the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 26 Aug 2025
| 26 Aug 2025
Changes in soil organic and inorganic carbon with elevation in a dry alpine rangeland of the northern Qinghai–Tibet Plateau
Qinglin Liu
Ailin Zhang
Xiangyi Li
Jinfei Yin
Yuxue Zhang
Osbert Jianxin Sun
Yong Jiang
The spatial patterns of soil carbon (C) in water constrained alpine ecosystems have rarely been investigated. It remains unclear how changes in biotic and abiotic factors with elevation would shape the distribution of soil C stocks when plant communities are co-limited by water and low temperature. To address this uncertainty, we systematically set up seven sampling points along an elevational gradient between 3000 and 4000 m above sea level and investigated the patterns of changes with elevation in the surface soil organic C (SOC) and soil inorganic C (SIC) in the northern part of the Qinghai–Tibet Plateau. Our results showed that the total soil C density (TCD) and the SOC density (SOCD) increased with rising elevation, but the SIC density (SICD) displayed a pattern of nonlinear change with a peak at the mid-slope of the elevational range. While SIC dominated the soil C pool, accounting for 64 %–90 % of TCD, the proportion of SOC increased from 10 % of the TCD at the lower range of the elevational gradient to 36 % at the upper range. The increases in SOCD with elevation were associated with changes from scrub dominated vegetation cover to herbaceous plant communities and decreasing MAT, suggesting a dual mechanism SOC accumulation at the higher elevation by increased level of plant derived C inputs and reduced SOC mineralization. In contrast, variations in SICD were mainly explainable by changes in soil C to N ratio and soil water content (SWC) and likely resulted from non-linear changes in factors related to inorganic C production and leaking losses. Findings from this study help fill the knowledge gap on the underlying controls of SOC and SIC distribution with changes in elevation in water and low temperature constrained alpine rangeland.
- Article
(2358 KB) - Full-text XML
-
Supplement
(375 KB) - BibTeX
- EndNote




Because soil contains the largest proportion of carbon (C) stocks in terrestrial ecosystems (Lal, 2018), the size, persistence and storage capacity of the soil C pool have been the focal issues in global change research. However, despite extensive studies on soil C in the past, there is still some degree of uncertainty regarding the response of soil C pools to global climate change. Part of the problem arises from differential alterations of pool size and functional structure of soil C among the world's terrestrial ecosystems as affected by environmental variability and climate change (Trumbore, 1997; Zhang et al., 2024). The soil C pool is made up of both soil organic C (SOC) and soil inorganic C (SIC) chemical compounds. In general, SOC dominates the soil C pool on well vegetated sites (Feyissa et al., 2023) and SIC constitutes a predominant component of soil C pool in drylands where plants tend to be scarce (Du and Gao, 2020). Previous research has well demonstrated that SOC is jointly controlled by vegetation, climate,, and soil physicochemical properties (Eswaran et al., 1993; Torn et al., 1997; Schuur et al., 2001; Callesen et al., 2003; Sun et al., 2004). In contrast, SIC pool is mainly affected by abiotic factors such as the property of soil parent material, climate and topography (Chang et al., 2012; Ma et al., 2022; Dong et al., 2024). The patterns of changes in soil organic C and silicon carbide reservoirs with elevation and vegetation types in dry alpine rangeland remain unclear.
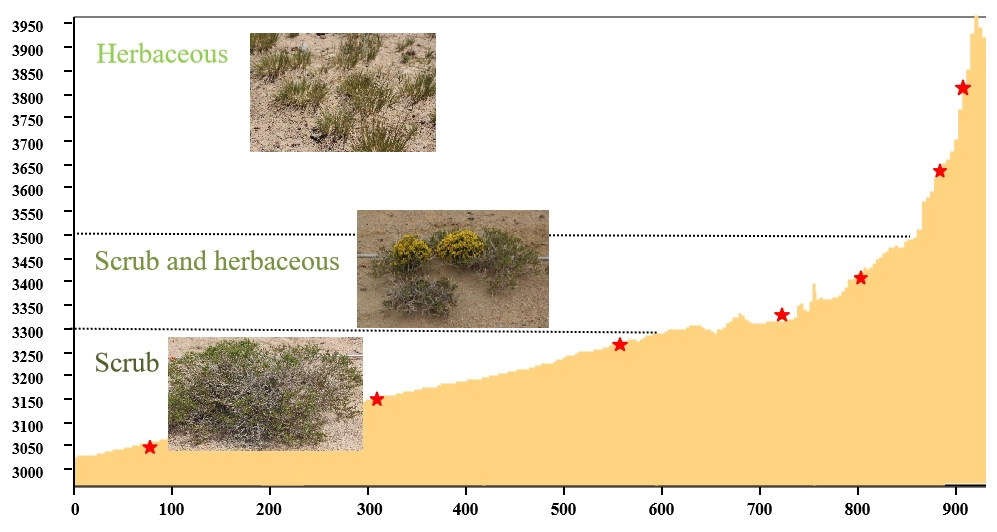
Figure 1The vertical profile of sample sites in the northern section of the Altun Mountain Nature Reserve, Qinghai–Tibet Plateau.
The Qinghai–Tibet Plateau rises sharply in elevation and contains diverse ecosystem types with contrasting climate, vegetation and soil characteristics. With rising elevation along the mountain slopes on the Plateau, air temperature markedly decreases and precipitation and the intensity of solar radiation increases, contributing to elevational changes in vegetation and soil nutrient availability (Tiemann and Billings, 2011; García-Palacios et al., 2013; Wang et al., 2024). The unique topographic features and presence of diverse ecosystem types have made the region a hotspot for research geared at better understanding of the impacts by climate change on ecosystem structure and function. However, most studies in this region have mainly focused on characterization of vegetation in relation to site conditions as well as determination of the responses of alpine meadow steppes to general climate change factors such as temperature and precipitation (Wang et al., 2023; Chen et al., 2017; Cai et al., 2025). There has been a lack of explicit information on the distribution of soil C pools in dry alpine rangeland, which constitutes an important part of the Qinghai–Tibet Plateau (Zhang et al., 2021). On the Qinghai–Tibet Plateau, the abiotic conditions, vegetation, and soil types along the elevational gradient vary greatly and experience different climate change impacts (Li et al., 2017). The elevational changes in vegetation type and hydrothermal conditions would inevitably result in variations in soil C pool size and dynamics (Rodeghiero and Cescatti, 2005). For example, it has been found that both the quality and quantity of SOC substantially vary along elevational gradients in mountainous landscapes (Pepin et al, 2015). The alpine grassland in the northern part of the Qinghai–Tibet Plateau is under the influence of an arid climate, with a relatively low vegetation coverage. The surface is severely eroded by wind and water, and the community type is mainly desert grassland. Under such conditions SIC plays a more dominant role in the soil C storage (Batjes, 2006; Du and Gao, 2020). Previous studies in the region have mostly neglected the role of SIC in the soil C inventory. Whether, or to what extent, the elevational changes in micro-environments and vegetation affect SOC and SIC yet remains an open question.
In this study, we investigated the patterns of changes in SOC and SIC in the top 30 cm soil along an elevational gradient of 3000–4000 m above sea level (a.s.l.) on the southern slope of Altun Mountain in the northern Qinghai–Tibet Plateau. Data were collected for characterization of vegetation and soil physicochemical properties. The aims of the study were to determine the elevational patterns in the density of SOC (SOCD) and SIC (SICD). We hypothesized that: (1) with decreases in temperature along the elevational gradient, slow-turnover woody plants give way to fast-turnover herbaceous plants due to energy constraints, leading to greater SOC preservation at the higher elevational sites as a result of a dual mechanism of greater plant derived C inputs and lower rate of decomposition; and (2) SIC dominates the soil C pool and would not display an apparent trend of variations with elevation as it is predominantly determined by soil parent materials and influenced by abiotic factors.
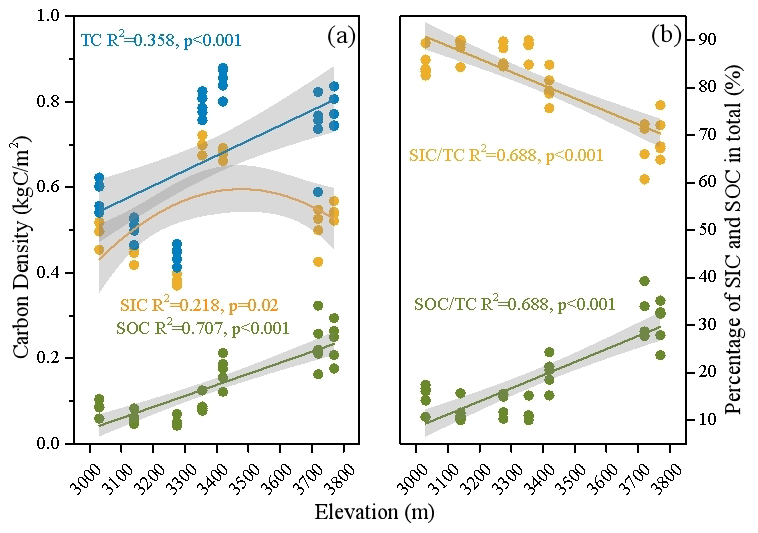
Figure 2Elevation changes in (a) densities of soil organic C (SOC), inorganic C (SIC), and total C (TC), and (b) proportions of SOC and SIC in TC in a dry alpine rangeland of the Qinghai–Tibet Plateau.
2.1 Study sites and experimental design
Our study sites are located in the Altun Mountain Nature Reserve in the south of Altyn Tagh, situated in the northeastern part of the Qinghai–Tibet Plateau (87°10′–91°18′ E, 36–37°49′ N). This area is known for harsh environmental conditions characterized by a dry climate, with an average annual temperature of 0 °C and an annual precipitation of around 110 mm. The soils are predominantly Yermosols (FAO, http://www.fao.org/soils-portal/so, last access: 15 March 2020). The reserve comprises diverse land cover types, including desert, scrub, and grassland. The herbaceous layer typically ranges from 5 to 20 cm in height, with a coverage of 10 %–30 %, occasionally reaching 60 %–80 %. The main vegetation types is dwarf scrub in the lower elevational range of the slope and grassland at the upper slope. In this study area, the vegetation types are mainly small shrubs at elevations of 3000–3300 m, mixed shrubs and herbs at elevations of 3300–3500 m, and herbs above 3500 m (Fig. 1). Dominant plant species are represented by Stipa purpurea Griseb. and Kobresia robusta Maxim., which are often accompanied by common grassland plants including Carex kunlumsannsis N. R. Cui, Koeleria cristata (L.) Pers., and Oxytropis falcata Bunge.
In August 2019, we conducted a plant community survey and soil sampling at seven representative elevations (designated as A1–A7) along an elevational gradient of 3000–4000 m a.s.l. in the northern section of the Altun Mountain Nature Reserve. The vertical distance between adjacent sampling elevations ranged from 60 to > 100 m. All sites are georeferenced. At each sample point, a 100 m × 100 m standard plot was set up for vegetation survey and sampling. Measurements of plant communities and soil sampling were made with 1 m × 1 m quadrats evenly spaced along the mid-line of each plot perpendicular to the contour. The elevational profile of the sampling sites is illustrated in Fig. 1.
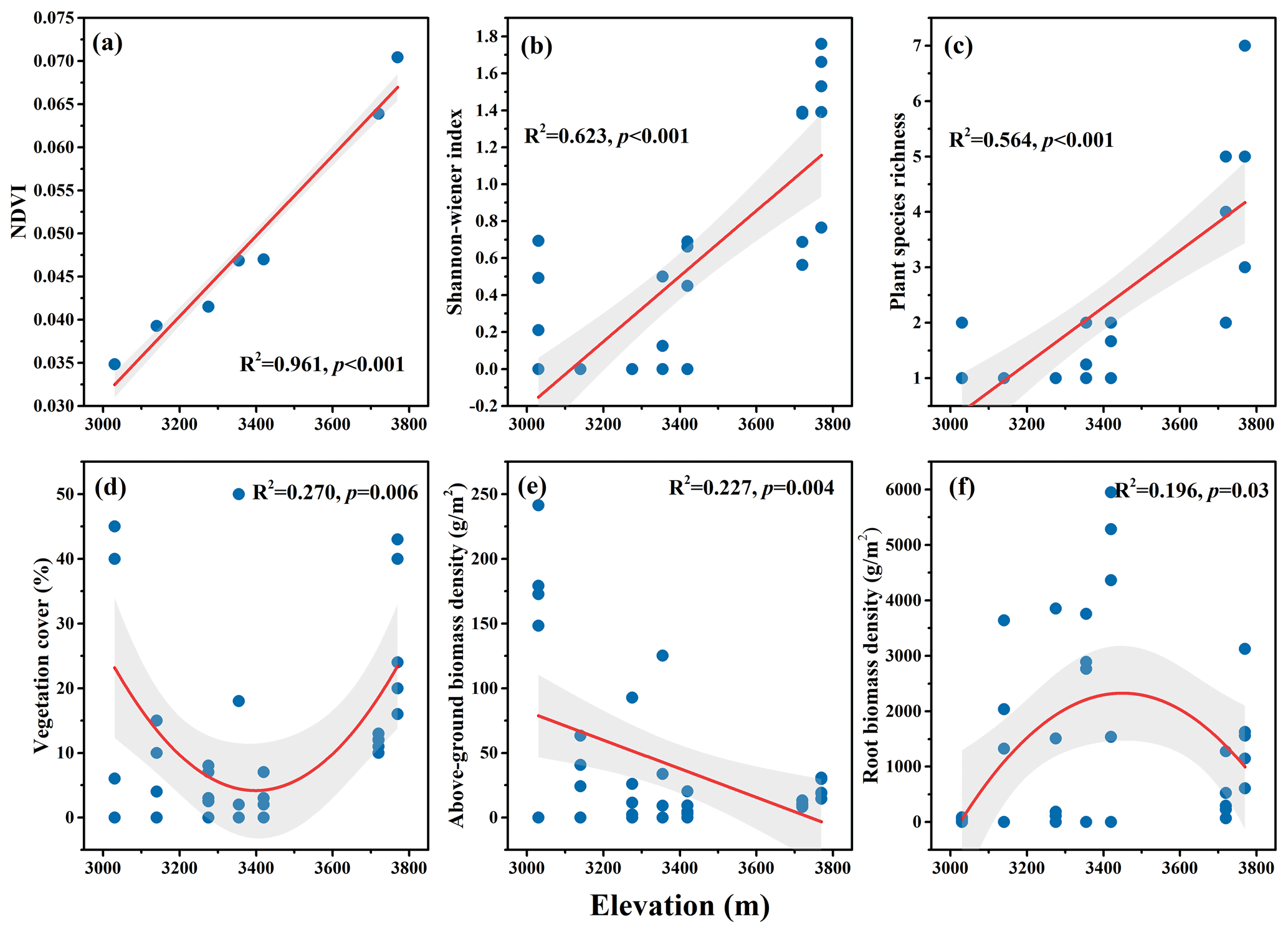
Figure 3The elevational variation of selective plant community characteristics in a dry alpine rangeland of the Qinghai–Tibet Plateau. (a) Normalized difference vegetation index (NDVI); (b) Shannon–Wiener index; (c) plant species richness; (d) vegetation cover; (e) aboveground biomass density; and (f) fine root biomass density.
2.2 Measurements of plants and soil
We measured the relative coverage of each plant species and the entire community and identified the dominant species. The coverage of the plant community was calculated as the sum of the coverage values for individual species as there are little overlaps among plant species at our study sites. The aboveground tissues of all plants within each quadrat were harvested and measured for both fresh and dry mass. Soil samples were collected to 30 cm depth using a 7 cm (inner diameter) augur at locations where the plants were harvested. Roots were picked out of soil samples and measured for dry mass weight. Upon completing the field survey, plant samples were transported to a laboratory and oven dried at 75 °C for 48 h for determination of biomass. Soil water content (SWC) was determined gravimetrically by determining the fresh soil weight and then dry mass after subjecting to oven drying at 105 °C for 48 h. Soil electrical conductivity and pH were determined using a conductometer (1:1 soil : water suspension) and acidimeter (1:5 soil : water suspension). Soil bulk density (BD) was determined using the cutting ring method. The SOC and plant C contents (aboveground tissues and roots) were measured using the K2Cr2O7 oxidation method and soil TC was measured using an elemental analyser (TOCV wp; Shimadzu Corp., Tokyo, Japan). The content of SIC was directly determined by neutralization titration. The measurements for plant C were made with oven dried samples.
2.3 Data processing and statistical analysis
The climate data (MAT and NDVI) used in this study were extracted from the website of global climate data (http://worldclim.org, last access: 03 May 2020). For plant community structure, we quantified the Shannon–Wiener index (H′) and species richness (Whittaker and Niering, 1965). One-way ANOVA was used to determine the effects of elevation on SWC and BD, and Duncan's multiple comparison test was used to determine the statistical significance of the differences of the variables among the elevational sites. Linear regression was applied to examine the relationships of SOC and SIC with the indices of climate, plant communities, and soil. Redundancy analysis (RDA) was performed for identification of significant factors influencing SOC and SIC. The direct and indirect influences of climate, plant, and soil variables on SOC and SIC were analysed with structural equation modelling. We developed structural equation models (SEMs) for SOCD and SICD that simultaneously consider vegetation, environmental factors, and soil properties. Based on the results of linear regression and RDA, the variables associated with the three potential predictors for SOCD are NDVI, species diversity, species richness, MAT, BD, and soil N content. In contrast, the variables associated with the three potential predictors for SICD are root biomass, SWC, BD, and soil . The sample size for calculating each index is 35. The initial model is presented in the Supplementary materials (Fig. S1). We acknowledge that the influence of certain excluded variables on SOCD and SICD may have been overlooked. Nonetheless, by minimizing the model's complexity, we could gain better insights on the primary direct and indirect effects of vegetation, environmental factors, and soil properties on the soil carbon pool. This approach allows us to address our research questions more comprehensively. All variables included in this study were initially categorized into composite groups before being incorporated into the SEM. To assess the robustness of the relationships of key ecosystem factors with SOCD and SICD, we employed piecewise SEM to analyse the random effects at the sampling sites, detailing the “marginal” and “conditional” contributions of the predictive factors (Tian et al., 2021). All statistical analyses were implemented within Origin 9.3 and R 4.2.1, and composite SEM model analysis was performed using the “piecewise SEM”, “nlme” and “lme4” procedures in R.
3.1 Changes in soil C pools, plant community, climate, and edaphic factors with elevation
The soil total C (TC) pool was predominantly made up of inorganic C components along the elevation gradient, which accounted for 64 %–90 % of TC (Fig. 2). With increases in elevation, the density of SOC significantly and linearly increased (Fig. 2a), leading to an increased proportion of SOC in TC from 10 %–36 % (Fig. 2b).
Among the variables characterizing plant communities, NDVI, plant species diversity, and richness all increased with rising elevation (p<0.01) (Fig. 3a–c); aboveground biomass density decreased (p<0.01) (Fig. 3e). However, both fine root biomass density and vegetation cover displayed curvilinear change patterns with elevation (Fig. 3d and f).
Among the climatic and edaphic variables, there was a significant linear decrease in MAT (p<0.01; Fig. 4b) and a significant linear increase in soil N content with increases in elevation (p<0.01; Fig. 4d). Soil pH, BD, and all exhibited a hump shaped pattern of change along the elevational gradient (p<0.01; Fig. 4c, e, f).
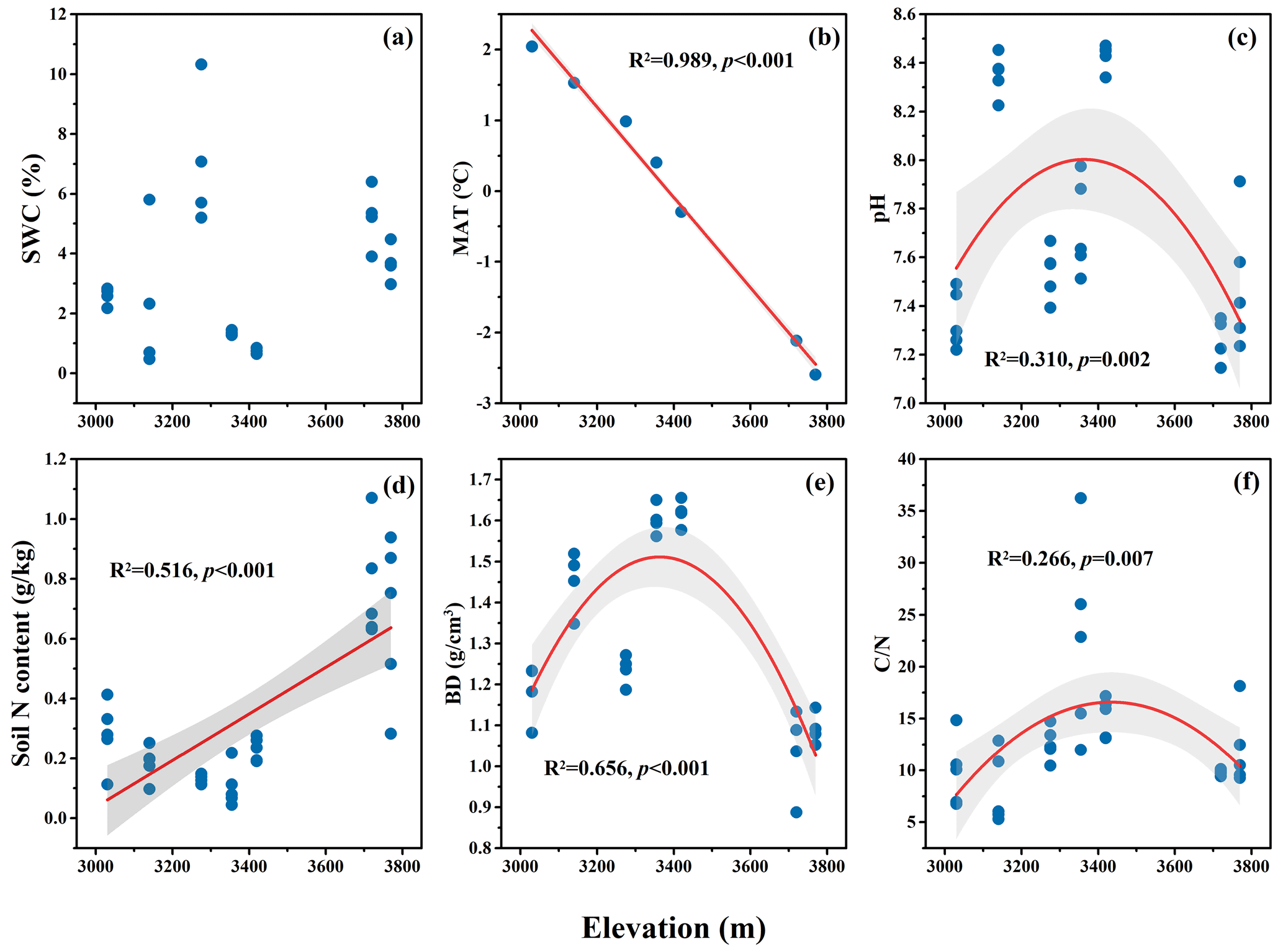
Figure 4The elevational variation of selective climatic and soil variables in a dry alpine rangeland of Qinghai–Tibet Plateau. (a) Soil water content (SWC); (b) mean annual temperature (MAT); (c) soil pH; (d) soil N content; (e) soil bulk density (BD); (f) soil C to N ratio ().
3.2 Influencing factors on changes in soil organic C density (SOCD) and inorganic C density (SICD)
The results from both correlation analysis and RDA showed that SOCD had a significant negative correlation with MAT, and significant positive correlations with BD, soil N content, NDVI, and plant diversity (Table 1; Fig. 5). In contrast, SICD was negatively correlated with SWC and positively with BD, soil , and fine root biomass density (Table 1; Fig. 5). In general, changes in SOCD were mostly explainable by variables related to climate and plant community traits, whereas SICD was predominantly associated with edaphic factors.
Table 1Summary of the correlation coefficients for relationships of SOCD and SICD with selective variables for climatic, edaphic and plant community characteristics. * p<0.05, p<0.01.
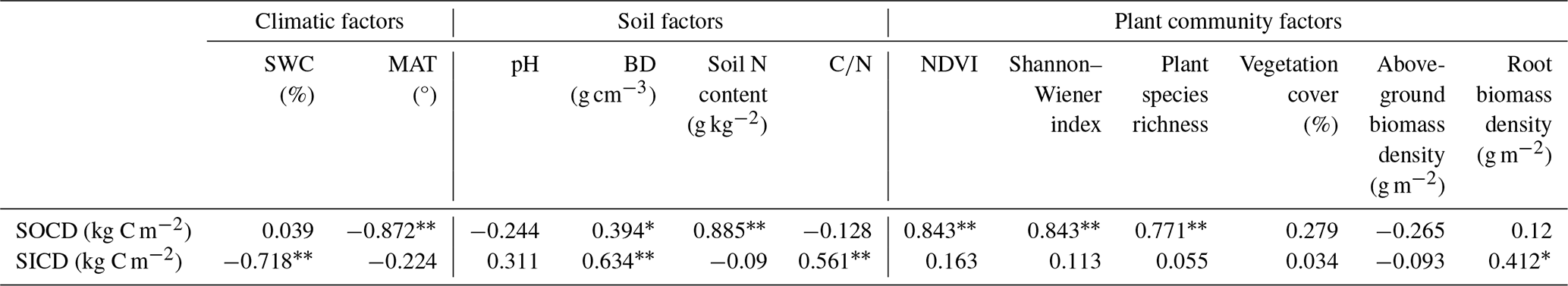
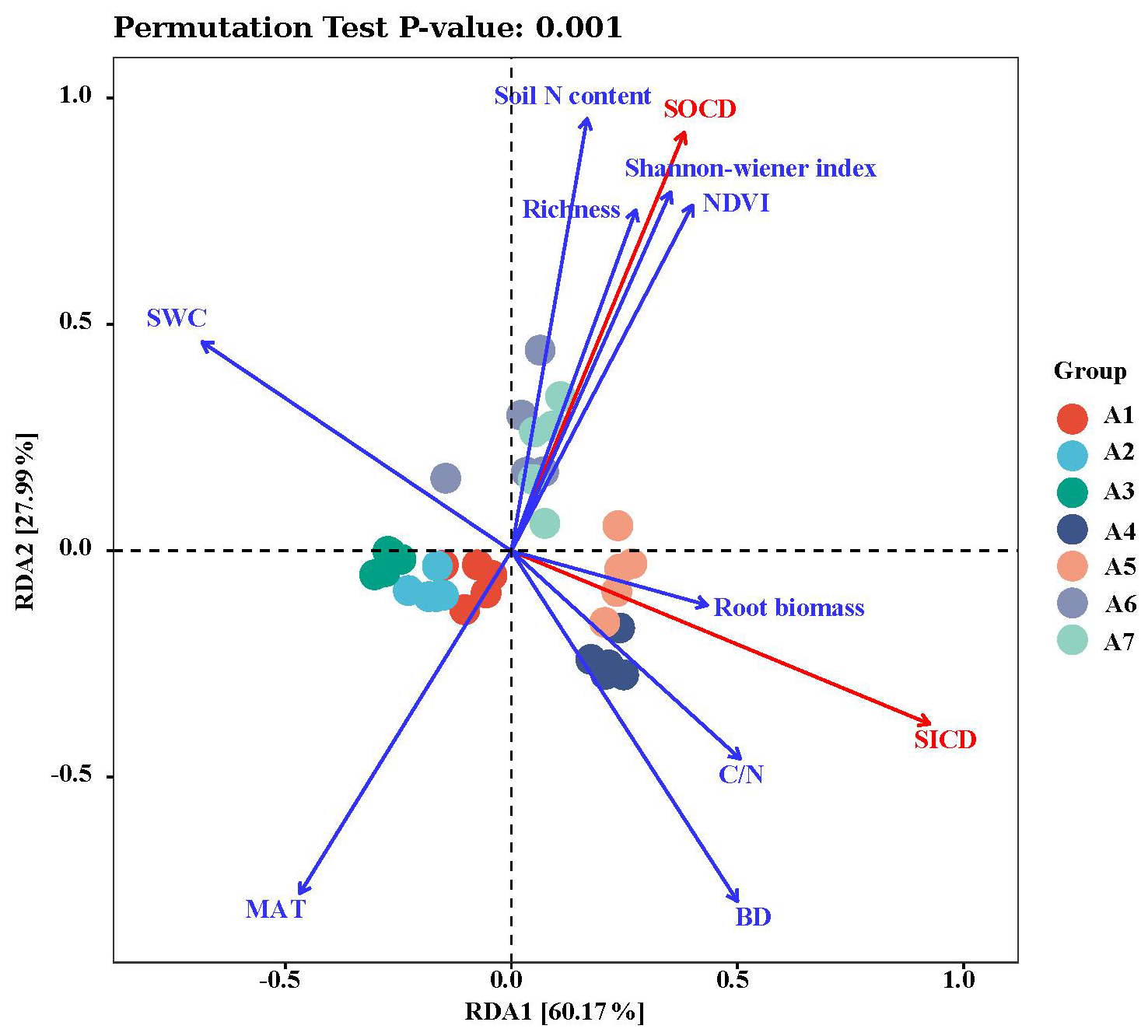
Figure 5RDA ranking of soil C pool (red lines) and environmental variables (blue lines) at different elevations. Arrow lines represent relative values of environmental variables and soil C pools. Correlations between environmental variables and soil C pools are indicated by the cosine of angles between the corresponding arrow lines; angles < 90° indicate a positive correlation, and > 90° a negative correlation. Projecting the arrow line for a soil C pool into an arrow line for a corresponding environmental variable, the distance from the origin to the projection point indicates the relative power of the environmental variable in explaining the size of soil C pools.
In the SEMs, the effects of elevation on SOCD were implemented via modifications of climate and plant communities (Fig. 6a) whereas variations in SICD were mainly associated with edaphic factors (Fig. 6b), consistent with the results from the RDA.
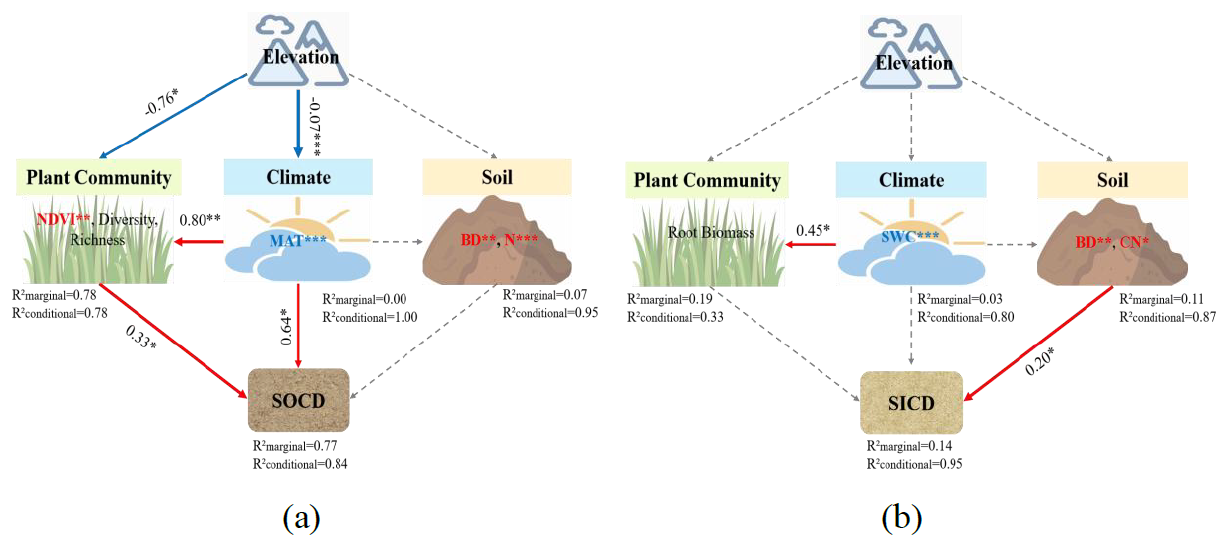
Figure 6Structural equation models (SEMs) of the influences on (a) soil organic C density (SOCD) and (b) soil inorganic C density (SICD) by elevation, climate, plant community, and soil. (a) Fisher's C = 4.714; p=0.318; df=4; AIC = 44.714; BIC = 75.821; (b) Fisher's C = 0.000; p=0.287; df=4; AIC = 45.000; BIC = 76.107. Numbers adjacent to arrows are the standardized path coefficients (equivalent to correlation coefficients). Arrow thickness indicates the strength of relationships. Red solid arrows denote significant positive effects (p<0.05) or marginally significant (0.05<p<0.1) effects. Blue solid arrows denote significant negative effects (p<0.05) or marginally significant (0.05<p<0.1) effects. R2 values associated with response variables indicate the variance accounted for by the model. * p<0.05; p<0.01; p<0.001.
The soil C pool, as the largest C stocks in terrestrial ecosystems, has been extensively studied at different scales and for various regions (Zhang et al., 2024; Chalchissa and Kuris, 2024). However, previous studies have not paid much attention to the contributions of different C components in extreme environments. We studied the elevational patterns of soil organic and inorganic C pools in an alpine rangeland where ecosystem processes are co-limited by drought and low temperature. Contrary to previous studies reporting that the soil carbon pool of alpine meadows and moist grasslands is predominantly organic (Chen et al., 2017; Chen et al., 2022), our findings indicate that inorganic carbon dominates in the soils of arid, alpine plateaus. Our results showed a linear increase in the soil organic C pool with rising elevation. We found that the soil organic C pool linearly increased with elevation but the inorganic C pool varied nonlinearly along the elevation gradient.
In this study, the linear changes in the soil organic C pool along the elevation gradient were positively related to plant diversity and NDVI but negatively related to aboveground biomass density. It is generally found that increases in plant diversity and species richness promote the formation of soil organic C (Gu et al., 2019; Xu et al., 2021; Spohn et al., 2023). This is because SOC is predominantly derived from plant residues (Schmidt et al., 2011). More diverse plant species optimize the complementary use of resources and increase community productivity in areas with lower species richness (Lehmann et al., 2020). Unexpectedly, this study found the negative correlation between soil organic C pool and aboveground biomass density. This could be explained by the shift in vegetation cover type from slow turnover scrub plants (e.g. Krascheninnikovia compacta (Losinsk.) Grubov and Salsola abrotanoides) at the lower elevational range to fast -turnover grassland plants (e.g. S. purpurea and P. bifurca) at the higher elevations. Scrub plants typically have greater standing biomass but a much slower turnover rate of the organs and tissues than herbaceous plants. Moreover, with increases in elevation, temperature decreased and precipitation increased, both of which would favour the preservation of soil organic C. Therefore, changes in both vegetation and climatic conditions led to an increased SOC pool size at the higher elevations (De Deyn et al., 2008).
Climate is often found to be an important abiotic factor affecting the size and stability of the soil C pool (Possinger et al., 2021; Zhang et al., 2024). Our study showed that decreases in temperature and soil water contributed to increased SOC density along the elevational gradient in the dry alpine rangeland. This is contrary to the findings from previous studies in humid environments wherein SOC increases with rising temperature (Chalchissa and Kuris, 2024; Jiang et al., 2024). The discrepancy is mainly because our study area is situated in an extremely arid region, such that water loss through transpiration is much greater than water input from precipitation. When the temperature decreases, the transpiration decreases significantly, which is more conducive to plant growth and soil C accrual (Schmidt et al., 2011). In addition, lower temperatures also significantly inhibit the activity of soil microorganisms, reducing the microbial decomposition of soil organic matter (Sun et al., 2019). In the case of small climate differences brought about by changes in elevation, increases in plant abundance and diversity can lead to increases in plant C input, while the mineralization of microorganisms remains unchanged (Yue et al., 2017).
Apart from the effects of climatic factors and vegetation, previous studies also suggested that soil properties had direct and major effects on soil C stocks (Hemingway et al., 2019). Overall, however, we found that the effect of soil factors on SOC was weak and indirectly through biological factors in the form of plant community structure (Fig. 6a). In this study, we found that soil N level was greater at higher elevations, which favoured the accumulation of SOC (Püspök et al., 2023). This is mainly because an increase in soil N content is often associated with the greater abundance of nitrogen-fixing plants and/or microorganisms and the acceleration of belowground N cycling. Under such conditions, plants generally grow faster and turn over more rapidly, thereby enhancing the inputs of soil organic matter (Reay et al., 2008; Sonam et al., 2016).
In contrast to the clear elevational patterns of changes in SOC density, the SIC density did not display a consistent pattern of elevational variations. Previous studies have proven that soil inorganic C is more influenced by abiotic factors such as topography, soil, and climate (Mi et al., 2008; Li et al., 2024). In this study, the SIC initially declined with elevation up to about 3300 m a.s.l. (Fig. 2) but peaked at about 3400 m a.s.l., at a position where the gentle slope at the lower elevation gave way to a much steeper mountain slope (Fig. 1). According to the sampling positions at each point, the abnormally high SIC value at the foot of the steep slope could be attributed to alluvial deposit and carbonate rock accumulations formed on the uphill slope, reflecting geological and hydrological effects. The correlation analyses revealed that SIC density was mostly related to non-biotic factors such as soil bulk density and soil water. The main constituents of the SIC reservoir are carbonate salts (Zhao et al., 2019). When soil water content is high, CO2 is readily transformed into carbonic acid (H2CO3), carbonate (CO), and bicarbonate (HCO3−), which promotes the dissolution of calcium carbonate and reduces the SIC content (Huber et al., 2019). The greater precipitation at higher elevations may facilitate the leaching of SIC to the deep layer, resulting in a decrease in the surface soil inorganic C pool (Du and Gao, 2020). As a result, the SIC content is higher and more stable in the drier soils at the lower elevations (Ren et al., 2024). Previous studies have shown that SIC may also be affected by biological factors (Ma et al., 2024). Increased plant growth and biological activity enhance root respiratory secretion, resulting in dissolution and loss of SIC (Kuzyakov and Razavi, 2019). In this study, however, SIC density was weakly correlated with root biomass, likely due to the poor soil development in the study area. In the composite SEM analysis, increases in BD and soil led to increased level of soil inorganic C (Fig. 6b). There are two main reasons explaining this phenomenon: the inherent determination of SIC by soil parent materials and the low turnover rate of SIC.
Results from this study show that in the dry and cold alpine rangeland of the Qinghai–Tibet Plateau, soil organic C increases linearly with rising elevation, whereas soil inorganic C does not appear to display a consistent pattern of variations explainable by factors associated with elevation. The relative contribution of SOC to the total soil C pool is greater at higher elevations because of changes in plant communities and climatic conditions promoting soil organic C production and preservation. Overall, inorganic C plays a predominant role in contributing to the soil C pool size in dry alpine rangeland; the distribution of SIC along the elevational gradient is not affected by changes in vegetation and climate factors. Our results show that the soil C pool in the alpine desert region is mainly composed of SIC, differing from findings for the humid regions. The influences of climate, vegetation, and other environmental conditions on soil C pool are mainly achieved by alteration of SOC. Therefore, maintaining ecological stability in cold and dry regions is important for enhancing the overall C sequestration capability of terrestrial ecosystems.
The data provided in this study can be found at Figshare: https://doi.org/10.6084/m9.figshare.29313044 (Zhang, 2025).
The supplement related to this article is available online at https://doi.org/10.5194/bg-22-4123-2025-supplement.
AZ conceived and designed the experiments; QL wrote the manuscript; JY and YZ analysed the data and contributed to the discussion; XL, OJS, and YJ edited and revised the article.
The contact author has declared that none of the authors has any competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
We thank all participants for their contribution to the fieldwork and laboratory analysis.
This paper was edited by Sara Vicca and reviewed by two anonymous referees.
Batjes, N. H.: Soil carbon stocks of Jordan and projected changes upon improved management of croplands, Geoderma, 132, 361–371, https://doi.org/10.1016/j.geoderma.2005.05.013, 2006.
Cai, W. X., Xu, L., Wen, D., Zhou, Z. Y., Li, M. X., Wang, T., and He, N. P.: The carbon sequestration potential of vegetation over the Tibetan Plateau, Renew. Sustain. Energ. Rev., 207, 114937, https://doi.org/10.1016/j.rser.2024.114937, 2025.
Callesen, I., Liski, J., Raulund-Rasmussen, K., Olsson, M. T., Tau-Strand, L., Vesterdal, L., and Westman, C. J.: Soil carbon stores in Nordic well-drained forest soils-Relationships with climate and texture class, Glob. Change Biol., 9, 358–370, https://doi.org/10.1046/j.1365-2486.2003.00587.x, 2003.
Chalchissa, F. B. and Kuris, B. K.: Modelling soil organic carbon dynamics under extreme climate and land use and land cover changes in Western Oromia Regional state, Ethiopia, J. Environ. Manage., 350, 119598, https://doi.org/10.1016/j.jenvman.2023.119598, 2024.
Chang, R. Y., Fu, B. J., Liu, G. H., Wang, S., and Yao, X. L.: The effects of afforestation on soil organic and inorganic carbon: A case study of the Loess Plateau of China, Catena, 95, 145–152, https://doi.org/10.1016/j.catena.2012.02.012, 2012.
Chen, L. T., Jing, X., Flynn, D. F. B., Shi, Y., Kühn, P., Scholten, T., and He, J. S.: Changes of carbon stocks in alpine grassland soils from 2002 to 2011 on the Tibetan Plateau and their climatic causes, Geoderma, 288, 166–174, https://doi.org/10.1016/j.geoderma.2016.11.016, 2017.
Chen, Y., Han, M. G., Yuan, X., Hou, Y. H., Qin, W. K., Zhou, H. K., Zhao, X. Q., Klein, J. A., and Zhu, B. A.: Warming has a minor effect on surface soil organic carbon in alpine meadow ecosystems on the Qinghai-Tibetan Plateau, Glob. Change Biol., 28, 1618–1629, https://doi.org/10.1111/gcb.15984, 2022.
De Deyn, G. B., Cornelissen, J. H. C., and Bardgett, R. D.: Plant functional traits and soil carbon sequestration in contrasting biomes, Ecol. Lett., 11, 516–531, https://doi.org/10.1111/j.1461-0248.2008.01164.x, 2008.
Du, C. J. and Gao, Y. H.: Opposite patterns of soil organic and inorganic carbon along a climate gradient in the alpine steppe of northern Tibetan Plateau, Catena, 186, 104366, https://doi.org/10.1016/j.catena.2019.104366, 2020.
Eswaran, H., Van Den Berg, E., and Reich, P.: Organic carbon in soils of the world, Soil Sci. Soc. Am. J., 57, 192–194, https://doi.org/10.2136/sssaj1993.03615995005700010034x, 1993.
Lehmann, J., Hansel, C. M., Kaiser, C., Kleber, M., Maher, K., Manzoni, S., Nunan, N., Reichstein, M., Schimel, J. P., Torn, M. S. and Wieder, W. R.: Persistence of soil organic carbon caused by functional complexity, Nat. Geosci., 13, 529–534, https://doi.org/10.1038/s41561-020-0612-3, 2020.
Feyissa, A., Raza, S. T., and Cheng, X. L.: Soil carbon stabilization and potential stabilizing mechanisms along elevational gradients in alpine forest and grassland ecosystems of Southwest China, Catena, 229, 107210, https://doi.org/10.1016/j.catena.2023.107210, 2023.
García-Palacios, P., Maestre, F. T., and Milla, R.: Community-aggregated plant traits interact with soil nutrient heterogeneity to determine ecosystem functioning, Plant Soil, 364, 119–129, https://doi.org/10.1007/s11104-012-1349-6, 2013.
Gu, X., Fang, X., Xiang, W. H., Zeng, Y. L., Zhang, S. J., Lie, P. F., Peng, C. H., and Kuzyakov, Y.: Vegetation restoration stimulates soil carbon sequestration and stabilization in a subtropical area of southern China, Catena, 181, 10409810, https://doi.org/10.1016/j.catena.2019.104098, 2019.
Hemingway, J. D., Rothman, D. H., Grant, K. E., Rosengard, S. Z., Eglinton, T. I., Derry, L. A., and Galy, V. V.: Mineral protection regulates long-term global preservation of natural organic carbon, Nature, 570, 228–231, https://doi.org/10.1038/s41586-019-1280-6, 2019.
Huber, D. P., Lohse, K. A., Commendador, A., Joy, S., Aho, K., Finney, B., and Germino, M. J.: Vegetation and precipitation shifts interact to alter organic and inorganic carbon storage in cold desert soils, Ecosphere, 10, e02655, https://doi.org/10.1002/ecs2.2655, 2019.
Jiang, M. D., Li, H. L., Zhang, W., Liu, J. B., and Zhang, Q.: Effects of climate change and grazing on the soil organic carbon stock of alpine wetlands on the Tibetan Plateau from 2000 to 2018, Catena, 238, 107870, https://doi.org/10.1016/j.catena.2024.107870, 2024.
Kuzyakov, Y. and Razavi, B. S.: Rhizosphere size and shape: Temporal dynamics and spatial stationarity, Soil Biol. Biochem., 135, 343–360, https://doi.org/10.1016/j.soilbio.2019.05.011, 2019.
Lal, R.: Digging deeper: A holistic perspective of factors affecting soil organic carbon sequestration in agroecosystems, Glob. Change Biol., 24, 3285–3301, https://doi.org/10.1111/gcb.14054, 2018.
Li, C. L., Cao, Z. Y., Chang, J. J., Zhang, Y., Zhu, G. L., Zong, N., He, Y. T., Zhang, J. J., and He, N. P.: Elevational gradient affect functional fractions of soil organic carbon and aggregates stability in a Tibetan alpine meadow, Catena, 156, 139–148, https://doi.org/10.1016/j.catena.2017.04.007, 2017.
Li, J., Pei, J., Fang, C., Li, B., and Nie, M.: Drought may exacerbate dryland soil inorganic carbon loss under warming climate conditions, Nat. Commun., 15, 617, https://doi.org/10.1038/s41467-024-44895-y, 2024.
Ma, Y. J., Xie, T., and Li, X. Y.: Spatial variation of soil organic carbon in the Qinghai Lake watershed, northeast Qinghai-Tibet Plateau, Catena, 213, 106187, https://doi.org/10.1016/j.catena.2022.106187, 2022.
Ma, Y. Q., Yu, Y., Nan, S. Z., Chai, Y., Xu, W. Y., Qin, Y., Li, X. L., and Bodner, G.: Conversion of SIC to SOC enhances soil carbon sequestration and soil structural stability in alpine ecosystems of the Qinghai-Tibet Plateau, Soil Biol. Biochem., 195, 109452, https://doi.org/10.1016/j.soilbio.2024.109452, 2024.
Mi, N. A., Wang, S., Liu, J., Yu, G., Zhang, W., and Jobbagy, E.: Soil inorganic carbon storage pattern in China, Glob. Change Biol., 14, 2380–2387, https://doi.org/10.1111/j.1365-2486.2008.01642.x, 2008.
Possinger, A. R., Weiglein, T. L., Bowman, M. M., Gallo, A. C., Hatten, J. A., Heckman, K. A., Matosziuk, L. M., Nave, L. E., SanClements, M. D., Swanston, C. W., and Strahm, B. D.: Climate Effects on Subsoil Carbon Loss Mediated by Soil Chemistry, Environ. Sci. Technol., 55, 16224–16235, https://doi.org/10.1021/acs.est.1c04909, 2021.
Püspök, J. F., Zhao, S., Calma, A. D., Vourlitis, G. L., Allison, S. D., Aronson, E. L., Schimel, J. P., Hanan, E. J., and Homyak, P. M.: Effects of experimental nitrogen deposition on soil organic carbon storage in Southern California drylands, Glob. Change Biol., 29, 1660–1679, https://doi.org/10.1111/gcb.16563, 2023.
Reay, D. S., Dentener, F., Smith, P., Grace, J., and Feely, R. A.: Global nitrogen deposition and carbon sinks, Nat. Geosci., 1, 430–437, https://doi.org/10.1038/ngeo230, 2008.
Ren, Z. B., Li, C. J., Fu, B. J., Wang, S., and Stringer, L. C.: Effects of aridification on soil total carbon pools in China's drylands, Glob. Change Biol., 30, e17091, https://doi.org/10.1111/gcb.17091, 2024.
Rodeghiero, M. and Cescatti, A.: Main determinants of forest soil respiration along an elevation/temperature gradient in the Italian Alps, Glob. Change Biol., 11, 1024–1041, https://doi.org/10.1111/j.1365-2486.2005.00963.x, 2005.
Schmidt, M. W. I., Torn, M. S., Abiven, S., Dittmar, T., Guggenberger, G., Janssens, I. A., Kleber, M., Kögel-Knabner, I., Lehmann, J., Manning, D. A. C., Nannipieri, P., Rasse, D. P., Weiner, S., and Trumbore, S. E.: Persistence of soil organic matter as an ecosystem property, Nature, 478, 49–56, https://doi.org/10.1038/nature10386, 2011.
Schuur, E. A., Chadwick, O. A., and Matson, P. A.: Carbon cycling and soil carbon storage in mesic to wet Hawaiian montane forests, Ecology, 82, 3182–3196, https://doi.org/10.1890/0012-9658(2001)082[3182:CCASCS]2.0.CO;2, 2001.
Sonam, T., Balwant, S., Claudia, K., and Mark, A.: Soil carbon and nitrogen stocks in forests along an altitudinal gradient in the eastern Himalayas and a meta-analysis of global data, Glob. Change Biol., 22, 2255–2268, https://doi.org/10.1111/gcb.13234, 2016.
Spohn, M., Bagchi, S., Biederman, L. A., Borer, E. T., Bråthen, K. A., Bugalho, M. N., Caldeira, M. C., Catford, J. A., Collins, S. L., Eisenhauer, N., and Hagenah, N.: The positive effect of plant diversity on soil carbon depends on climate, Nat. Commun., 14, 6624, https://doi.org/10.1038/s41467-023-42340-0, 2023.
Sun, O. J., Campbell, J., Law, B. E., and Wolf, V.: Dynamics of carbon stocks in soils and detritus across chronosequences of different forest types in the Pacific Northwest, USA, Glob. Change Biol., 10, 1470–1481, https://doi.org/10.1111/j.1365-2486.2004.00829.x, 2004.
Sun, X., Tang, Z., Ryan, M. G., You, Y., and Sun, O. J.: Changes in soil organic carbon contents and fractionations of forests along a climatic gradient in China, Forest Ecosyst., 6, 1–12, https://doi.org/10.1186/s40663-019-0161-7, 2019.
Tian, P., Liu, S., Zhao, X., Sun, Z., Yao, X., Niu, S., Crowther, T. W., and Wang, Q.: Past climate conditions predict the influence of nitrogen enrichment on the temperature sensitivity of soil respiration, Commun. Earth d Environ., 2, 251, https://doi.org/10.1038/s43247-021-00324-2, 2021.
Tiemann, L. K. and Billings, S. A.: Changes in variability of soil moisture alter microbial community C and N resource use, Soil Biol. Biochem., 43, 1837–1847, https://doi.org/10.1016/j.soilbio.2011.04.020, 2011.
Torn, M. S., Trumbore, S. E., Chadwick, O. A., Vitousek, P. M., and Hendricks, D. M.: Mineral control of soil organic carbon storage and turnover, Nature, 389, 170–173, https://doi.org/10.1038/38260, 1997.
Trumbore, S. E.: Potential responses of soil organic carbon to global environmental change, P. Natl. Acad. Sci. USA, 94, 8284–8291, https://doi.org/10.1073/pnas.94.16.8284, 1997.
Wang, M. M., Zhang, S., Guo, X. W., Xiao, L. J., Yang, Y. H., Luo, Y. Q., Mishra, U., and Luo, Z. K.: Responses of soil organic carbon to climate extremes under warming across global biomes, Nat. Clim. Change, 14, 98–105, https://doi.org/10.1038/s41558-023-01874-3, 2024.
Wang, Z. W., Huang, L. M., and Shao, M. A.: Spatial variations and influencing factors of soil organic carbon under different land use types in the alpine region of Qinghai-Tibet Plateau, Catena, 220, 106706, https://doi.org/10.1016/j.catena.2022.106706, 2023.
Whittaker, R. H. and Niering, W. A.: Vegetation of the Santa Catalina Mountains, Arizona: a gradient analysis of the south slope, Ecology, 46, 429–452, https://doi.org/10.2307/1934875, 1965.
Xu, M., Li, X. L., Kuyper, T. W., Xu, M., Li, X. L., and Zhang, J. L.: High microbial diversity stabilizes the responses of soil organic carbon decomposition to warming in the subsoil on the Tibetan Plateau, Glob. Change Biol., 27, 2061–2075, https://doi.org/10.1111/gcb.15553, 2021.
Yue, K., Fornara, D. A., Yang, W. Q., Peng, Y., Peng, C. H., Liu, Z. L., and Wu, F. Z.: Influence of multiple global change drivers on terrestrial carbon storage: additive effects are common, Ecol. Lett., 20, 663–672, https://doi.org/10.1111/ele.12767, 2017.
Zhang, A.: Changes in soil organic and inorganic carbon with elevation in a1dry alpine rangeland of northern Qinghai-Tibetan Plateau, Figshare [data set], https://doi.org/10.6084/m9.figshare.29313044, 2025.
Zhang, A., Li, X., Wu, S., Li, L., Jiang, Y., Wang, R., Ahmed, Z., Zeng, F., Lin, L., and Li, L.: Spatial pattern of C : N : P stoichiometry characteristics of alpine grassland in the Altunshan Nature Reserve at North Qinghai-Tibet Plateau, Catena, 207, 105691, https://doi.org/10.1016/j.catena.2021.105691, 2021.
Zhang, Y. X., Guo, X. W., Chen, L. X., Kuzyakov, Y., Wang, R. Z., Zhang, H. Y., Han, X. G., Jiang, Y., and Sun, O. J.: Global pattern of organic carbon pools in forest soils, Glob. Change Biol., 30, 17386, https://doi.org/10.1111/gcb.17386, 2024.
Zhao, W., Zhang, R., Cao, H., and Tan, W. F.: Factor contribution to soil organic and inorganic carbon accumulation in the Loess Plateau: Structural equation modeling, Geoderma, 352, 116–125, https://doi.org/10.1016/j.geoderma.2019.06.005, 2019.