGVM), Glob. Change Biol., 15, 2224–2246, https://doi.org/10.1111/j.1365-2486.2008.01838.x, 2009. a
Scheiter, S., Langan, L., and Higgins, S. I.: Next generation dynamic global vegetation models: learning from community ecology, New Phytol., 198, 957–969, https://doi.org/10.1111/nph.12210, 2013. a, b
Scheiter, S., Schulte, J., Pfeiffer, M., Martens, C., Erasmus, B. F. N., and Twine, W. C.: How does climate change influence the economic value of ecosystem services in savanna rangelands, Ecol. Econ., 157, 342–356, https://doi.org/10.1016/j.ecolecon.2018.11.015, 2019. a
Schrader, L.: Rinderzucht und Fleischerzeugung – Empfehlungen für die Praxis, chap. Verhalten und Tierhaltung, 89–107, Landbauforschung Völkenrode, Sonderheft 313, https://literatur.thuenen.de/digbib_extern/dk038909.pdf (last access: 24 August 2022), 2007. a
Smith, T.: Some tools to combat dry season nutritional stress in ruminants under African conditions, Proceedings of the Final Review Meeting of an IAEA Technical Cooperation, Regional AFRA Project IAEA-TeCDOC-1294, https://inis.iaea.org/collection/NCLCollectionStore/_Public/33/032/33032982.pdf?r=1, (last access: 24 August 2022), 2002. a
Sumberg, J.: Toward a dis-aggregated view of crop-livestock integration in Western Africa, Land Use Policy, 20, 253–264, https://doi.org/10.1016/S0264-8377(03)00021-8, 2003. a
Suzuki, S., Noble, A. D., Ruaysoongnern, S., and Chinabut, N.: Improvement in water-holding capacity and structural stability of a sandy soil in northeastern Thailand, Arid Land Res. Manag., 21, 37–49, https://doi.org/10.1080/15324980601087430, 2007. a
Tarawali, S., Herrero, M., Descheemaeker, K., Grings, E., and Blümmel, M.: Pathways for sustainable development of mixed crop livestock systems: Taking a livestock and pro-poor approach, Livest. Sci., 139, 11–21, https://doi.org/10.1016/j.livsci.2011.03.003, 2011. a
Thornton, P. K. and Herrero, M.: Adapting to climate change in the mixed crop and livestock farming systems in sub-Saharan Africa, Nat. Clim. Change, 5, 830–836, https://doi.org/10.1038/nclimate2754, 2015. a
Tilman, D., C., B., Hill, J., and Befort, B. L.: Global food demand and the sustainable intensification of agriculture, P. Natl. Acad. Sci. USA, 108, 20260–20264, https://doi.org/10.1073/pnas.1116437108, 2011. a
Tscharntke, T., Clough, Y., Wanger, T. C., Jackson, L., Motzke, I., Perfecto, I., Vandermeer, J., and Whitbread, A.: Global food security, biodiversity conservation and the future of agricultural intensification, Biol. Conserv., 151, 53–59, https://doi.org/10.1016/j.biocon.2012.01.068, 2012. a
UN General Assembly: Transforming our World: the 2030 Agenda for Sustainable Development, A/RES/70/1, https://www.refworld.org/docid/57b6e3e44.html (last access: 24 August 2022), 2015. a
Valbuena, D., Erenstein, O., Homann-Kee Tui, S., Abdoulaye, T., Claessens, L., Duncan, A. J., Gérard, B., Rufino, M. C., Teufel, N., van Rooyen, A., and van Wijk, M. T.: Conservation Agriculture in mixed crop-livestock systems: Scoping crop residue trade-offs in Sub-Saharan Africa and South Asia, Field Crop. Res., 132, 175–184, https://doi.org/10.1016/j.fcr.2012.02.022, 2012. a
van de Ven, G. W. J., de Ridder, N., van Keulen, H., and van Ittersum, M. K.: Concepts in production ecology for analysis and design of animal and plant-animal production systems, Agr. Syst., 76, 507–525, https://doi.org/10.1016/S0308-521X(02)00110-5, 2003. a
Vanlauwe, B. and Dobermann, A.: Sustainable intensification of agriculture in sub-Saharan Africa: first things first!, Front. Agr. Sci. Eng., 7, 376–382, https://doi.org/10.15302/J-FASE-2020351, 2020. a
Vignola, R., Harvey, C. A., Bautista-Solis, P., Avelino, J., Rapidel, B., Donatti, C., and Martinez, R.: Ecosystem-based adaptation for smallholder farmers: Definitions, opportunities and constraints, Agr. Ecosyst. Environ., 211, 126–132, https://doi.org/10.1016/j.agee.2015.05.013, 2015. a
Waha, K., van Wijk, M. T., Fritz, S., See, L., Thornton, P. K., Wichern, J., and Herrero, M.: Agricultural diversification as an important strategy for achieving food security in Africa, Glob. Change Biol., 24, 3390–3400, https://doi.org/10.1111/gcb.14158, 2018. a
Walpole, M., Smith, J., Rosser, A., Brown, C., Schulte-Herbruggen, B., Booth, H., Sassen, M., Mapendeme, A., Fancourt, M., Bieri, M., Glaser, S., Corrigan, C., Narloch, U., Runsten, L., Jenkins, M., Gomera, M., and Hutton, J. M.: Smallholders, food security, and the environment, Unep technical report, IFAD, https://www.ifad.org/documents/38714170/39135645/smallholders_report.pdf/133e8903-0204-4e7d-a780-bca847933f2e (last access: 24 August 2022), 2013. a, b
Wang, E., Bell, M., Luo, Z., Moody, P., and Probert, M. E.: Modelling crop response to phosphorus inputs and phosphorus use efficiency in a crop rotation, Field Crop. Res., 155, 120–132, https://doi.org/10.1016/j.fcr.2013.09.015, 2014. a
Whitbread, A. M., Robertson, M. J., Carberry, P. S., and Dimes, J. P.: How farming systems simulation can aid the development of more sustainable smallholder farming systems in southern Africa, Eur. J. Agron., 32, 51–58, https://doi.org/10.1016/j.eja.2009.05.004, 2010. a, b
Whitbread, A. M., Hoffmann, M. P., Davoren, W., Mowat, D., and Baldock, J. A.: Measuring and Modeling the Water Balance in Low-Rainfall Cropping Systems, T. ASABE, 60, 2097–2110, https://doi.org/10.13031/trans.12581, 2017. a
Williams, A., Jordan, N. R., Smith, R. G., Hunter, M. C., Kammerer, M., A., K. D., Koide, R. T., and Davis, A. S.: A regionally-adapted implementation of conservation agriculture delivers rapid improvements to soil properties associated with crop yield stability, Sci. Rep., 8, 8467, https://doi.org/10.1038/s41598-018-26896-2, 2018. a