the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 27 Feb 2026
| 27 Feb 2026
Consequences of the 2022 drought on nutrient dynamics in forest soil solutions of a declining spruce plot in the Strengbach catchment (Vosges Mountains, France)
Adrien Saphy
Emilie Beaulieu
Benjamin Belfort
Marie-Noëlle Pons
Marie-Claire Pierret
The year 2022 was the hottest and driest year ever recorded in France, including within the Strengbach catchment, a Critical Zone Observatory (http://ohge.unistra.fr, last access: 18 September 2025) located in a forested watershed of the Vosges Mountains and characterized by declining Norway spruce (Picea abies) stands. During and following the summer drought of 2022, an unusual chemical signature was detected in soil solutions, marked by elevated concentrations of K+, Ca2+, Mg2+, NO, NH, Al3+, and Dissolved Organic Carbon (DOC) with significant variations of fluorescence indices (HIX, BIX and FI). The long-term monitoring of soil solution chemistry exhibited that drought affects soil nutrient dynamics at the soil-water-plant interface through different mechanisms: (1) lower mineral dissolution, (2) reduced plant nutrient uptake, (3) increased concentrations in throughfall, (4) organic matter accumulation during the dry period and subsequent release upon rewetting, (5) changes in dissolved organic matter quality suggesting soil microbial stress, (6) disruption of the nitrogen cycle, with ammonium accumulation during drought followed by intense nitrification after rainfall resumed, and (7) acidification of the soil solution, enhancing the desorption of both nutrient cations and toxic Al3+. The decline in soil fertility during and after drought is especially concerning in forest ecosystems already subject to nutrient deficiency, such as those in the Strengbach catchment. Understanding these drought-induced biogeochemical disturbances is essential for predicting ecosystem responses to extreme climatic events, whose intensity and frequency are expected to increase in the Vosges Mountains under ongoing climate change.
- Article
(6575 KB) - Full-text XML
-
Supplement
(726 KB) - BibTeX
- EndNote




The year 2022 was the hottest ever recorded in mainland France to date (since 1900), +2.9 °C compared with the mean temperature during 1900–1930 period, with a sequence of heatwaves from May onwards (Faranda et al., 2023; Sorel et al., 2023). With a major annual precipitation deficit of 25 % in 2022, the months of May and July were the driest on record in France (with deficits of nearly 60 % and 85 % respectively compared to the normals based on data since 1959), giving rise to a severe meteorological summer drought that spread across Europe (Sorel et al., 2023; Copernicus Climate Change Service (C3S), 2023). Climate change projections for the end of the century remain very worrying, as simulations predict for France: (i) an increase in the number of consecutive days without precipitation of around +5 and +10 d respectively for the scenarios RCP4.5 and RCP8.5, and (ii) an increase in the frequency of heat waves in summer of +10 to +15 d for the RCP4.5 scenario and approximately double for RCP8.5. (Soubeyroux et al., 2021). Droughts have a major impact on the functioning of forest ecosystems and, whether or not combined with other disturbances, such as fires or insect attacks, they can cause forest dieback (Anderegg et al., 2013; Senf et al., 2020; Gharun et al., 2024; Knutzen et al., 2025).
Forest dieback is increasing in France, as shown by the latest report from the French National Institute of Geographic and Forest Information (Institut national de l’information géographique et forestière (IGN), 2024), with mortality rising from 7.4 Mm3 yr−1 for the period 2005–2013 to 15.2 Mm3 yr−1 for the period 2014–2022 (with statistical uncertainty of around 0.6 Mm3 yr−1) (Institut national de l’information géographique et forestière (IGN), 2024). Drought has direct physiological consequences on trees and can lead to direct mortality by xylem embolism for instance (Bréda et al., 2006; Rodríguez-Calcerrada et al., 2017; Wagner et al., 2023). The 2022 drought had important consequences for French forests, notably mega-fires that burned 58 275 ha of forest in 2022 (San-Miguel-Ayanz et al., 2023). In addition, global warming and frequent droughts have accelerated the spread and intensity of insect attacks, as documented in U.S. forests (Weed et al., 2013; Vose et al., 2016; Frank, 2021) and in Europe (Hartmann et al., 2022). In western Europe, a recent initiative to address our lack of field knowledge regarding the ecological processes underlying biotic forest disturbances has been the creation of the Database of European Forest Insect and Disease Disturbances (DEFID2) (Forzieri et al., 2023). Spruces are trees from boreal regions. Out of its natural localisation, spruces can be more affected by droughts, leading to an increase in the tree's vulnerability to parasites, such as the bark beetle (Ips typographus), which devastates Vosges spruce forests (Saintonge, 2022; Gomez et al., 2023; Knutzen et al., 2025). However, all these disturbances affect forests and therefore the carbon sink provided by the soils and tree photosynthetic activity. While forest ecosystems represent the largest terrestrial carbon stock, estimated at 662 gigatonnes worldwide, a decrease of 0.9 % has already been estimated between 1990 and 2020 (FAO, 2020). Forest dieback thus threatens the mitigation role of forests in climate change, and foreshadows negative feedback (Ciais et al., 2005; Anderegg et al., 2020; Quirion et al., 2021).
Another factor of tree vulnerability to drought may be a decrease in nutrient availability (Lévesque et al., 2016; Gessler et al., 2017; Schmied et al., 2023). How drought affects nutrient dynamics in forest soil is a high concern issue for the prediction of the complex responses of forest ecosystems to drought, in a warming world. Nitrogen is an essential plant nutrient, and the consequences of drought on the nitrogen cycle are documented (Lamersdorf et al., 1998; Muhr et al., 2008; Deng et al., 2021; Krüger et al., 2021; Winter et al., 2023, 2025). However, a lack of information persists about the dynamics of all the other nutrients, such as major cations K+, Ca2+ and Mg2+.
To better understand and predict drought-related biogeochemical disturbance, some studies have been carried out on the impact of water shortage on nutrient cycles in forests (Sardans et al., 2008; Touche at al., 2022) and in particular in spruce forests (Dambrine et al., 1993; Nilsen, 1995). These authors observed a decrease in K concentration in biomass, suggesting a reduction of K availability for trees during water deficit. Moreover, exceptional K concentrations in soil solutions at 30 cm depth following natural drought events were documented, such as in 2012, in a Canadian boreal forest (Houle et al., 2016). This nutrient export flux was explained by a high K concentration coming from throughfall and the higher mobility of K compared with other major cations through the soil profile. To assess the impact of drought events on forest and soil fertility, the composition of soil solutions can be used as an indicator of nutrient availability. However, studies addressing the impact of natural drought on mineral nutrient dynamics in soil solutions remain scarce and require multiple observations to be better understood.
The hydrogeochemical environmental observatory of the Strengbach catchment in the Vosges Mountains (France) is a critical zone observatory belonging to the French Network of Critical Zone Observatories (OZCAR). The continuous recording of numerous variables since 1986 makes it a key site for observing the consequences of global changes on environmental parameters (Pierret et al., 2018; Strohmenger et al., 2022). An experiment of water shortage was conducted on a spruce plot in the Strengbach catchment in 1990 (Dambrine et al., 1993; Lu et al., 1995). These studies reported reduced sap flow as well as lower concentration of nutrients Ca, Mg and K in the sap during periods of water deficit. This indicates that tree nutrient uptake is reduced during a water deficit field simulation. However, this reduction may also result from disturbances in various biogeochemical processes that control nutrient availability in the soil, such as mineral weathering, cation exchange, or organic matter degradation. In addition, a severe drought such as the one experienced in 2022 is far more complex than an experimental water deficit, as high temperatures influence all the biogeochemical processes. In 2022, long-term monitoring of soil solution chemistry captured an unprecedented severe drought event, providing a unique opportunity to elucidate its impact on the biogeochemical cycling of major elements and organic matter. Such climatic extremes may be a high cause of concern for nutrient-deficient soils of the Strengbach catchment (Probst et al., 1992; Dambrine et al., 1998; Oursin et al., 2023).
The main objectives of this study were to evaluate the potential impacts of drought 2022 on biogeochemical cycles of major elements, on organic matter and on the fertility of forest soils. Soil solutions were sampled under a spruce plot during the period 2015–2023 and the concentrations of major cations (K+, Na+, Ca2+, Mg2+), Al, Si, as well as nitrogen in various forms (NH, NO) and Dissolved Organic Matter (DOC) were measured. To better understand DOC dynamics, fluorescence of soil solution was monitored between 2020 and 2023. The combined analysis of mineral nutrients and dissolved organic matter, supported by hydrological modelling, provides a global view of the biogeochemical consequences of drought on the chemistry of soil solutions and, more generally, on soil fertility.
2.1 Site presentation
The Strengbach catchment, located in the municipality of Aubure (Haut-Rhin, France) in the Vosges Mountains, has an area of 0.8 km2 and an elevation between 850 and 1150 m. This catchment is mainly covered by acid brown soil and ochreous podzolic soil (Fig. 1) on Ca-poor granitic bedrock (Pierret et al., 2018). The forest covers 90 % of the catchment with 80 % planted with a monospecific spruce plantation (Picea abies) and the rest with beech (Fagus sylvatica) (Fichter et al., 1997; Pierret et al., 2018).
Meteorological, hydrological and geochemical variables have been monitored since 1986 in the Strengbach catchment by the Observatoire hydro-geochimique de l'environnement (OHGE; https://ohge.unistra.fr, last access: 18 September 2025). The creation of the observatory (with the installation of the first equipment) and the first studies were performed in order to understand the impact of acid rain on forested ecosystems (Probst et al., 1990, 1992, 1995; Pierret et al., 2019). Many studies are still being conducted on environmental issues concerning forest mountainous ecosystems and particularly about forest decline and nutrient deficiency (Fichter et al., 1997; Poszwa et al., 2003; Cenki-Tok et al., 2009; Brioschi et al., 2013; Beaulieu et al., 2020; Oursin et al., 2023).
The climate is oceanic mountainous with an annual mean temperature of 6.17 °C, and a mean precipitation of about 101 mm per month during the time period 1987–2024. The annual precipitation (over a calendar year) has varied between 814 mm yr−1 (2023) and 1568 mm yr−1 (2007) with an average of 1214 mm yr−1. Outlet water flows vary between 464 and 1132 with an average of 732 mm yr−1. Historically, rainfall was uniformly distributed throughout the year. But climate change consequences were observed at the local scale in the OHGE, with wetter summers and winters, and drier autumns when comparing before and after 2006 (Strohmenger et al., 2022). The OHGE databank is a key tool to understand environmental change consequences (Pierret et al., 2018; Strohmenger et al., 2022).
2.2 Stand presentation
The plot (VP plot, Fig. 1) is a monospecific spruce (Picea abies) plot, planted in the 1900s, which has been declining for several decades due to different reasons: nutrient-poor soils amplified by acid rain until the 1980s (Probst et al., 1990, 1992, 1995), several recent violent storms, successive bark beetle attacks as well as intense drought episodes. The site, where trees were still alive, was instrumented from 2015 to 2023. Soil solutions were collected with 40×40 cm zero-tension lysimetric plates installed at four depths (5, 10, 30, 60 cm) in 1990 and sampled every six weeks during the period 2015–2023. Throughfall (fraction of precipitation that reaches the ground after passing through the canopy) samples were collected with two gutters (2.0×0.2 m) every two weeks (Pierret et al., 2018). The minimum volume of water needed for analyses is 25 mL, volumes of soil solution samples collected are given in Supplement (Table S1).
The studied soil profile (Fig. 1) is a Cambisol (Alocrisol in the French taxonomy) on granitic bedrock. We can distinguish two litter horizons. The upper litter (OL) which is composed of old needles, herbaceous roots and pieces of wood, is 5 cm thick. The lower humic horizon (OH-OF) is 6 cm thick and is black-colored with a smooth texture. The dark brown/grey horizon A of 5–6 cm is unequally distributed on the profile with wavy transitions. It is followed by a transition layer of 5 cm. At 17 cm depth, a brown/red mineral horizon B is found with a lumpy texture. This horizon is homogeneous and very sandy with centimetric gravel.
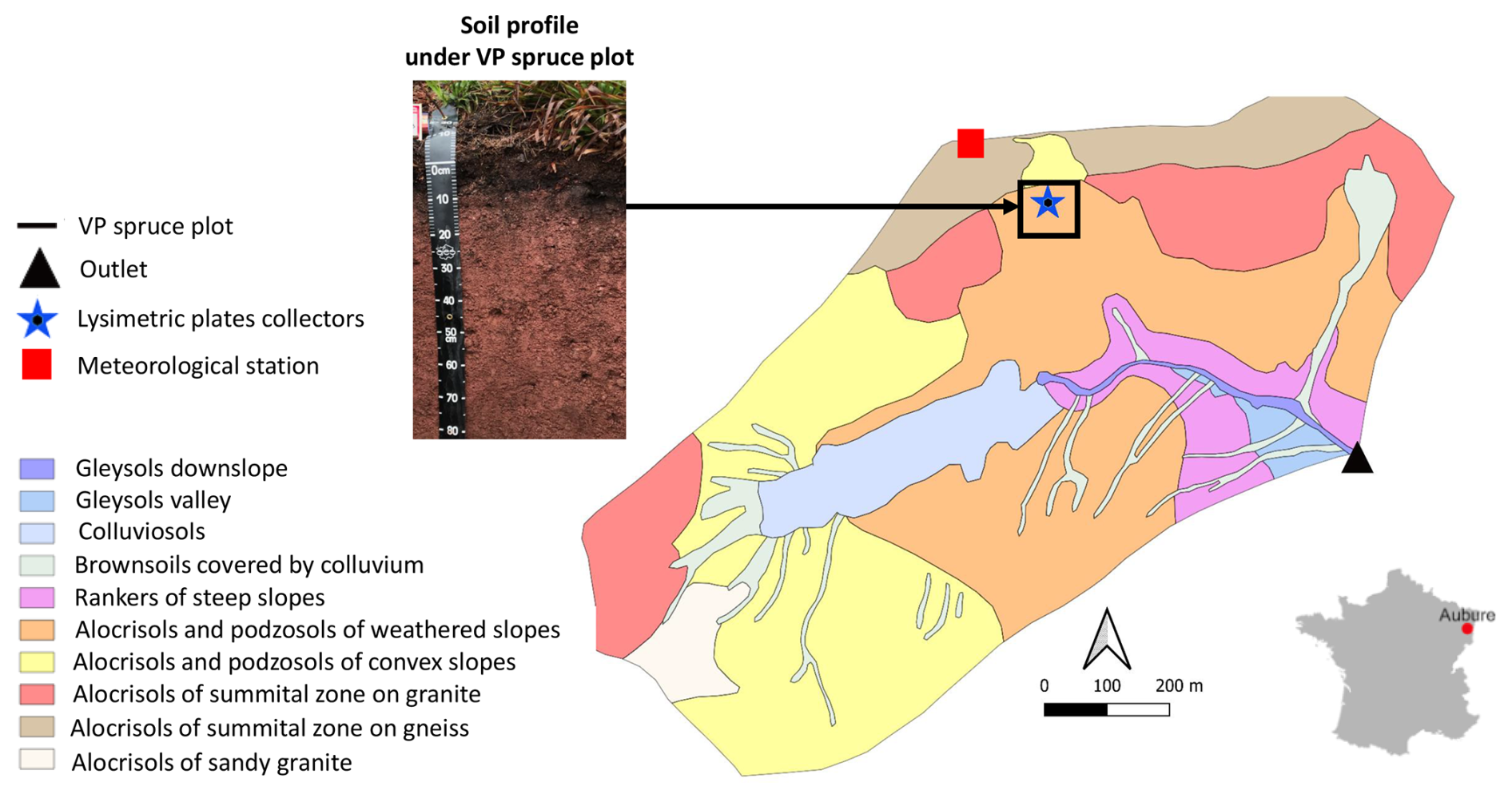
Figure 1Soil map of the Strengbach watershed with different monitoring equipment and stations, with the location of the studied soil profile of the VP spruce plot.
2.3 Chemical analysis of soil solution
Soil solutions were collected in clean polyethylene (HDPE) containers sampled from the field and filtered with cellulose acetate filters 0.45 µm pore diameter membrane Merck-Millipore on a PTFE filtration unit. Conductivity and pH were measured with HQ440d Hach multimeter. The dissolved organic carbon (DOC) was determined using a carbon analyser (Shimadzu TOC-VCPH) with an uncertainty of 2 % to 3.5 %. The accuracy of the analysis was assessed by regular measurement of PERADE-17 river water CRM (Environment and Climate Change Canada matrix reference materials). K+, Na+, Ca2+, Mg2+, NH and NO concentrations were determined by ion chromatography on a Thermo Scientific Dionex ICS-5000+. Al and Si concentrations were measured by ICP-AES on a Thermo Scientific iCAP 6000 SERIES. PERADE-17 and SLRS-6 water CRMs (certified by Environment and Climate Change Canada) were used as standards and the analytical uncertainty for ions concentrations is ± 2 %.
To characterise the dissolved organic matter UV-fluorescence of soil solution was measured. Fluorescence spectra were obtained on a Hitachi F-2500 spectrofluorometer equipped with a Xenon lamp, using FL Solution 2.0 software and a 1 cm × 1 cm (3.5 mL) quartz cuvette. Emission spectra were collected for three excitation wavelengths (λex= 254, 310 and 370 nm) with a step of 1 nm and slits of 2.5 nm. The humification index HIX was calculated on emission spectra for λex= 254 nm as the ratio of the sum of the fluorescence intensities (F) between 435 and 480 nm over the sum of the fluorescence intensities between 300 and 345 nm (Zsolnay et al., 1999), (Eq. 1). HIX quantifies aromatic molecules in solutions and is also qualitative of the complexity of aromatics (Zsolnay et al., 1999; Serène et al., 2025). The index BIX was calculated as the ratio of fluorescence intensity emitted at 380 nm over 430 nm for λex= 310 nm (Huguet et al., 2009) (Eq. 2). BIX is a proxy of biological activity in aquatic environment (Parlanti et al., 2000; Huguet et al., 2009). The index FI was calculated as the ratio of fluorescence intensities at 450 and 500 nm for λex= 370 nm (Eq. 3) (McKnight et al., 2001). FI indicates the origin of DOM and distinguishes two endmembers: terrestrially and microbially derived DOM.
2.4 Hydrological soil modelling
Drought events can create variations in soil water content. To evaluate the changes in water availability in the soil profile induced by rainfall decrease, an in-house daily water balance calculation code was applied to a soil profile under spruces. The BILHYDAY code, whose structure is inspired by the model BILJOU developed by Granier et al. (1999), takes meteorological data as inputs with stand parameters (leaf area index, soil porosity and residual water content). Based on a conceptual approach, the model operates on a daily time step to describe the hydrological processes of the soil–tree–atmosphere continuum. Potential evapotranspiration is estimated following the Penman formulation. Precipitation feeds a canopy reservoir that simulates interception up to a threshold capacity, while transpiration is represented as a function of potential evapotranspiration, soil water storage, and leaf area index. Transpiration is distributed among soil layers according to the root density profile. Soil evaporation is estimated from the incident energy (induced from direct radiation) and an extinction coefficient. Five soil layers are represented as cascading reservoirs that drain sequentially from top to bottom according to their field capacity. The soil is divided into 5 layers of 5-5-20-30-40 cm thickness (from the surface). This discretization of the vertical soil profile is specifically based on the installed instrumentation. It includes a top layer (0 to 5 cm), which is particularly sensitive to exchanges with the atmosphere, and a bottom layer (60 to 100 cm) that drains toward the deeper soil zone, with three intermediate layers (5 to 10 cm, 10 to 30 cm, and 30 to 60 cm) corresponding to the layers at which soil solution samples are collected. The uppermost layer is supplied by non-intercepted precipitation and is also subject to evaporation losses. Root water uptake associated with transpiration is distributed among the different soil layers according to the root density profile. The BILHYDAY model is used as a diagnostic tool to provide complementary hydrological information supporting the interpretation of geochemical measurements of soil solutions. The model structure and parameterisation are based on previous developments and applications, notably those described by Biron (1994), during which the different components of the soil water balance were assessed. In the present study, this existing parameterisation was retained for soil properties, and the leaf area index (LAI) was updated to account for changes in forest stand development. Such an approach has recently been used for the soil hydrodynamic characterization of two other plots in the Strengbach catchment; BILHYDAY was run in combination with a physically based hydrological model coupled with an inverse parameter estimation module (Belfort et al., 2025). To address potential concerns regarding the realism of the simulated soil moisture dynamics, additional soil water content measurements collected at locations close to the soil solution sampling site are provided in Supplement (Fig. S2). Although these measurements are not strictly co-located with the simulated soil profile, they exhibit temporal dynamics consistent with the model outputs, thereby supporting the coherence of the simulated soil moisture values used in this study. The relative extractable soil water was calculated similarly to Granier et al. (1999) and can be used to assess water stress. Although the model does not simulate photosynthesis by integrating a dynamic stomatal conductance, transpiration is nevertheless adjusted according to soil water availability, with regulation occurring below a threshold of 0.4, as proposed by Bréda et al. (2006).
2.5 Statistical analysis
To find outlier points, we used a robust method based on the calculation of the first and the third quartiles (Q1 and Q3) and the interquartile range (IQR = Q3−Q1). It consists of determining an interval with the lower bound ( IQR) and the upper bound ( IQR), whereby points outside this interval are considered outliers. The value of 3 was chosen to identify only extreme outliers (Schwertman et al., 2004; Grunsky, 2010). A box plot representation allows outliers to be visualized as points outside this interval (Supplement Fig. S3).
Non-metric multidimensional scaling (NMDS) was performed to identify atypical chemical signatures within the long-term monitoring dataset. NMDS is effective for highlighting atypical observations in long-term monitoring and can reveal the impact of extreme events on soil when many parameters are measured (Knight et al., 2024), or the consequences of extreme meteorological events on hydrochemistry (Setia et al., 2021). The analysis was conducted using the metaMDS function in the `vegan' library in R (Oksanen et al., 2001) on the dataset of chemical measurements (pH, conductivity, Na+, K+, Mg2+, Ca2+, Al, Si, NH, NO, DOC) for the period 2015–2023. To evaluate depth-specific anomalies, the data were partitioned by sampling depth (5, 10, 30, and 60 cm). Plotting the coordinates of different soil solution samples highlighted atypical points. Finally, the scores assigned to the chemical parameters were analyzed to determine which variables primarily drove the observed anomalies and distinguished the 2022 drought samples from the long-term baseline.
3.1 Meteorological drought and soil water deficit: insights from hydrological modelling
On the OHGE site, considering the hydrological years over the recorded period from 1987–2023 (year running Y from 1 October to 31 September of year Y+1), 2021/2022 (including the 2022 drought) was the third driest year on record (after 1990/1991 and 1995/1996) with a cumulated precipitation amount of 970 mm (−20 % compared to the average value over the period), the fourth warmest year with an average annual temperature of 7.47 °C (+20 % compared to the average for the period – just after 2019/2020, 2023/2024 and 2022/2023), and with the second highest number of days with an average daily temperature above 18 °C, i.e. 39 d compared to an average of 17 d. Among the years with the least precipitation, 2021/2022 was the warmest year, which in combination makes 2021/2022 the most drought-affected year.
Meteorological drought is generally defined as a prolonged period during which precipitation is significantly below normal or expected levels for a given location and season, resulting in a deficit of moisture relative to climatic norms (Wilhite and Glantz, 1985). The longest period of summer meteorological drought (defined here as consecutive days without rainfall) observed between 1987 and 2024 was 19 d in 2018, followed by 17 d in 2022, 2013 and 1990. Nevertheless, the summer of 2022 was exceptional because this first drought period from 2 July to 19 July was very quickly followed by a second period of 24 d during which daily rainfall did not exceed 0.7 mm (between 21 July and 13 August 2022; see the red band in Fig. 2A). Notably, a rainfall event of 8.9 mm on 20 July 2022 separated these two meteorological drought events (Fig. 2A). The summer months of July and August, with only 61 mm of rainfall, were the driest ever recorded at the site (Fig. S1), representing a decrease of nearly 70 % relative to normal conditions.The particularly dry month of July (10.2 mm cumulated; Fig. S1; Fig. 2A) was amplified by the previous dry March and May months with respectively 33 and 36 mm cumulated, compared with the 102 mm/month usually observed in the Strengbach watershed.
To assess the impact of rainfall deficit on soil water content, BILHYDAY was used with meteorological data. The water content of the three intermediate layers (denoted by their centers at 7.5, 20 and 45 cm) and the relative extractable soil water (REW) during the 2021–2022 hydrological year are depicted in Fig. 2B–C. The simulations showed that the water deficit began in the middle of May in the 7.5 cm depth layer, and the few rainy events in the first half and last week of June did not prevent it from falling to a minimum value of 0.14 in mid-July. The 20 cm depth layer experienced a gradual drying out from May to 22 June; after the rains at the beginning of July, the minimum water content was recorded on 30 July reflecting a significant water deficit (Fig. 2B). The water content remained constant and low in these layers until 16 August 2022. The water content decrease was slower in the 45 cm depth layer with a minimum on 14 August 2022. Following the return of the rainy season, there was an increase in soil water content during the second half of August, particularly in the second layer, while recharge was slower for deeper layers. This was followed by a rapid rise in September, coinciding with the return of heavier rains, resulting in a value of 0.42 and 0.35 at 7.5 and 20 cm depth respectively (Fig. 2B). During the summer of 2022, a total of 25 d of water deficit occurred with the relative extractable soil water below the threshold of 0.4 (Fig. 2C). Trees transpiration increased during spring and summer, reaching its maximum (2.5 mm d−1) in the middle of June and then sharply decreased (Fig. 2C). During the water stress period from mid-July to mid-August, tree transpiration reached a very low value due to stomatal closure or downregulation, and this continued until rain returned.
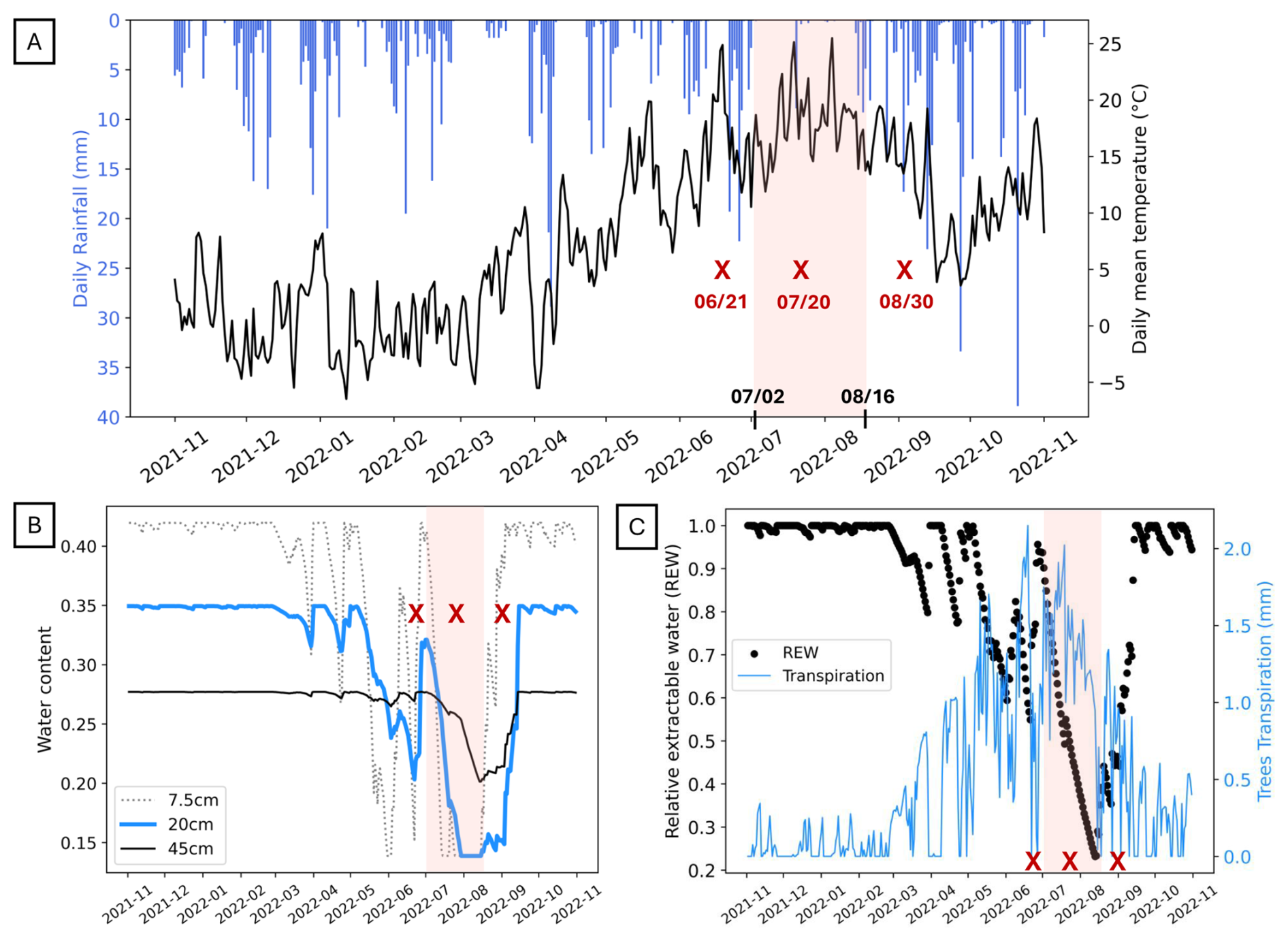
Figure 2(A) Daily rainfall (mm) in blue bar plot and daily mean temperature (°C); (B) simulated soil water content at 7.5 cm depth in dotted line, 20 cm in blue line, 45 cm in black line; (C) simulated relative extractable water (REW) in black points and simulated trees transpiration in blue line. All data are plotted during the period from November 2021 to November 2022. The red background indicates the meteorological drought period, and the red crosses indicate the date of three field soil solution sampling corresponding to the drought period (21 June 2022, 20 July 2022 and 30 August 2022).
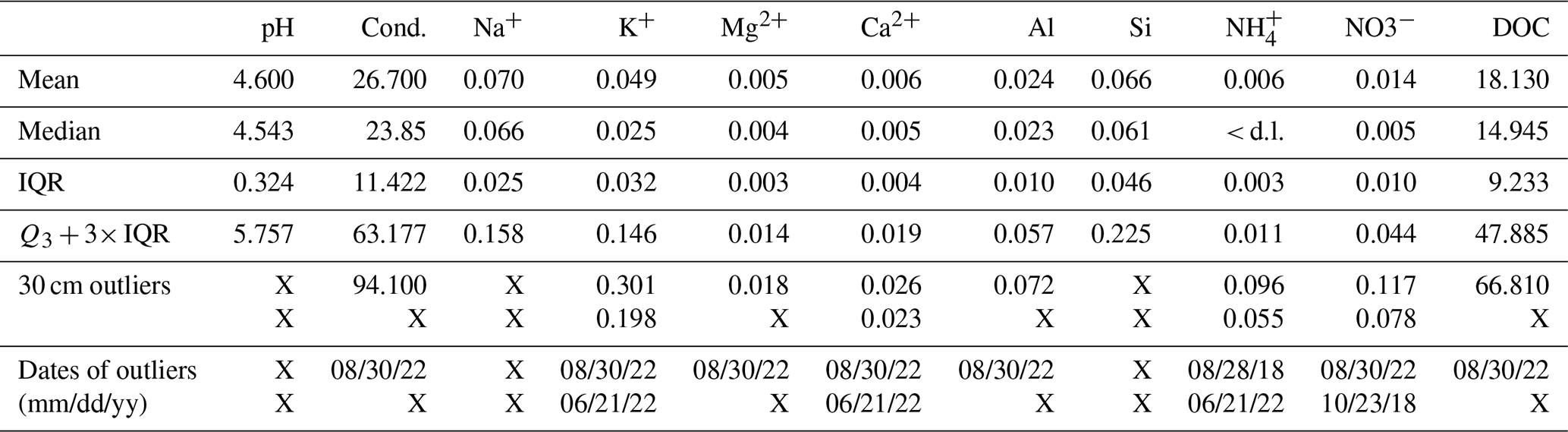
Table 1Mean and median of pH, Conductivity (Cond. in µS cm−1), Dissolved Organic Carbon (DOC in ppm-C), and concentrations (mmol L−1) in ions (Na+, K+, Mg2+, Ca2+, Al, Si, NH and NO) for the 2015–2023 period in the soil solutions at 30 cm depth. Q3 is 75th percentile, IQR the interquartile range and ( IQR) the upper bound above which points are considered outliers. Values of outliers at 30 cm depth are given in the fifth line with the corresponding date in the sixth line. “X” means that no outlier is found at 30 cm depth during the 2015–2023 period. “< d.l.” means below detection limit.
3.2 Chemical composition of field soil solutions
The field monitoring of soil solution concentrations in the Strengbach catchment allowed for the observation of long-term variations of major cations concentrations through the 2015–2023 period. The time series exhibited annual seasonality in the chemical signal, as well as significant anomalies during the exceptionally severe drought of summer 2022 with numerous outlier concentrations (Figs. 3 and S2, Table 1).
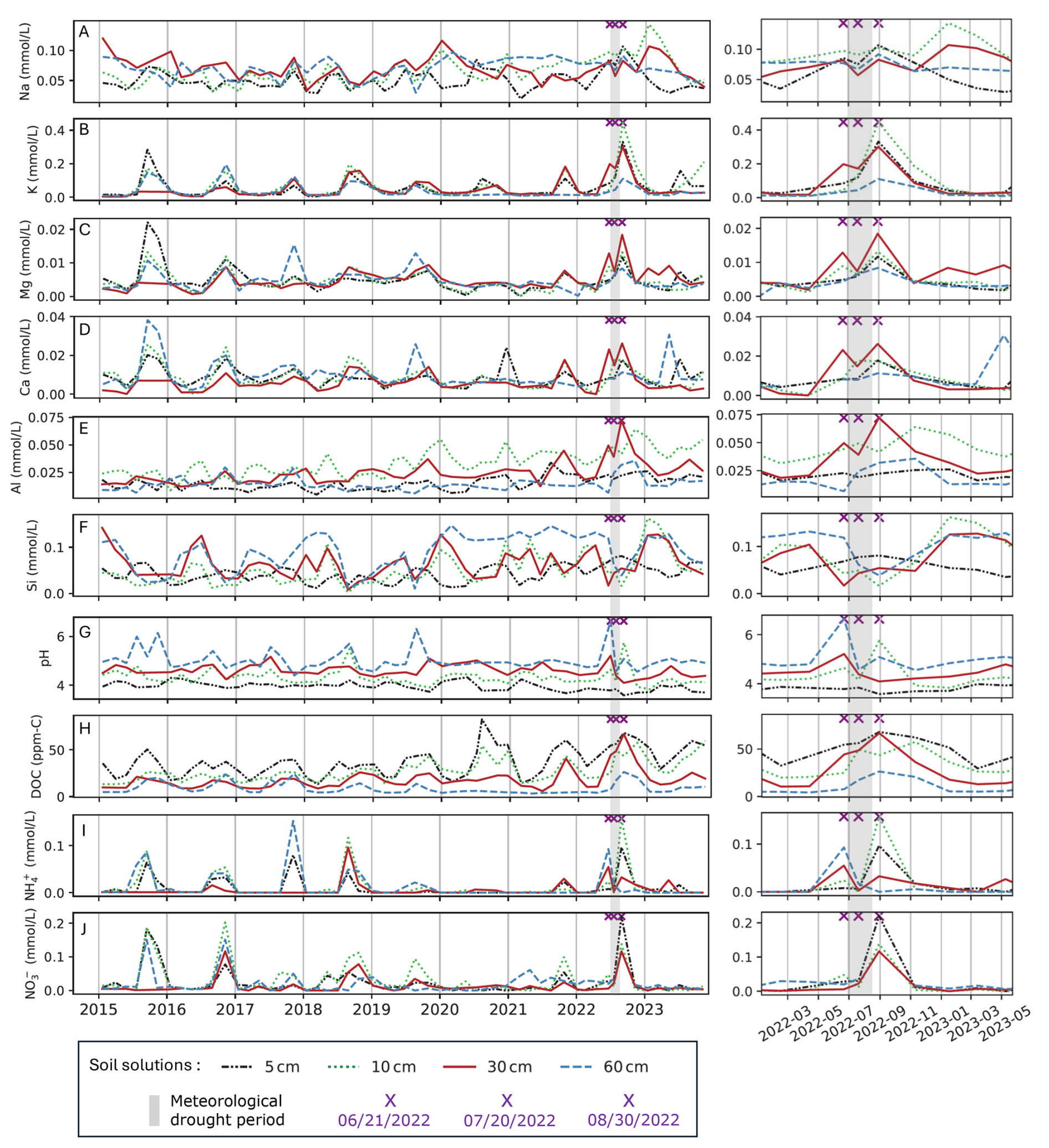
Figure 3Soil solution cations concentrations (mmol L−1) of Na+ (A), K+ (B), Mg2+ (C), Ca2+ (D), Al (E), Si (F), pH (G), Dissolved Organic Carbon DOC (ppm-C) (H), NH (I) and NO (J) at four different depths (5, 10, 30 and 60 cm) for the 2015–2023 time period. On the right side, a zoom on the 2022 hydrological year is plotted. The 2022 meteorological drought period is represented by a grey band. The purple crosses indicate the date of three field soil solution sampling corresponding to the drought period (21 June 2022, 20 July 2022 and 30 August 2022).
The most extreme chemical anomaly was observed at 30 cm depth on 30 August 2022 with an exceptionally high conductivity (94.1 µS cm−1) and high DOC (66.81 ppm-C). These values were well above the inter-annual variability and were the highest ever registered at this depth, considered as outliers because they exceed IQR (Table 1; Fig. S3). The soil solution sample of 30 August integrated the period from 20 July to 30 August coinciding with the return of rainfall after the long dry period (Fig. 2). This high conductivity corresponded to peaks of concentration of many ions such as K+, H+, Ca2+, Mg2+, Al, Mn2+, Fe3+, NO, NH, Cl− except for Na+, SO and Si. These peaks were specially marked at 30 cm (Fig. 3) and were confirmed as outlier concentrations for K+, Ca2+, Mg2+, Al, and NO on the 30 August 2022 (Table 1). Focusing on major cations at 30 cm depth, the K+, Ca2+, Mg2+ and Al concentrations increased twice successively between 21 June 2022 and 30 August 2022 (Fig. 3). K+, Ca2+, NH also reached outlier concentrations on the 21 June 2022 (Table 1; Fig. S3). The highest concentrations ever registered during 2015–2023 period in 30 cm depth soil solution were reached for the second peak on the 30 August 2022 (Fig. 3, Table 1). K+ concentration exhibited exceptionally high concentrations also at 5 and 10 cm depth.
The 30 cm depth soil solution anomaly during 2022 drought was also characterized by the lowest pH of 4.1 observed on 30 August 2022 over the 2015–2023 period (Fig. 3G). Notably, this low pH anomaly occurred only at 30 cm. However, high pH anomalies were observed at 30 and 60 cm on 21 June 2022 (4.1 and 6.6) and at 10 cm on 30 August 2022 with a pH equal to 5.8. Those values were the maximum pH ever observed at each depth during the period 2015–2023.
For nitrogen-related ions (NO, NH), a first peak in NH concentrations was observed on 21 June 2022 at 30 cm and 60 cm, followed by a second peak of lower concentrations on 30 August 2022 at 30 cm (Fig. 3I). The NO concentration peak of 0.117 mmol L−1 at 30 cm occurred only on 30 August 2022 (Fig. 3J). For superficial soil solutions, at 5 and 10 cm depth, high concentrations were observed on 30 August 2022 for NH (0.10 and 0.16 mmol L−1 respectively) and for NO (0.28 and 0.14 mmol L−1 respectively).
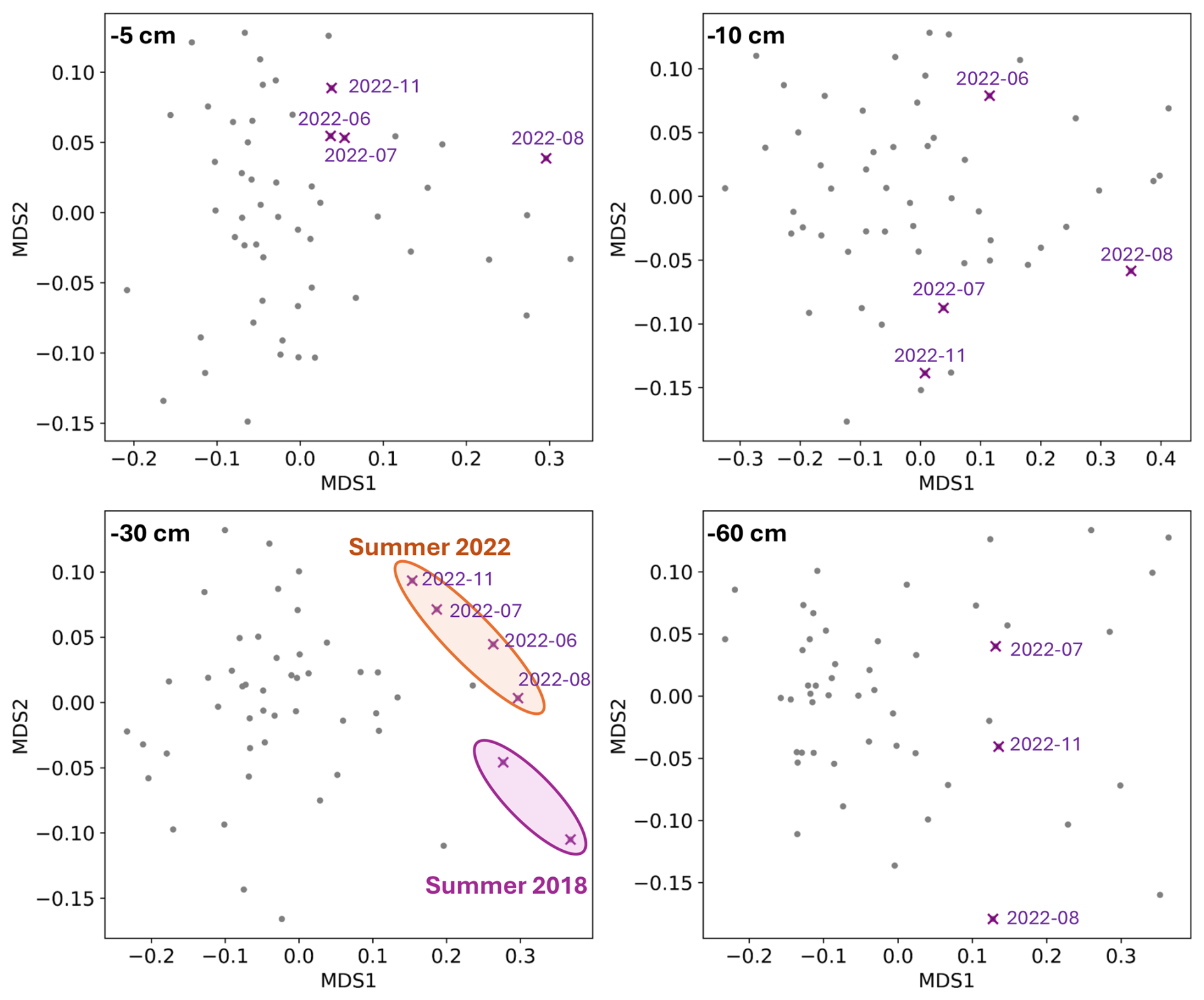
Figure 4Plots of NMDS coordinates of soil solution samples, with separate analysis for each depth. The dates (purple crosses) correspond to soil solution samples during and after the 2022 drought. In the 30 cm figure, two crosses corresponding to the summer of 2018 are positioned on the graph. The number of crosses is lower for samples at 60 cm depths because no water is collected at this depth during certain periods of drought.
Table 2NMDS scores for the various physical and chemical parameters of soil solutions sampled at different depths (5, 10, 30 and 60 cm depths).
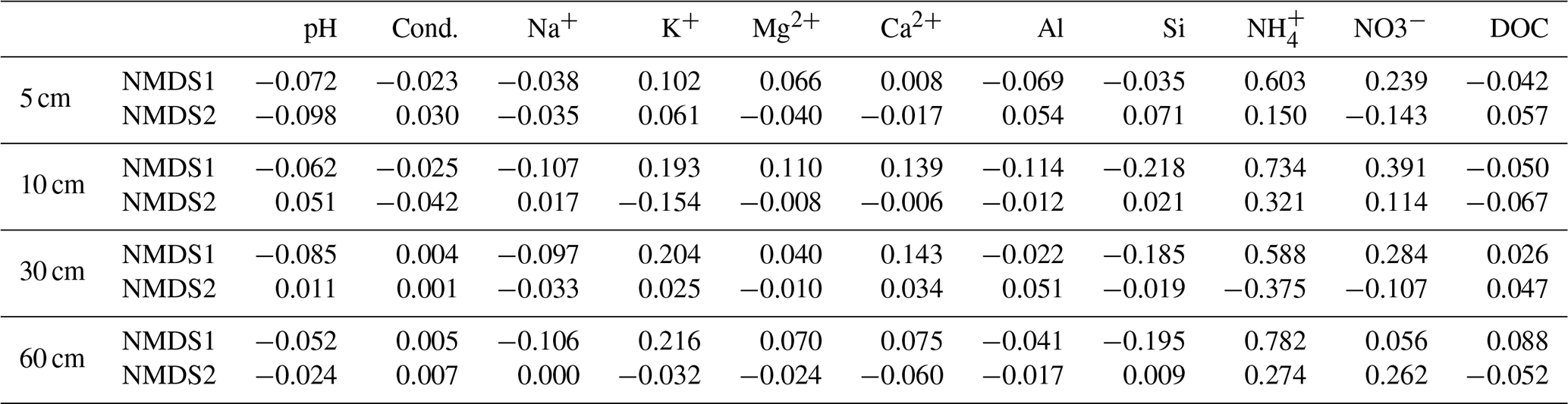
At 30 cm depth, the three points between June and August 2022, which were particularly affected by drought events, were significantly shifted to the right of the plot along the MDS1 axis (Fig. 4). The samples from the summer 2018, which was also a drought episode (cf. 1), also show statistically different coordinates at this depth. Conversely, the values for soil solutions in summer 2022 at the three other depths (5, 10, and 60 cm) did not show any atypical values, except for those in August but only at 5 and 10 cm (Fig. 4). However, these values were less atypical than those observed for soil solutions at a depth of 30 cm (Fig. 4). These observations confirm that the impact of droughts on soil solution chemistry was the most important at 30 cm depth. The NMDS scores for 30 cm soil solutions showed that MDS1 exhibits the highest positive value for NH, followed by NO, meaning a particular anomaly concerning nitrogen species (Table 2). K+ and Ca2+ concentrations also had high scores along the MDS1 axis, indicating that the dynamics of cationic mineral nutrients were also atypical during 2022 drought.
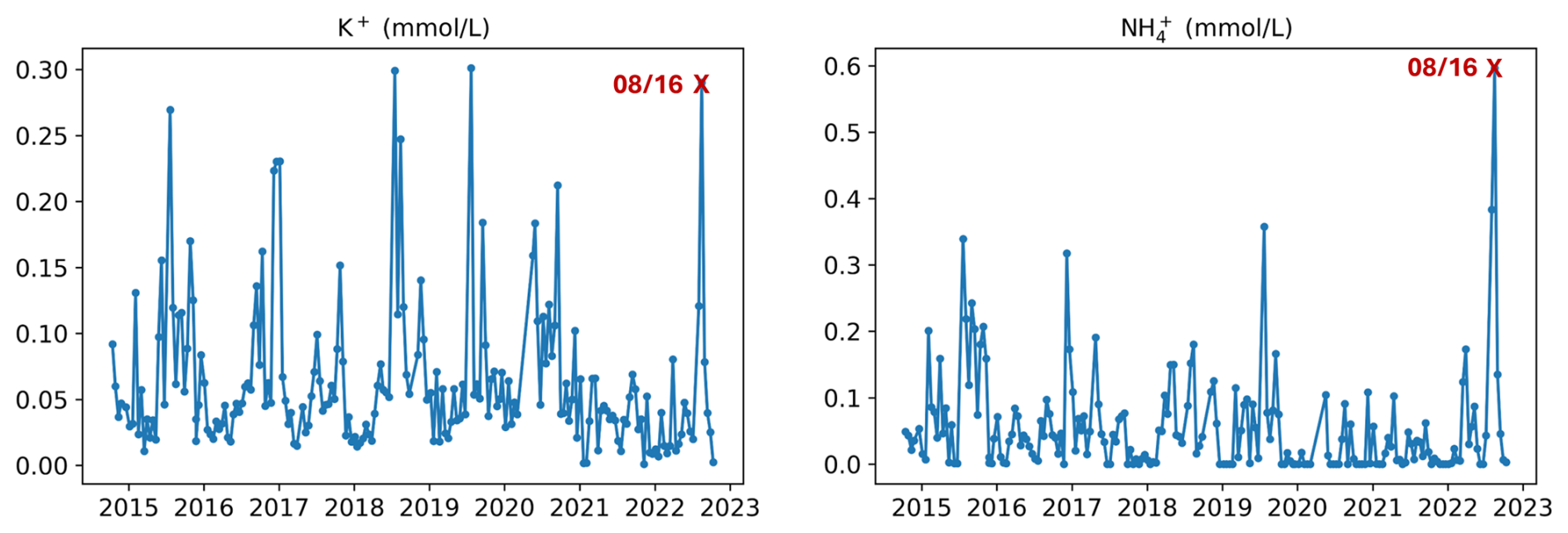
Figure 5K+ and NH concentrations (mmol L−1) in throughfall under spruces in the Strengbach watershed during the period 2015–2023, with the specified point of 16 August 2022 (08/16, red cross) corresponding to the period of rain return after the summer 2022 drought period.
Concentrations in throughfall are highly variable under the studied spruce plot, with numerous concentration peaks, often higher during summer than winter (Fig. 5). Throughfall of 16 August 2022 (red cross in Fig. 5) corresponded to the end of the dry period with the return of rainfall on the 14 August 2022 (Fig. 2A). On this date, throughfall exhibited the highest concentration of NH (0.597 mmol L−1; Fig. 4) and high concentrations of K+ (0.290 mmol L−1).
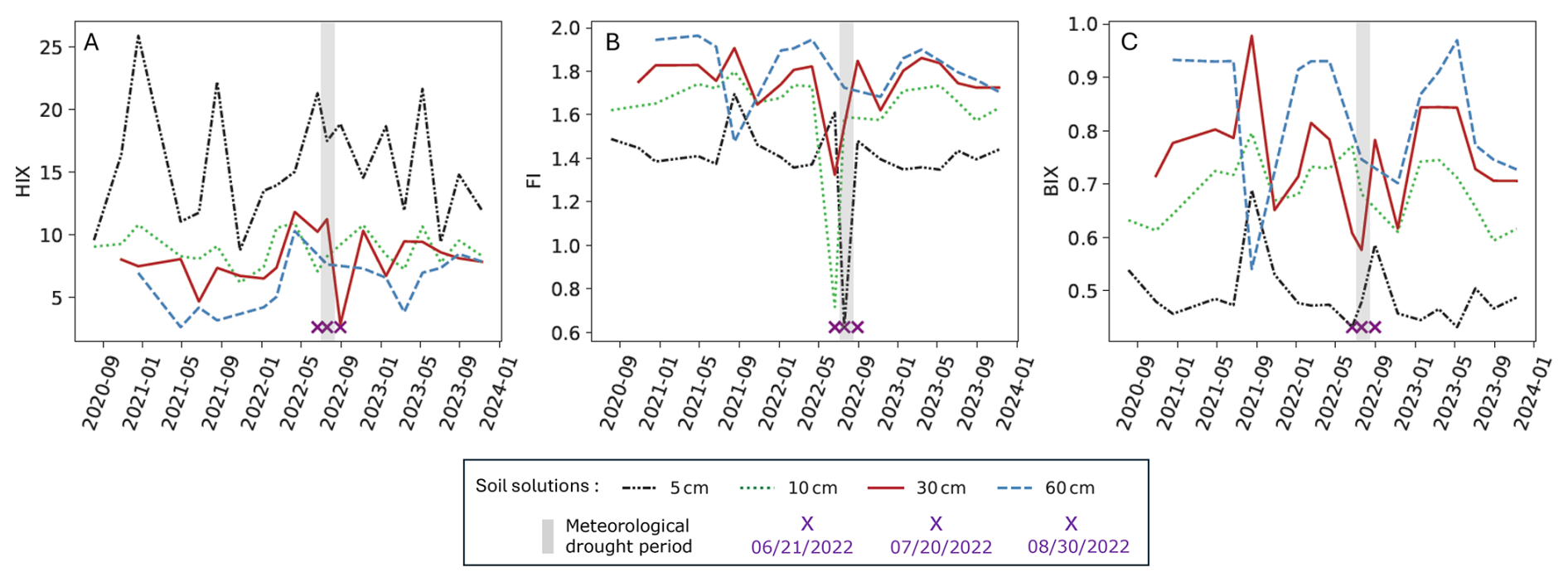
Figure 6Fluorescence index of soil solutions from four different depths (5, 10, 30 and 60 cm) for the 2020–2023 period. HIX (A), FI (B), BIX (C). 2022 Meteorological drought period is represented by a grey band. The purple crosses indicate the dates of three field soil solution sampling corresponding to the drought period (21 June 2022, 20 July 2022 and 30 August 2022).
Table 3Mean, median of HIX, FI and BIX in soil solution at 30 cm depth during the period 2020–2023. Q1 is the 25th percentile, IQR the interquartile range, ( IQR) the lower bound below which points are considered outliers. Values of outliers at 30 cm depth are given in the fourth line with the corresponding date in the fifth line. “X” means that no outlier is found at 30 cm depth during the 2020–2023 period.
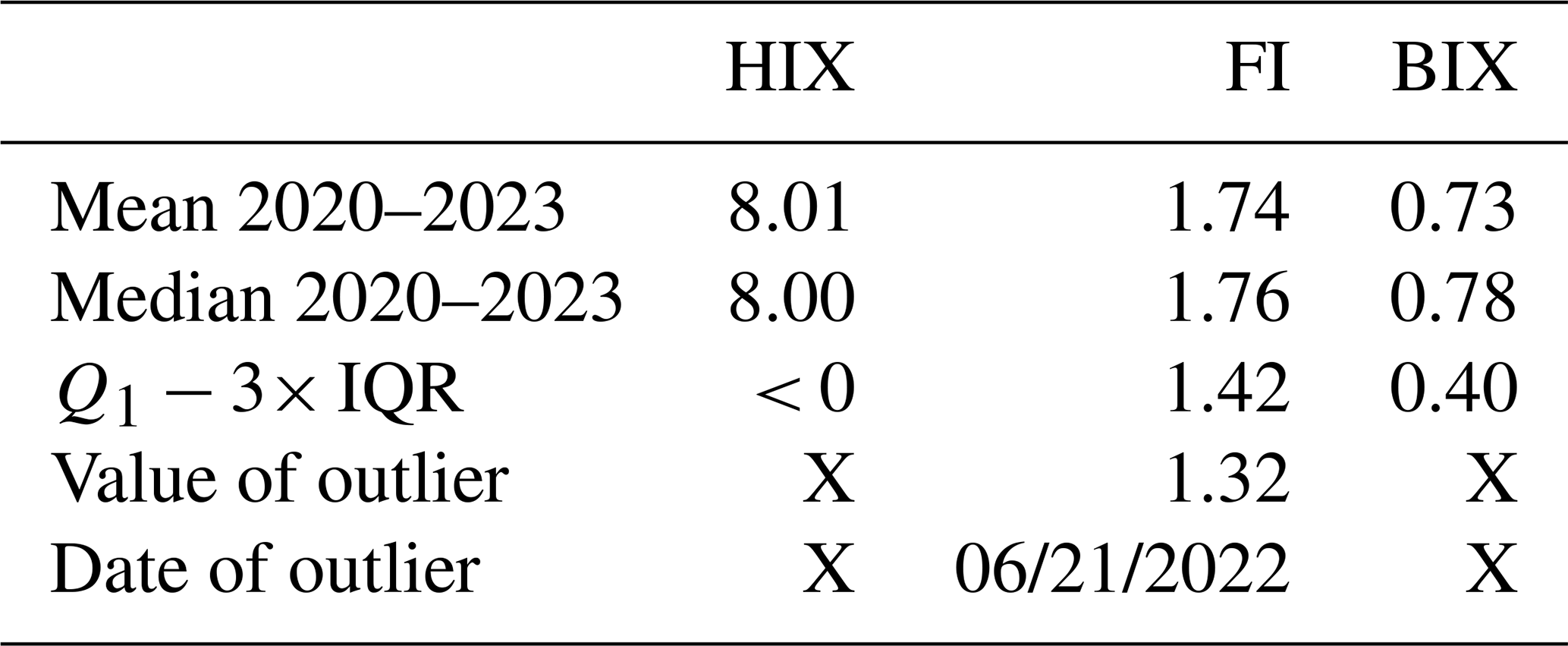
The qualitative indicators of dissolved organic matter presented significant anomalies during the 2022 summer drought. At 30 cm depth, the FI was 1.32 on 21 June 2022, considered as an outlier because the value was below ( IQR = 1.42; Table 3). FI also showed an anomaly at 5 and 10 cm during this drought period with the lowest and atypical values of about 0.62 on 20 July 2022 and 0.72 on 21 June 2022 respectively (Fig. 6). Low BIX values of 0.60 and 0.58 on 21 June 2022 and 20 July 2022 respectively were measured whereas the average value was 0.73 ± 0.11 at 30 cm depth over the 2020–2023 period. HIX reached a minimum value of 2.83 on 30 August 2022, only at 30 cm depth, whereas no significant anomalies were noted at any other depths.
Soil solutions appeared to be most affected by drought at a depth of 30 cm, with significantly different chemical signatures in June, July and August as shown by NMDS analysis (Fig. 4). The highest spruce root density is found in the upper soil, from the base of organic layer to 20 cm depth and sharply decreases with depth (Schmid and Kazda, 2002; Borja et al., 2008). At the Strengbach watershed, the maximum root density ranges from 10 to 30 cm depth (Oursin et al., 2023). Since soil solutions collected at a depth of 30 cm using a zero-tension lysimeter plate incorporate the top 30 centimeters of the profile, this depth is essential for better understanding the processes of tree uptake, and their modifications in response to drought. Moreover, the availability of nutrients for trees is a major concern in soils as poor in alkaline earth elements (Ca and Mg) as those in the Strengbach watershed (Probst et al. 1992; Dambrine et al., 1998; Oursin et al., 2023). We will therefore focus the rest of the discussion on interpreting the data obtained at 30 cm, depth of maximum water/soil/plant interactions, using data from other depths where necessary.
Chemical concentrations in soil solutions are the result of a balance between inputs (throughfall, mineral dissolution, cationic exchange desorption and organic matter degradation) and outputs (plant uptake, mineral precipitation, cationic exchange adsorption and drainage). The following discussion aims to better understand how drought affected the biogeochemical processes responsible for soil solution composition, as weathering, atmospheric inputs, vegetation uptake, organic matter reactivity, N cycle dynamics and acidification.
4.1 Weathering process
While most chemical concentrations showed a peak during the drought of 2022, Na+ and Si exhibited no significant peak (Fig. 3). Their average concentrations during the period between 21 June and 30 August 2022 were 0.074 and 0.038 mmol L−1 respectively, which were within the range of inter-annual variability with a mean concentration during 2015–2023 period of 0.066 ± 0.025 mmol L−1 (median ± IQR) and 0.061 ± 0.046 mmol L−1 for Na+ and Si respectively (Table 1).
In the soils from the Strengbach catchment, sodium is mainly included in albite, a primary mineral from the granite (Fichter et al., 1997). Na is not considered to precipitate or integrate in secondary phases (Chou and Wollast, 1985). It represents only 0.6 % of the cationic exchange capacity and its concentration is negligible in the litter compared to other elements (Oursin et al., 2023). Thus, the main sources of sodium in soil solution are the throughfall and the dissolution of albite. The concentrations of Na+ in throughfall did not seem to be affected by drought, the concentration on the 16 August 2022, date of rain return after drought, was 0.033 mmol L−1, which was comparable with the mean concentration of 0.042 ± 0.021 (Supplement Table S2). As no significant consequences of drought on Na+ concentration in soil solution were observed, we can propose that drought does not affect albite dissolution in soil. Moreover, the study of Oursin et al. (2023) showed that the contribution of primary mineral weathering in the chemical signature (Ca, Mg and K concentrations) of soil solution from the same site was low and even negligible when compared with other fluxes (alteration of secondary minerals, cation exchange, litter degradation), except for Na. This lack of impact on albite dissolution may be extended to other primary minerals composing granite. Consequently, it can be proposed that the impact of drought on the weathering of all primary minerals is negligible.
In theory, thermodynamic and kinetic of mineral dissolution depend highly on soil temperature and water content (Lasaga et al., 1994; Kump et al., 2000). Studies modelling the effect of climate change on future forest soil weathering rate proposed that a global increase in temperature can increase the chemical weathering. However, because of soil moisture limitation, summer droughts limit the expected gain in weathering associated with higher temperatures (Belyazid et al., 2022; Kronnäs et al., 2023). Thus, severe water deficits in soil tend to reduce weathering rates.
The concentrations of Si in rain and throughfall are below the detection limit (Pierret et al., 2019) and the concentration of Si in litter is negligible compared with other elements (Oursin et al., 2023). Therefore, Si mainly originates from mineral weathering. Hydrological modelling of soil water content (Fig. 2B) exhibited an important diminution during June 2022 in the third layer (between 10 and 30 cm) and during August in the fourth layer (30 to 60 cm), consistent with the Si concentrations with significant reduction on 21 June 2022 at 30 cm and on 30 August 2022 at 60 cm (Fig. 3F). In addition, two minimal Si concentrations, at 30 and 60 cm depth, were observed in the summers 2018 and 2022, the two main drought events of the period 2015–2023, during which impacts on forests were observed (Gharun et al., 2024; Knutzen et al., 2025). So that we can deduce a reduction in silicate mineral dissolution due to low water content during drought. Silica is mainly controlled by clay weathering in the soil of the Strengbach catchment, as shown by geochemical modelling (Goddéris et al., 2006; Beaulieu et al., 2020). As the reduction was visible for Si and not for Na, it is suggested that clay mineral dissolution is reduced during drought events. As the dissolution kinetic is higher for secondary minerals compared to primary, a reduction of secondary minerals dissolution should have more pronounced consequences on the chemical composition of the solution (Drever and Clow, 1995). Thus, water deficit in soils leads to a decrease in weathering rate, as proposed by Belyazid et al. (2022) and Kronnäs et al. (2023), but only visible on secondary minerals such as clays. Silica was the only element showing a reduction in concentration during drought. However, chemical soil solutions signals exhibited positive peaks in many elements such as K+, Ca2+, Mg2+ and Al (Fig. 3). These peaks cannot be explained by an expected reduction in weathering due to water deficit, which would produce the opposite effect. Therefore, other processes will be discussed in the following sections.
4.2 The role of throughfall
Rainfall passing through the canopy, known as throughfall, differs from open field precipitation because of its interactions with leaf surfaces. Throughfall integrates wet and dry atmospheric deposits as well as biological interactions with the canopy during leaf or needle leaching (Ulrich, 1983; Berger et al., 2008; Eisalou et al., 2013; Pierret et al., 2019; Ponette-González et al., 2020). Compared to rain, throughfalls are enriched in each chemical element. On the basis of a monitoring over the 1986–2012 period, Pierret et al. (2019) showed that throughfalls under spruces are enriched by a factor of 15 for K+, 5 for Ca2+ and Mg2+ and 2 for NH compared to open field precipitations from the same studied site, the Strengbach catchment. K+ in throughfalls comes mainly from biological excretion by leaves, whereas Ca2+ and Mg2+ are a mixture between wet and dry atmospheric deposits with a weak influence of biological exchanges (Lovett and Lindberg, 1984; Kopáček et al., 2009; Adriaenssens et al., 2012; Pierret et al., 2019). NH mainly comes from atmospheric deposition, which might be from NH3 atmospheric emission due to agricultural activities (Avila et al., 2017; Pierret et al., 2019). But NH is involved in numerous biological processes, can also be excreted, absorbed by leaves or organisms living on the leaves (Woods et al., 2012; Ponette-González et al., 2020).
K+ dynamics are highly affected by the biological functioning of trees. Reduction of rainfall or drought might reduce the mobility of K+ within the canopy by decreasing foliar leaching (Sardans and Peñuelas, 2015; Schlesinger et al., 2016; Touche et al., 2024). Consequently, biologically excreted potassium accumulated on spruce needles during the 2022 dry period. Upon the return of rain, these accumulated elements were leached, leading to high concentrations in the throughfall (Fig. 5). Thus, the exceptionally high peak in K+ observed in soil solutions at 5, 10, and 30 cm on 30 August 2022 can be linked to the high K+ content in throughfall on 16 August 2022. Indeed, K+ concentration on the 30 August 2022 soil solution at 5 cm depth (integrating 20 July to 30 August period and including the rain episode of 16 August; Fig. 2) was 0.331 mmol L−1, similar to the K+ concentration in throughfall on the 16 August 2022 (Fig. 3B; 0.290 mmol L−1), thus supporting the important contribution of throughfall in superficial soil solution K+ concentration. Similar observations of K+ concentrations peak in soil solutions at different depths have been reported on drought experiments on European spruce plots (Lamersdorf et al., 1998) but also in Canadian boreal forest with very high K+ concentrations in the soil solution at 30 cm and in throughfall, as observed by Houle et al. (2016). This vertical propagation of K+ in the soil profile was likely driven by the increased concentration observed in the throughfall. This spread of K+ diminished between 30 and 60 cm, probably because this cation was sorbed, exchanged, precipitated or uptaken by trees. K+ was the only cation that exhibited a maximum peak in both throughfall and surface soil solutions (5 and 10 cm deep). This specificity of K+ is linked to the important contribution of throughfall in K+ soil solution concentration compared to other elements.
NH showed a strong positive anomaly in the throughfall for 16 August 2022, with concentration of 0.597 mmol L−1, more than 10 times the average for the period 2015–2022 of 0.055 mmol L−1 (Fig. 5). This ammonium peak was also found in soil solutions at depths of 5 and 10 cm on 30 August 2022 (Fig. 3). Similarly to K+, the ammonium flux in throughfalls resulted from the leaching of spruce needles during the first rain after drought. However, due to the high reactivity of ammonium and nitrogen species in soils (as nitrification converts ammonium into nitrite and/or nitrate), the NH concentrations of soil solutions at 5 and 10 cm depth were significantly lower (0.075 and 0.150 mmol L−1) than in throughfall (0.597 mmol L−1). More frequent sampling of soil solutions would have provided a better match between the chemical composition of throughfall (sampled every two weeks) and that of soil solutions (sampled every six weeks), particularly for better observation of the behaviour of highly reactive species such as nitrates and ammonium. It can still be argued that, after a drought period, throughfall represents an important source of ammonium for soil solutions.
Since 2015, other peaks of high concentrations in the throughfall series have been observed, but not in the soil solutions (Figs. 3, 5). This highlights that additional processes must be involved to explain why, in 2022, K, Ca or Mg accumulated in soil solutions, whereas in other periods, part of the throughfall appeared to be mobilized in the soil.
4.3 Vegetation uptake
Soil solutions contain elements that are essential plant nutrients such as K+, Ca2+, Mg2+, NO, NH (Ingestad, 1979). K is the most abundant cation in plant cells and a limiting element for plant growth (Leigh and Jones, 1984; Sardans and Peñuelas, 2015). Ca and Mg are also two essential nutrients for trees, found in very small quantities in Strengbach soils and maintained in sufficient concentration in the soil solution through recycling by vegetation (Ovington, 1959; Fichter et al., 1997; Beaulieu et al., 2020; Oursin et al., 2023). Beaulieu et al. (2020) simulated biogeochemical processes under spruce plot in the Strengbach catchment, and they estimated that tree uptake is the most important flux of Ca and Mg in the system, two times higher than the flux coming from the throughfall. In contrast to K, the uptake flux is two times lower than the flux coming from throughfall. So, trees uptake large quantities of Ca2+, Mg2+, K+, NH, NO, and influence nutrient concentrations in solution. The field data showed an increase of Ca2+, Mg2+, K+, NH and NO concentrations in the soil solutions, especially at 30 cm depth, during or after the drought (Fig. 3). We hypothesize that these increases in concentrations were linked to a decrease in biological uptake by vegetation.
The reactions of trees to drought, such as stomate closure or reducing leaf area (needle loss), for instance, decrease water loss through transpiration and so decrease water uptake by roots (Bréda et al., 2006; Ditmarova et al., 2010; Hesse et al., 2024). The decrease in tree transpiration during the drought period, simulated by the model BILHYDAY used in this study (Fig. 2), implies a reduction in water uptake by tree roots. This was also observed in the Strengbach watershed, with measurements of reduced sap flow during an experimental water deficit on spruce trees (Lu et al., 1995). In addition, an experimental water deficit study carried out on a spruce stand at the same site showed that the composition of the xylem sap was lower in nutrients content (Ca, Mg and K) during the water deficit period (Dambrine et al., 1993). Thus, trees take up less Ca, Mg and K during drought. Reduced nutrient uptake can explain the lower biomass concentrations found in numerous experimental water deficit studies on forest plots. For example, lower concentrations of K, B and Mg were observed after droughts in spruce needles (Nilsen, 1995), but also in the biomass of other tree species (Sardans et al., 2008; Touche et al., 2022).
However, soil solution collected by zero-tension lysimeters is the fraction of soil water participating in gravitational flow, circulating mainly in the macropores (Singh et al., 2018). It cannot be considered similar to water collected by vegetation, since roots also collect water from microporosity with a certain suction pressure (Rüdinger et al., 1994). Furthermore, gravitational solutions have a very different chemical composition from other fractions of soil water collected under a certain pressure (Haines et al., 1982; Watmough et al., 2013), also under the spruce forests of the Strengbach watershed (Ranger et al., 1993). But the soil on this plot is very sandy, with a sand content ranging from 50 % on the surface to 70 % at depth, and are relatively homogeneous (Oursin et al., 2023). Thus, the studied soil is very porous (more macro- than micro-porosity) and well-drained (more gravity flow than capillarity), therefore we assume that gravitational soil solution is representative of the whole soil water. In addition, gravity soil solutions enable input-output balances to be calculated, given that the soil solution at a depth of 30 cm can be considered as water leaving the root zone of spruce trees (Ranger et al., 1993; Marques et al., 1996). The chemical composition of the solution thus collected depends on all the biogeochemical processes taking place in the soil and can indirectly indicate a change in root uptake.
Thus, the decrease in biological uptake by trees during drought may explain the maximum concentrations observed for Ca, Mg, K, nitrate and ammonium in soil solutions, particularly at a depth of 30 cm, which corresponds to the depth after the maximal root density zone, in the first 20 cm of the mineral soil for spruces (Schmid and Kazda, 2002; Borja et al., 2008). It explains why high concentrations were found at 30 cm depth particularly for Ca and Mg, the two nutrients most impacted by trees uptake.
4.4 Organic carbon reactivity
A DOC increase was observed in soil solution at 30 cm depth during and after drought period, with a maximum concentration of 66.81 ppm-C after the rain return (Table 1; Fig. 3). On other sites, similar DOC peaks have been reported under spruce plots, after a water deficit experiment at 20 cm depth (Lamersdorf et al., 1998) or after 2006 drought at 90 cm depth (Schulze et al., 2011). The DOC increase during the drought period could have been induced by different organic inputs. Indeed, Gaul et al. (2008) reported that water deficit can lead to an increase in fine root mortality up to 61 %, not compensated by an increase in fine root production on a Norway spruce forest in Germany. This root mortality, also observed in Norway spruce located in Sweden subjected to drought (Persson et al., 1995), can represent a carbon source to soil organic matter pool in a temperate ecosystem.
Moreover, another source of carbon in upper soil horizons in response to drought could stem from stress and mortality in bacteria and fungi. As reported in many studies, water deficit leads to a decrease in organic matter degradation (Schimel et al., 2007; Boczón et al., 2021; Gehring et al., 2017; Bogati and Walczak, 2022). Indeed, even if heterotrophic respiration is controlled by temperature in wet soils, it is limited by water deficit during dry period (Curiel Yuste et al., 2007). The degradation decrease could lead to accumulation of organic carbon in superficial soil layer. On the other hand, the high concentration of DOC observed after the rain return in soil solutions could be the result of fast-moving water and transport of DOC accumulated in superficial soil layer during drought period through soil profile. A high dissolution of organic matter would also lead to an input of other elements contained in this organic matter to the soil solution. Therefore, this mechanism likely contributed to the observed increase in Ca, Mg or K concentrations in soil solutions at 30 cm after rewetting, in addition to other mechanisms disturbed during drought.
To better understand this modification of DOC dynamics, optical characterization by fluorescence is employed. Three fluorescence indices are frequently used to characterize natural organic matter in water: the fluorescence index (FI, McKnight et al., 2001), the biological index (BIX, Huguet et al., 2009) and the humification index (HIX, Zsolnay et al., 1999). HIX was defined by Zsolnay et al. (1999) and is an indicator sensitive to the quantity of aromatic molecules in solution, but also to the type of molecules, i.e., more or less aromatic, heavy, complex (Serène et al., 2025). HIX was relatively high before and during the drought in 30 cm depth soil solution (median of 8.00; Table 3). It then fell sharply when the rain returned after the long dry period (HIX = 2.83 on 30 August 2022; Fig. 6) during the DOC peak. A similar dynamic between DOC and aromaticity during and after drought has been observed in several streams in boreal ecosystems (Tiwari et al., 2022). As these large aromatic molecules precipitate at acid pH (Swift, 1996), the decrease in pH may explain the decrease in HIX on 30 August 2022. HIX is often correlated with DOC (Pearson coefficient = 0.54 on all four depths of the plot), but the solution showing the DOC peak after the drought was relatively low in aromatic molecules (Figs. 3, 6), reflecting the atypical composition of this solution, reinforcing the hypothesis of a new source of organic matter originating from biological stress on soil organisms, rather than usual litter decomposition.
4.5 Biological stress monitored with UV-fluorescence spectroscopy
The FI in superficial waters and groundwaters is used to distinguish sources of terrestrially or microbially derived fulvic acids. McKnight et al. (2001) propose index values of 1.4 for terrestrial origin and of 1.9 for microbial origin. In our study, FI averaged 1.74 in soil solution at 30 cm depth over the period 2020–2023 (Table 3), which could highlight predominantly microbial sources of organic material. However, a decrease of FI to 1.3 was observed in June 2022 at 30 cm depth (Fig. 6), an index value consistent with predominantly terrestrial sources. The low contribution of microbial source to organic material could be explained by a reduction of fungal and bacterial activity due to low water content during the May–June period (Fig. 2B), as mentioned in Sect. 4.5. The index value increased to 1.8 in soil solution with the rain return, indicating a recovery of regular microbial activity (Fig. 6).
The BIX is commonly used as a proxy of biological activity in aquatic environments (Parlanti et al., 2000; Huguet et al., 2009). High values of BIX suggest a high biological activity and so an organic matter recently produced by microbial activity, while the low values are supposed to stem from relatively low biological activity. The BIX values decreased in soil solution at 30 cm depth during the May–June period, then increased upon rewetting (Fig. 6), consistent with the FI value evolution.
The drought characterized by a decrease in soil water content from May to July (Fig. 2), led to a DOM with a lower microbial signature. This supports the hypothesis of a decrease in soil microbial and fungal activity during drought. On 30 August 2022, after the soil rewetting, the DOM regained its usual microbial source signature, reflecting a resumption of biological activity once favorable water conditions were restored.
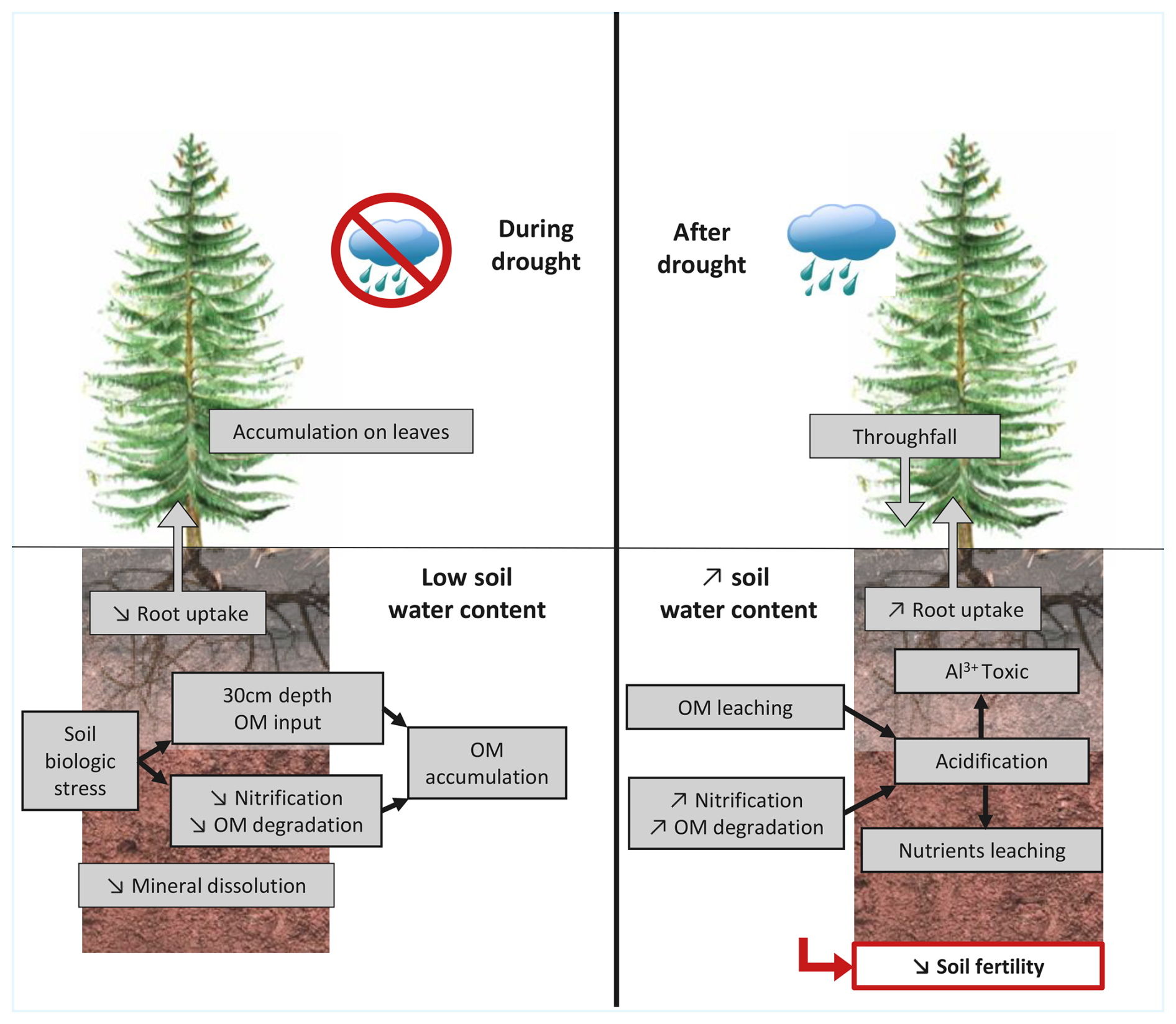
Figure 7Graphical summary of drought perturbation on soil solution chemistry and soil reactivity with grey boxes representing biogeochemical processes and a red box representing the consequences of drought on soil fertility.
4.6 Nitrogen cycle disturbance during drought
While a NO peak was only observed after the return of rainfall, NH exhibited two distinct dynamics: high concentrations at 30 and 60 cm in June and high concentrations at 5 and 10 cm after rewetting in August. These temporal dynamics are explained below in chronological order.
During the water deficit period in soil (Fig. 2), a peak of NH concentration on 21 June 2022 (Fig. 3) occurred only at 30 and 60 cm depth. The disturbance of the nitrogen cycle during drought has been the subject of numerous historical studies, notably with experimental droughts on various spruce plots on acid soils in Europe, where canopy roofs were installed (Lamersdorf et al., 1998). They all observed high concentrations of ammonium and dissolved organic carbon in soil solution. Gangloff et al. (2014) observed that ammonium peaks in soil solutions are also linked to specific periods of bacterial activity and to hydrological conditions, in a spruce plot from the Strengbach catchment. This supports the idea of an accumulation of organic matter during water deficit due to stress on the soil biosphere (cf. 4.4). The mortality of bacteria and fungi may release cell content that is rich in nitrogen-containing organic matter. This organic nitrogen is degraded to ammoniacal nitrogen by ammonification (Kuypers et al., 2018). The accumulation of NH is attributed to the interruption of nitrification during water deficits, whereas ammonification is less sensitive, as demonstrated by experiments on spruce soils carried out by Hentschel et al. (2007) or Chen et al. (2011). Thus, the high concentrations of NH at deeper horizons, i.e. 30 and 60 cm depth, can be related to the punctual stress of the soil microfauna. This is consistent with the FI and BIX index, which showed a less microbially derived signature at the corresponding period (cf. 4.5).
After the return of rainfall (30 August 2022), the NH concentrations peaked in soil solutions at 5 and 10 cm depths. This can be attributed to the throughfall inputs, as previously discussed (cf. 4.2). At the same time, high nitrate concentrations were observed in soil solutions at all depths (Fig. 3). After soil rewetting, conditions became favorable for nitrifying bacteria: high ammonium concentration, high pH, high temperature and sufficient water content. This led to significant development of nitrifying populations at 30 cm depth, as observed by Krüger et al. (2021) after the 2018 drought under a beech plot. This resulted in high nitrification after rain returned, leading to the observed high NO concentrations in soil solutions. In addition, nitrate accumulation in soil solutions was also exacerbated by the lower tree uptake (cf. 4.3).
Thus, periods of water deficit and the first episodes of rewetting, by modifying bacterial activity, severely disrupt the cycle of nitrogen and nitrogen nutrients (NH and NO) concentrations in soil solutions.
4.7 Acidification
The most acidic pH (4.1) recorded in soil solution at 30 cm depth during the 2015–2023 monitoring period was observed on 30 August 2022. In contrast, a significantly higher pH (5.2) was measured on June 21, coinciding with low soil moisture content (Fig. 2). The ammonification consumes protons and consequently increases pH (Kuypers et al., 2018). This process likely explains the highest pH of the 2015–2023 period at 30 cm with a value of 5.21 observed on 21 June 2022 (Fig. 3). Conversely, the low pH coincided with exceptionally elevated nitrate concentrations observed in August (Fig. 3) because nitrification is acidifying (Kuypers et al., 2018). The sharp pH variation illustrates the strong influence of the nitrogen cycle on the pH of those soil solutions. In addition, this low value of pH can also be related to the high DOC concentration, which was the highest ever measured at 30 cm depth (Fig. 3). Indeed, DOC is composed mainly of organic acids, contributing to acidification (Strobel et al., 2001).
Furthermore, this acidification has consequences for the dynamics of some elements in the soil, as protons can desorb cations from soil exchangeable surface, in particular Al3+, Ca2+, Mg2+ and K+ (Ulrich et al., 1980; Fest et al., 2005; Meng et al., 2019). This process resulted in the release of those cations into soil solution and likely contributed to the elevated concentrations in solution at 30 cm depth (Fig. 3). This is especially noticeable with Al concentrations, with the highest concentration ever observed during the 2015–2023 period at 30 cm depths on 30 August 2022 (Fig. 3). It raises issues for tree health, as Al3+ is toxic in high concentrations (Ulrich et al., 1980; De Wit et al., 2001). The desorption of cationic nutrients such as Ca2+, Mg2+ and K+ is of particular concern in acid soils that are already low in cationic nutrients, such as those in the Strengbach catchment (Probst et al., 1992; Dambrine et al., 1998; Oursin et al., 2023).
The summer drought of 2022 was the most severe ever recorded at the observatory of the Strengbach catchment since 1986. Ten years of monitoring at the spruce plot have enabled the identification and characterisation of the main geochemical disturbances in soil solutions (in terms of major elements, dissolved organic matter and its indicators) linked to both drought and the effects of the first rains, as well as the consequences for nutrient biogeochemical cycles and for the whole functioning of the forest ecosystem. We focused our discussions on soil solutions at 30 cm depth because they exhibited the highest sensitivity to drought and represent the preferential depth for root uptake. The 2022 drought caused a significant increase in DOC and ion concentrations in soil solutions (K+, Ca2+, Mg2+, Al3+, NH, NO) and atypical values of fluorescence indices HIX, FI and BIX. The consequences of drought on biogeochemical processes can be separated into two time periods: during drought and after drought (summarized in Fig. 7).
During drought, the impact on primary mineral dissolution remained negligible, whereas the decrease in water content in soil induced a decrease in secondary mineral weathering such as clays. Concurrently, the water deficit reduced the tree uptake of water and nutrients. A modification of the DOM quality in soil solution was also observed suggesting a biological stress on soil biosphere due to the exceptionally dry belowground conditions. This resulted in the accumulation of organic matter within the soil. Nitrification was also inhibited, resulting in an accumulation of NH and an increase in pH in soil solutions.
After drought, the return of rainfall leached the accumulated atmospheric depositions and biological recretion on spruce needles. It resulted in high concentrations in throughfall, particularly for K+ and NH. The increase in soil water content mobilised the organic matter accumulated during the dry period and induced the highest DOC (66 ppm-C) ever registered at 30 cm deep since 2015. In addition, the degradation of organic matter also generated an important flux of Ca, Mg and K into soil solutions. Sufficient water content, high NH and high temperature triggered nitrification to reactivate and produce high NO concentration in solution. As organic matter degradation and nitrification are acidifying processes, it resulted in the lowest pH (4.1) ever registered at 30 cm during the 2015–2023 period. This acidification led to a desorption of soil cationic nutrients from the clay-humic complex. Thus, immediately after the return of the rainfall, the soil solutions at 30 cm depth had the highest concentrations of nutrients (K+, Ca2+, Mg2+, NH, NO) for the 2015–2023 period. It also generated a very high concentration of desorbed Al3+, which is toxic for trees. This acidification promotes nutrient leaching and thus leads to a decline in the fertility of forest soils, which is particularly worrying in already nutrient-poor acidic soils such as those in the Strengbach watershed.
In addition to the water deficit that threatens spruces with severe physiological consequences, drought also induces nutrient loss and so accelerates forest decline and fragility. Climate forecasts predict an increase in the frequency and intensity of droughts in the Vosges Mountains, which will likely exacerbate mortality in spruce forests that are particularly non-resilient to drought. This aggravation of forest decline by drought suggests a negative feedback loop that could exacerbate climate change by reducing the carbon storage capacity of forests and by releasing carbon due to tree mortality. Spruce trees, already weakened by nutrient-poor soils and bark beetle attacks, appear to be poorly adapted in temperate regions to drought conditions, which are likely to recur. Forest management must take this into account to achieve more sustainable forests and preserving the ecosystem services provided by forests, such as carbon sinks, biodiversity preservation, cooling effects, and protection of water quality in mountain areas.
Weather data and data on the chemistry of soil solution and throughfall are available on the BDOH platform: https://bd-ohge.unistra.fr/OHGE/, last access: 18 September 2025. Data concerning soil solution and throughfall are also provided in the Supplement (Tables S1, S2).
The supplement related to this article is available online at https://doi.org/10.5194/bg-23-1569-2026-supplement.
AS, MCP and EB designed the study. MNP produces soil solution fluorescence measurements. BB processed meteorological data and produced hydrological simulations. AS ran statistical analysis. AS interpreted data and wrote the manuscript. MCP, EB, BB and MNP commented the manuscript.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We acknowledge all the team of the Observatoire hydrogéochimique de l'environnement: Solenn Cotel, Colin Fourtet, Anthony Julien and Alain Hernandez for field sampling and laboratory analysis.
This research has been supported by the Conseil régional du Grand Est, the École Nationale du Génie de l'Eau et de l'Environnement de Strasbourg, and the Centre National de la Recherche Scientifique, Institut national des sciences de l'Univers (REFORESTATION research project).
This paper was edited by Cindy De Jonge and reviewed by two anonymous referees.
Adriaenssens, S., Hansen, K., Staelens, J., Wuyts, K., De Schrijver, A., Baeten, L., Boeckx, P., Samson, R., and Verheyen, K.: Throughfall deposition and canopy exchange processes along a vertical gradient within the canopy of beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst), Sci. Total Environ., 420, 168–182, https://doi.org/10.1016/j.scitotenv.2011.12.029, 2012.
Anderegg, W. R. L., Kane, J. M., and Anderegg, L. D. L.: Consequences of widespread tree mortality triggered by drought and temperature stress, Nat. Clim. Change, 3, 30–36, https://doi.org/10.1038/nclimate1635, 2013.
Anderegg, W. R. L., Trugman, A. T., Badgley, G., Konings, A. G., and Shaw, J.: Divergent forest sensitivity to repeated extreme droughts, Nat. Clim. Chang., 10, 1091–1095, https://doi.org/10.1038/s41558-020-00919-1, 2020.
Avila, A., Aguillaume, L., Izquieta-Rojano, S., García-Gómez, H., Elustondo, D., Santamaría, J. M., and Alonso, R.: Quantitative study on nitrogen deposition and canopy retention in Mediterranean evergreen forests, Environ. Sci. Pollut. Res., 24, 26213–26226, https://doi.org/10.1007/s11356-017-8861-4, 2017.
Beaulieu, E., Pierret, M.-C., Legout, A., Chabaux, F., Goddéris, Y., Viville, D., and Herrmann, A.: Response of a forested catchment over the last 25 years to past acid deposition assessed by biogeochemical cycle modeling (Strengbach, France), Ecol. Modell., 430, 109124, https://doi.org/10.1016/j.ecolmodel.2020.109124, 2020.
Belfort, B., Alzein, A., Cotel, S., Julien, A., and Weill, S.: Hydrodynamic Parameter Estimation for Simulating Soil-Vegetation-Atmosphere Hydrology Across Forest Stands in the Strengbach Catchment, Hydrology, 13, 11, https://doi.org/10.3390/hydrology13010011, 2025.
Belyazid, S., Akselsson, C., and Zanchi, G.: Water limitation in forest soils regulates the increase in weathering rates under climate change, Forests, 13, 310, https://doi.org/10.3390/f13020310, 2022.
Berger, T. W., Untersteiner, H., Schume, H., and Jost, G.: Throughfall fluxes in a secondary spruce (Picea abies), a beech (Fagus sylvatica) and a mixed spruce–beech stand, For. Ecol. Manage, 255, 605–618, https://doi.org/10.1016/j.foreco.2007.09.030, 2008.
Biron, P.: Le Cycle de l'eau En Forêt de Moyenne Montagne: Flux de Sêve et Bilans Hydriques Stationnels: Bassin Versant Du Strengbach à Aubure, Hautes Vosges, Ph.D. Thesis, Université Louis Pasteur (1971–2008), Strasbourg, France, OCLC Number: 496099637, 1994.
Boczoń, A., Hilszczańska, D., Wrzosek, M., Szczepkowski, A., and Sierota, Z.: Drought in the forest breaks plant–fungi interactions, Eur. J. For. Res., 140, 1301–1321, https://doi.org/10.1007/s10342-021-01409-5, 2021.
Bogati, K. and Walczak, M.: The Impact of Drought Stress on Soil Microbial Community, Enzyme Activities and Plants, Agronomy, 12, 189, https://doi.org/10.3390/agronomy12010189, 2022.
Borja, I., De Wit, H. A., Steffenrem, A., and Majdi, H.: Stand age and fine root biomass, distribution and morphology in a Norway spruce chronosequence in southeast Norway, Tree Physiol., 28, 773–784, https://doi.org/10.1093/treephys/28.5.773, 2008.
Bréda, N., Huc, R., Granier, A., and Dreyer, E.: Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences, Ann. For. Sci., 63, 625–644, https://doi.org/10.1051/forest:2006042, 2006.
Brioschi, L., Steinmann, M., Lucot, E., Pierret, M.C., Stille, P., Prunier, J., and Badot, P. M.: Transfer of rare earth elements (REE) from natural soil to plant systems: implications for the environmental availability of anthropogenic REE, Plant Soil, 366, 143–163, https://doi.org/10.1007/s11104-012-1407-0, 2013.
Cenki-Tok, B., Chabaux, F., Lemarchand, D., Schmitt, A.-D., Pierret, M.-C., Viville, D., Bagard, M.-L., and Stille, P.: The impact of water–rock interaction and vegetation on calcium isotope fractionation in soil- and stream waters of a small, forested catchment (the Strengbach case), Geochimica et Cosmochimica Acta, 73, 2215–2228, https://doi.org/10.1016/j.gca.2009.01.023, 2009.
Chen, Y.-T., Borken, W., Stange, C. F., and Matzner, E.: Effects of decreasing water potential on gross ammonification and nitrification in an acid coniferous forest soil, Soil Biol. Biochem., 43, 333–338, https://doi.org/10.1016/j.soilbio.2010.10.020, 2011.
Chou, L. and Wollast, R.: Steady-state kinetics and dissolution mechanisms of albite, Am. J. Sci., 285, 963–993, https://doi.org/10.2475/ajs.285.10.963, 1985.
Ciais, Ph., Reichstein, M., Viovy, N., Granier, A., Ogée, J., Allard, V., Aubinet, M., Buchmann, N., Bernhofer, Chr., Carrara, A., Chevallier, F., De Noblet, N., Friend, A. D., Friedlingstein, P., Grünwald, T., Heinesch, B., Keronen, P., Knohl, A., Krinner, G., Loustau, D., Manca, G., Matteucci, G., Miglietta, F., Ourcival, J. M., Papale, D., Pilegaard, K., Rambal, S., Seufert, G., Soussana, J.F., Sanz, M. J., Schulze, E. D., Vesala, T., and Valentini, R.: Europe-wide reduction in primary productivity caused by the heat and drought in 2003, Nature, 437, 529–533, https://doi.org/10.1038/nature03972, 2005.
Copernicus Climate Change Service (C3S): European State of the Climate 2022, ERA5, E-OBS [data set], https://doi.org/10.24381/gvaf-h066, 2023.
Curiel Yuste, J., Baldocchi, D.D., Gershenson, A., Goldstein, A., Misson, L., and Wong, S.: Microbial soil respiration and its dependency on carbon inputs, soil temperature and moisture, Global Change Biol., 13, 2018–2035, https://doi.org/10.1111/j.1365-2486.2007.01415.x, 2007.
Dambrine, E., Carisey, N., Pollier, B., and Granier, A.: Effects of drought on the yellowing status and the dynamics of mineral elements in the xylem sap of declining spruce (Picea abies L.), Plant Soil, 150, 303–306, https://doi.org/10.1007/BF00013028, 1993.
Dambrine, E., Pollier, B., Poszwa, A., Ranger, J., Probst, A., Viville, D., Biron, P., and Granier, A.: Evidence of current soil acidification in spruce stands in the Vosges Mountains, north-eastern France, Water Air Soil Pollut., 105, 43–52, https://doi.org/10.1023/A:1005030331423, 1998.
Deng, L., Peng, C., Kim, D.-G., Li, J., Liu, Y., Hai, X., Liu, Q., Huang, C., Shangguan, Z., and Kuzyakov, Y.: Drought effects on soil carbon and nitrogen dynamics in global natural ecosystems, Earth Sci. Rev., 214, 103501, https://doi.org/10.1016/j.earscirev.2020.103501, 2021.
De Wit, H. A., Mulder, J., Nygaard, P. H., and Aamlid, D.: Testing the aluminium toxicity hypothesis: a field manipulation experiment in mature spruce forest in Norway, in: Acid rain 2000, edited by: Satake, K., Shindo, J., Takamatsu, T., Nakano, T., Aoki, S., Fukuyama, T., Hatakeyama, S., Ikuta, K., Kawashima, M., Kohno, Y., Kojima, S., Murano, K., Okita, T., Taoda, H., Tsunoda, K., and Tsurumi, M., Springer Netherlands, Dordrecht, 995–1000, https://doi.org/10.1007/978-94-007-0810-5_13, 2001.
Drever, J. and Clow, D.: Weathering rates in catchments, in: Chemical Weathering Rates of Silicate Minerals, edited by: White, A. F. and Brantley, S. L., De Gruyter, Berlin, Boston, 463–484, https://doi.org/10.1515/9781501509650-012, 1995.
Ditmarova, L., Kurjak, D., Palmroth, S., Kmet, J., and Strelcova, K.: Physiological responses of Norway spruce (Picea abies) seedlings to drought stress, Tree Physiol., 30, 205–213, https://doi.org/10.1093/treephys/tpp116, 2010.
Eisalou, H.K., Şengönül, K., Gökbulak, F., Serengil, Y., and Uygur, B.: Effects of forest canopy cover and floor on chemical quality of water in broad leaved and coniferous forests of Istanbul, Turkey, For. Ecol. Manage., 289, 371–377, https://doi.org/10.1016/j.foreco.2012.10.031, 2013
FAO: Global Forest Resources Assessment 2020: Main report, Rome, FRA [data set], https://doi.org/10.4060/ca9825en, 2020
Faranda, D., Pascale, S., and Bulut, B.: Persistent anticyclonic conditions and climate change exacerbated the exceptional 2022 European-Mediterranean drought, Environ. Res. Lett., 18, 034030, https://doi.org/10.1088/1748-9326/acbc37, 2023.
Fest, E. P. M. J., Temminghoff, E. J. M., Griffioen, J., and Van Riemsdijk, W. H.: Proton buffering and metal leaching in sandy soils, Environ. Sci. Technol., 39, 7901–7908, https://doi.org/10.1021/es0505806, 2005.
Fichter, J., Dambrine, E., Turpault, M.-P., and Ranger, J.: Base cation supply in spruce and beech ecosystems of the Strengbach catchment (Vosges Mountains, N-E France), Water Air Soil Pollut., 104, 125–148, https://doi.org/10.1023/A:1004966302517, 1997.
Forzieri, G., Dutrieux, L. P., Elia, A., Eckhardt, B., Caudullo, G., Taboada, F. Á., Andriolo, A., Bălăcenoiu, F., Bastos, A., Buzatu, A., Dorado, F. C., Dobrovolný, L., Duduman, M., Fernandez-Carrillo, A., Hernández-Clemente, R., Hornero, A., Ionu?, S., Lombardero, M. J., Junttila, S., Lukeš, P., Marianelli, L., Mas, H., Mlčoušek, M., Mugnai, F., Ne?oiu, C., Nikolov, C., Olenici, N., Olsson, P., Paoli, F., Paraschiv, M., Patočka, Z., Pérez-Laorga, E., Quero, J. L., Rüetschi, M., Stroheker, S., Nardi, D., Ferenčík, J., Battisti, A., Hartmann, H., Nistor, C., Cescatti, A., and Beck, P. S. A.: The Database of European Forest Insect and Disease Disturbances: DEFID2, Global Change Biology, 29, 6040–6065, https://doi.org/10.1111/gcb.16912, 2023.
Frank, S. D.: Review of the direct and indirect effects of warming and drought on scale insect pests of forest systems, Forestry: An International, Journal of Forest Research, 94, 167–180, https://doi.org/10.1093/forestry/cpaa033, 2021.
Gangloff, S., Stille, P., Schmitt, A.-D., and Chabaux, F.: Impact of bacterial activity on Sr and Ca isotopic compositions (87Sr 86Sr and δ44 40Ca) in soil solutions (the Strengbach CZO), Procedia Earth and Planet. Sci., 10, 109–113, https://doi.org/10.1016/j.proeps.2014.08.038, 2014.
Gaul, D., Hertel, D., Borken, W., Matzner, E., and Leuschner, C.: Effects of experimental drought on the fine root system of mature Norway spruce, For. Ecol. Manage., 256, 1151–1159, https://doi.org/10.1016/j.foreco.2008.06.016, 2008.
Gehring, C. A., Swaty, R. L., and Deckert, R. J.: Mycorrhizas, drought, and host-plant mortality, in: Mycorrhizal mediation of soil, Elsevier, 279–298, https://doi.org/10.1016/B978-0-12-804312-7.00016-4, 2017.
Gessler, A., Schaub, M., and McDowell, N.G.: The role of nutrients in drought-induced tree mortality and recovery, New Phytol., 214, 513–520, https://doi.org/10.1111/nph.14340, 2017.
Gharun, M., Shekhar, A., Xiao, J., Li, X., and Buchmann, N.: Effect of the 2022 summer drought across forest types in Europe, Biogeosciences, 21, 5481–5494, https://doi.org/10.5194/bg-21-5481-2024, 2024.
Goddéris, Y., François, L.M., Probst, A., Schott, J., Moncoulon, D., Labat, D., and Viville, D.: Modelling weathering processes at the catchment scale: The WITCH numerical model, Geochimica et Cosmochimica Acta, 70, 1128–1147, https://doi.org/10.1016/j.gca.2005.11.018, 2006.
Gomez, D. F., Riggins, J. J., and Cognato, A. I.: Bark beetles, in: Forest entomology and pathology, edited by: Allison, D. J., Paine, T. D., Slippers, B., and Wingfield, M. J., Springer International Publishing, Cham, 299–337, https://doi.org/10.1007/978-3-031-11553-0_10, 2023.
Granier, A., Bréda, N., Biron, P., and Villette, S.: A lumped water balance model to evaluate duration and intensity of drought constraints in forest stands, Ecological Modelling, 116, 269–283, https://doi.org/10.1016/S0304-3800(98)00205-1, 1999.
Grunsky, E. C.: The interpretation of geochemical survey data, GEEA, 10, 27–74, https://doi.org/10.1144/1467-7873/09-210, 2010.
Haines, B. L., Waide, J. B., and Todd, R. L.: Soil Solution Nutrient Concentrations Sampled with Tension and Zero-Tension Lysimeters: Report of Discrepancies, Soil Science Soc. Amer. J., 46, 658–661, https://doi.org/10.2136/sssaj1982.03615995004600030042x, 1982.
Hartmann, H., Bastos, A., Das, A. J., Esquivel-Muelbert, A., Hammond, W. M., Martínez-Vilalta, J., McDowell, N. G., Powers, J. S., Pugh, T. A. M., Ruthrof, K. X., and Allen, C. D.: Climate Change Risks to Global Forest Health: Emergence of Unexpected Events of Elevated Tree Mortality Worldwide, Annual Review of Plant Biology, 73, 673–702, https://doi.org/10.1146/annurev-arplant-102820-012804, 2022.
Hentschel, K., Borken, W., and Matzner, E.: Leaching losses of inorganic N and DOC following repeated drying and wetting of a spruce forest soil, Plant Soil, 300, 21–34, https://doi.org/10.1007/s11104-007-9385-3, 2007.
Hesse, B. D., Hikino, K., Gebhardt, T., Buchhart, C., Dervishi, V., Goisser, M., Pretzsch, H., Häberle, K.-H., and Grams, T. E. E.: Acclimation of mature spruce and beech to five years of repeated summer drought – The role of stomatal conductance and leaf area adjustment for water use, Sci. Total Environ., 951, 175805, https://doi.org/10.1016/j.scitotenv.2024.175805, 2024.
Houle, D., Lajoie, G., and Duchesne, L.: Major losses of nutrients following a severe drought in a boreal forest, Nat. Plants, 2, 16187, https://doi.org/10.1038/nplants.2016.187, 2016.
Huguet, A., Vacher, L., Relexans, S., Saubusse, S., Froidefond, J. M., and Parlanti, E.: Properties of fluorescent dissolved organic matter in the Gironde Estuary, Org. Geochem., 40, 706–719, https://doi.org/10.1016/j.orggeochem.2009.03.002, 2009.
Institut national de l'information géographique et forestière (IGN): Mémento 2024, Inventaire forestier national, https://inventaire-forestier.ign.fr (last access: 18 September 2025), 2024.
Ingestad, T.: Mineral Nutrient Requirements of Pinus silvestris and Picea abies Seedlings, Physiol. Plant., 45, 373–380, https://doi.org/10.1111/j.1399-3054.1979.tb02599.x, 1979.
Knight, C. G., Nicolitch, O., Griffiths, R. I., Goodall, T., Jones, B., Weser, C., Langridge, H., Davison, J., Dellavalle, A., Eisenhauer, N., Gongalsky, K. B., Hector, A., Jardine, E., Kardol, P., Maestre, F. T., Schädler, M., Semchenko, M., Stevens, C., Tsiafouli, M. A., Vilhelmsson, O., Wanek, W., and De Vries, F. T.: Soil microbiomes show consistent and predictable responses to extreme events, Nature, 636, 690–696, https://doi.org/10.1038/s41586-024-08185-3, 2024.
Knutzen, F., Averbeck, P., Barrasso, C., Bouwer, L. M., Gardiner, B., Grünzweig, J. M., Hänel, S., Haustein, K., Johannessen, M. R., Kollet, S., Müller, M. M., Pietikäinen, J.-P., Pietras-Couffignal, K., Pinto, J. G., Rechid, D., Rousi, E., Russo, A., Suarez-Gutierrez, L., Veit, S., Wendler, J., Xoplaki, E., and Gliksman, D.: Impacts on and damage to European forests from the 2018–2022 heat and drought events, Nat. Hazards Earth Syst. Sci., 25, 77–117, https://doi.org/10.5194/nhess-25-77-2025, 2025.
Kopáček, J., Turek, J., Hejzlar, J., and Šantrůčková, H.: Canopy leaching of nutrients and metals in a mountain spruce forest, Atmos. Environ., 43, 5443–5453, https://doi.org/10.1016/j.atmosenv.2009.07.031, 2009.
Kronnäs, V., Lucander, K., Zanchi, G., Stadlinger, N., Belyazid, S., and Akselsson, C.: Effect of droughts and climate change on future soil weathering rates in Sweden, Biogeosciences, 20, 1879–1899, https://doi.org/10.5194/bg-20-1879-2023, 2023.
Krüger, M., Potthast, K., Michalzik, B., Tischer, A., Küsel, K., Deckner, F. F. K., and Herrmann, M.: Drought and rewetting events enhance nitrate leaching and seepage-mediated translocation of microbes from beech forest soils, Soil Biol. Biochem., 154, 108153, https://doi.org/10.1016/j.soilbio.2021.108153, 2021.
Kump, L. R., Brantley, S. L., and Arthur, M. A.: Chemical weathering, atmospheric CO2, and climate, Annu. Rev. Earth Planet. Sci., 28, 611–667, https://doi.org/10.1146/annurev.earth.28.1.611, 2000.
Kuypers, M. M. M., Marchant, H. K., and Kartal, B.: The microbial nitrogen-cycling network, Nat. Rev. Microbiol., 16, 263–276, https://doi.org/10.1038/nrmicro.2018.9, 2018.
Lamersdorf, N. P., Beier, C., Blanck, K., Bredemeier, M., Cummins, T., Farrell, E. P., Kreutzer, K., Rasmussen, L., Ryan, M., Weis, W., and Xu, Y.-J.: Effect of drought experiments using roof installations on acidification/nitrification of soils, Forest Ecology and Management, 101, 95–109, https://doi.org/10.1016/S0378-1127(97)00128-X, 1998.
Lasaga, A. C., Soler, J. M., Ganor, J., Burch, T. E., and Nagy, K. L.: Chemical weathering rate laws and global geochemical cycles, Geochimica et Cosmochimica Acta, 58, 2361–2386, https://doi.org/10.1016/0016-7037(94)90016-7 , 1994.
Leigh, R.A. and Wyn Jones, R.G.: A hypothesis relating critical potassium concentrations for growth to the distribution and functions of this ion in the plant cell, New Phytol., 97, 1–13, https://doi.org/10.1111/j.1469-8137.1984.tb04103.x, 1984.
Lévesque, M., Walthert, L., and Weber, P.: Soil nutrients influence growth response of temperate tree species to drought, Journal of Ecology, 104, 377–387, https://doi.org/10.1111/1365-2745.12519, 2016.
Lovett, G. M. and Lindberg, S. E.: Dry deposition and canopy exchange in a mixed Oak forest as determined by analysis of throughfall, J. Appl. Ecol., 21, 1013, https://doi.org/10.2307/2405064, 1984.
Lu, P., Biron, P., Bréda, N., and Granier, A.: Water relations of adult Norway spruce (Picea abies (L) Karst) under soil drought in the Vosges mountains: water potential, stomatal conductance and transpiration, Ann. For. Sci., 52, 117–129, https://doi.org/10.1051/forest:19950203, 1995.
Marques, R., Ranger, J., Gelhaye, D., Pollier, B., Ponette, Q., and Gœdert, O.: Comparison of chemical composition of soil solutions collected by zero-tension plate lysimeters with those from ceramic-cup lysimeters in a forest soil, European J. Soil Science, 47, 407–417, https://doi.org/10.1111/j.1365-2389.1996.tb01414.x, 1996.
McKnight, D. M., Boyer, E. W., Westerhoff, P. K., Doran, P. T., Kulbe, T., and Andersen, D. T.: Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity, Limnol. Oceanogr., 46, 38–48, https://doi.org/10.4319/lo.2001.46.1.0038, 2001.
Meng, C., Tian, D., Zeng, H., Li, Z., Yi, C., and Niu, S.: Global soil acidification impacts on belowground processes, Environ. Res. Lett., 14, 074003, https://doi.org/10.1088/1748-9326/ab239c, 2019.
Muhr, J., Goldberg, S.D., Borken, W., and Gebauer, G.: Repeated drying–rewetting cycles and their effects on the emission of CO2, N2O, NO, and CH4 in a forest soil, Z. Pflanzenernähr. Bodenk., 171, 719–728, https://doi.org/10.1002/jpln.200700302, 2008.
Nilsen, P.: Effect of nitrogen on drought strain and nutrient uptake in Norway spruce Picea abies (L.) Karst. trees, Plant Soil, 172, 73–85, https://doi.org/10.1007/BF00020861, 1995.
Oksanen, J., Simpson, G. L., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O'Hara, R. B., Solymos, P., Stevens, M. H. H., Szoecs, E., Wagner, H., Barbour, M., Bedward, M., Bolker, B., Borcard, D., Carvalho, G., Chirico, M., De Caceres, M., Durand, S., Evangelista, H. B. A., FitzJohn, R., Friendly, M., Furneaux, B., Hannigan, G., Hill, M. O., Lahti, L., McGlinn, D., Ouellette, M.-H., Ribeiro Cunha, E., Smith, T., Stier, A., Ter Braak, C. J. F., Weedon, J., and Borman, T.: vegan: Community Ecology Package, R package version 2.8-0, https://doi.org/10.32614/CRAN.package.vegan, 2001.
Oursin, M., Pierret, M.-C., Beaulieu, É., Daval, D., and Legout, A.: Is there still something to eat for trees in the soils of the Strengbach catchment?, For. Ecol. Manage., 527, 120583, https://doi.org/10.1016/j.foreco.2022.120583, 2023.
Ovington, J. D.: The calcium and magnesium contents of tree species grown in close stands, New Phytologist, 58, 164–175, https://doi.org/10.1111/j.1469-8137.1959.tb05348.x, 1959.
Parlanti, E., Wörz, K., Geoffroy, L., and Lamotte, M.: Dissolved organic matter fluorescence spectroscopy as a tool to estimate biological activity in a coastal zone submitted to anthropogenic inputs, Org. Geoch., 31, 1765–1781, https://doi.org/10.1016/S0146-6380(00)00124-8, 2000.
Persson, H., Von Fircks, Y., Majdi, H., and Nilsson, L. O.: Root distribution in a Norway spruce (Picea abies (L.) Karst.) stand subjected to drought and ammonium-sulphate application, in: Nutrient uptake and cycling in forest ecosystems, edited by: Nilsson, L. O., Hüttl, R. F., and Johansson, U. T., Developments in Plant and Soil Sciences, 62, Springer, Dordrecht, https://doi.org/10.1007/978-94-011-0455-5_17, 1995.
Pierret, M.-C., Cotel, S., Ackerer, P., Beaulieu, E., Benarioumlil, S., Boucher, M., Boutin, R., Chabaux, F., Delay, F., Fourtet, C., Friedmann, P., Fritz, B., Gangloff, S., Girard, J.-F., Legtchenko, A., Viville, D., Weill, S., and Probst, A.: The Strengbach catchment: a multidisciplinary environmental sentry for 30 years, Vadose Zone Journal, 17, 1–17, https://doi.org/10.2136/vzj2018.04.0090, 2018.
Pierret, M.-C., Viville, D., Dambrine, E., Cotel, S., and Probst, A.: Twenty-five year record of chemicals in open field precipitation and throughfall from a medium-altitude forest catchment (Strengbach – NE France): An obvious response to atmospheric pollution trends, Atmospheric Environment, 202, 296–314, https://doi.org/10.1016/j.atmosenv.2018.12.026, 2019.
Ponette-González, A. G., Van Stan II, J. T., and Magyar, D.: Things seen and unseen in Throughfall and stemflow, in: Precipitation partitioning by vegetation, edited by: Van Stan, II, J., Gutmann, E., and Friesen, J., Springer, Cham., https://doi.org/10.1007/978-3-030-29702-2_5, 2020.
Poszwa, A., Wickman, T., Dambrine, E., Ferry, B., Dupouey, J.-L., Helle, G., Schleser, G., and Breda, N.: A Retrospective isotopic study of spruce decline in the Vosges Mountains (France), Water, Air and Soil Pollution: Focus, 3, 201–222, https://doi.org/10.1023/A:1022176025379, 2003.
Probst, A., Dambrine, E., Viville, D., and Fritz, B.: Influence of acid atmospheric inputs on surface water chemistry and mineral fluxes in a declining spruce stand within a small catchment (Vosges massif, France), J. Hydrol., 116, 101–124, https://doi.org/10.1016/0022-1694(90)90118-H, 1990.
Probst, A., Viville, D., Fritz, B., Ambroise B., and Dambrine E.: Hydrochemical budgets of a small forested granitic catchment exposed to acid deposition: The strengbach catchment case study (Vosges massif, France), Water Air Soil Pollut, 62, 337–347, https://doi.org/10.1007/BF00480265, 1992.
Probst, A., Fritz, B., and Viville, D.: Mid-term trends in acid precipitation, streamwater chemistry and element budgets in the Strengbach catchment (Vosges mountains, France), Water, Air and Soil Pollution, 79, 39–59, https://doi.org/10.1007/BF01100429, 1995.
Quirion, B. R., Domke, G. M., Walters, B. F., Lovett, G. M., Fargione, J. E., Greenwood, L., Serbesoff-King, K., Randall, J. M., and Fei, S.: Insect and Disease Disturbances Correlate With Reduced Carbon Sequestration in Forests of the Contiguous United States, Front. For. Glob. Change, 4, 716582, https://doi.org/10.3389/ffgc.2021.716582, 2021.
Ranger, J., Discours, D., Mohamed Ahamed, D., Moares, C., Dambrine, É., Merlet, D., and Rouiller, J.: Comparaison des eaux liées et des eaux libres des sols de 3 peuplements d'épicéa (Picea abies Karst) des Vosges. Application à l'étude du fonctionnement actuel des sols et conséquences pour l'état sanitaire des peuplements, Ann. For. Sci., 50, 425–444, https://doi.org/10.1051/forest:19930502, 1993.
Rüdinger, M., Hallgren, S. W., Steudle, E., and Schulze, E.-D.: Hydraulic and osmotic properties of spruce roots, J. Exp. Bot., 45, 1413–1425, https://doi.org/10.1093/jxb/45.10.1413, 1994.
Rodríguez-Calcerrada, J., Li, M., López, R., Cano, F. J., Oleksyn, J., Atkin, O. K., Pita, P., Aranda, I., and Gil, L.: Drought-induced shoot dieback starts with massive root xylem embolism and variable depletion of nonstructural carbohydrates in seedlings of two tree species, New Phytol., 213, 597–610, https://doi.org/10.1111/nph.14150, 2017.
Saintonge, F.-X., Gillette, M., Blaser, S., Queloz, V., and Leroy, Q.: Situation et gestion de la crise liée aux scolytes de l'Épicéa commun fin 2021 dans l'est de la France, en Suisse et en Wallonie, Revue forestière française, 73, 619–641, https://doi.org/10.20870/revforfr.2021.7201, 2022.
San-Miguel-Ayanz, J., Durrant, T., Boca, R., Maianti, P., Libertà, G., Jacome Felix Oom, D., Branco, A., De Rigo, D., Suarez-Moreno, M., Ferrari, D., Roglia, E., Scionti, N., Broglia, M., Onida, M., Tistan, A., and Loffler, P.: Forest fires in Europe, Middle East and North Africa 2022, Publications Office of the European Union, Luxembourg, 2023, EFFIS [data set], https://doi.org/10.2760/871593, 2023.
Sardans, J. and Peñuelas, J.: Potassium: a neglected nutrient in global change, Global Ecology and Biogeography, 24, 261–275, https://doi.org/10.1111/geb.12259, 2015.
Sardans, J., Peñuelas, J., and Ogaya, R.: Drought's impact on Ca, Fe, Mg, Mo and S concentration and accumulation patterns in the plants and soil of a Mediterranean evergreen Quercus ilex forest, Biogeochemistry, 87, 49–69, https://doi.org/10.1007/s10533-007-9167-2, 2008.
Schimel, J., Balser, T. C., and Wallenstein, M.: Microbial stress-response physiology and its implications for ecosystem function, Ecology, 88, 1386–1394, https://doi.org/10.1890/06-0219, 2007.
Schlesinger, W. H., Dietze, M. C., Jackson, R. B., Phillips, R. P., Rhoades, C. C., Rustad, L. E., and Vose, J. M.: Forest biogeochemistry in response to drought, Global Change Biol., 22, 2318–2328, https://doi.org/10.1111/gcb.13105, 2016.
Schmid, I. and Kazda, M.: Root distribution of Norway spruce in monospecific and mixed stands on different soils, For. Ecol. Manage., 159, 37–47, https://doi.org/10.1016/S0378-1127(01)00708-3, 2002.
Schmied, G., Hilmers, T., Mellert, K.-H., Uhl, E., Buness, V., Ambs, D., Steckel, M., Biber, P., Šeho, M., Hoffmann, Y.-D., and Pretzsch, H.: Nutrient regime modulates drought response patterns of three temperate tree species, Sci Total Environ., 868, 161601, https://doi.org/10.1016/j.scitotenv.2023.161601, 2023.
Schulze, K., Borken, W., and Matzner, E.: Dynamics of dissolved organic 14C in throughfall and soil solution of a Norway spruce forest, Biogeochemistry, 106, 461–473, https://doi.org/10.1007/s10533-010-9526-2, 2011.
Schwertman, N. C., Owens, M. A., and Adnan, R.: A simple more general boxplot method for identifying outliers, Computational Statistics and Data Analysis, 47, 165–174, https://doi.org/10.1016/j.csda.2003.10.012, 2004.
Senf, C., Buras, A., Zang, C. S., Rammig, A., and Seidl, R.: Excess forest mortality is consistently linked to drought across Europe, Nat. Commun., 11, 6200, https://doi.org/10.1038/s41467-020-19924-1, 2020.
Serène, L., Mazzilli, N., Batiot-Guilhe, C., Emblanch, C., Gillon, M., Babic, M., Dupont, J., Simler, R., and Blanc, M.: Questioning calculation and interpretation of fluorescence indices in natural waters studies, J. Hydrol., 650, 132524, https://doi.org/10.1016/j.jhydrol.2024.132524, 2025.
Setia, R., Lamba, S., Chander, S., Kumar, V., Singh, R., Litoria, P. K., Singh, R. P., and Pateriya, B.: Spatio-temporal variations in water quality, hydrochemistry and its controlling factors in a perennial river in India, Appl. Water Sci., 11, 169, https://doi.org/10.1007/s13201-021-01504-3, 2021.
Singh, G., Kaur, G., Williard, K., Schoonover, J., and Kang, J.: Monitoring of Water and Solute Transport in the Vadose Zone: A Review, Vadose Zone Journal, 17, 1–23, https://doi.org/10.2136/vzj2016.07.0058, 2018
Sorel, M., Mittelberger, S., Berne, C., and Ribes, A.: Bilan climatique de l'année 2022, La Météorologie, 120, 2–4, https://doi.org/10.37053/lameteorologie-2023-0002, 2023.
Soubeyroux, J.-M., Bernus, S., Core, L., Drouin, A., Dubuisson, B., Etchevers, P., Gouget, V., Josse, P., Kerdoncuff, M., Samacoits, R., and Tocquer, F.: Les nouvelles projections climatiques de référence Drias 2020 pour la métropole, http://www.drias-climat.fr/document/rapport-DRIAS-2020-red3-2.pdf (last access: 19 August 2025), 2021.
Strobel, B. W., Hansen, H. C. B., Borggaard, O. K., Andersen, M. K., and Raulund-Rasmussen, K.: Composition and reactivity of DOC in forest floor soil solutions in relation to tree species and soil type, Biogeochemistry, 56, 1–26, https://doi.org/10.1023/A:1011934929379, 2001.
Strohmenger, L., Ackerer, P., Belfort, B., and Pierret, M. C.: Local and seasonal climate change and its influence on the hydrological cycle in a mountainous forested catchment, J. Hydrol., 610, 127914, https://doi.org/10.1016/j.jhydrol.2022.127914, 2022.
Swift, R. S.: Organic matter characterization, in: Methods of soil analysis, edited by: Sparks, D. L., Page, A. L., Helmke, P. A., Loeppert, R. H., Soltanpour, P. N., Tabatabai, M. A., Johnston, C. T., and Sumner, M. E., https://doi.org/10.2136/sssabookser5.3.c35, 1996.
Tiwari, T., Sponseller, R. A., and Laudon, H.: The emerging role of drought as a regulator of dissolved organic carbon in boreal landscapes, Nat. Commun., 13, 5125, https://doi.org/10.1038/s41467-022-32839-3, 2022.
Touche, J., Calvaruso, C., De Donato, P., and Turpault, Mp.: Five successive years of rainfall exclusion induce nutritional stress in a mature beech stand, For. Ecol. Manage., 507, 119987, https://doi.org/10.1016/j.foreco.2021.119987, 2022.
Touche, J., Calvaruso, C., De Donato, P., and Turpault, M.-P.: Drought events influence nutrient canopy exchanges and green leaf partitioning during senescence in a deciduous forest, Forest Ecosystems, 11, 100173, https://doi.org/10.1016/j.fecs.2024.100173, 2024.
Ulrich, B.: Interaction of forest canopies with atmospheric constituents: SO2, alkali and earth alkali cations and chloride, in: Effects of Accumulation of Air Pollutants in Forest Ecosystems, edited by: Ulrich, B. and Pankrath, J., Springer, Dordrecht, 33–45, https://doi.org/10.1007/978-94-009-6983-4_2, 1983.
Ulrich, B., Mayer, R., and Khanna, P. K.: Chemical changes due to acid precipitation in a loess-derived soil in central Europe, Soil Sci., 130, 193–199, https://doi.org/10.1097/00010694-198010000-00005, 1980.
Vose, J. M., Miniat, C. F., Luce, C. H., Asbjornsen, H., Caldwell, P. V., Campbell, J. L., Grant, G. E., Isaak, D. J., Loheide, S. P., and Sun, G.: Ecohydrological implications of drought for forests in the United States, For. Ecol. Manage., 380, 335–345, https://doi.org/10.1016/j.foreco.2016.03.025, 2016.
Wagner, Y., Volkov, M., Nadal-Sala, D., Ruehr, N. K., Hochberg, U., and Klein, T.: Relationships between xylem embolism and tree functioning during drought, recovery, and recurring drought in Aleppo pine, Physiol. Plant., 175, 13995, https://doi.org/10.1111/ppl.13995, 2023.
Watmough, S. A., Koseva, I., and Landre, A.: A Comparison of Tension and Zero-Tension Lysimeter and PRS™ Probes for Measuring Soil Water Chemistry in Sandy Boreal Soils in the Athabasca Oil Sands Region, Canada, Water Air Soil Pollut, 224, 1663, https://doi.org/10.1007/s11270-013-1663-5, 2013.
Weed, A. S., Ayres, M. P., and Hicke, J. A.: Consequences of climate change for biotic disturbances in North American forests, Ecol. Monogr., 83, 441–470, https://doi.org/10.1890/13-0160.1, 2013.
Wilhite, D. A. and Glantz, M. H.: Understanding: the Drought Phenomenon: The Role of Definitions, Water International, 10, 111–120, https://doi.org/10.1080/02508068508686328, 1985.
Winter, C., Nguyen, T. V., Musolff, A., Lutz, S. R., Rode, M., Kumar, R., and Fleckenstein, J. H.: Droughts can reduce the nitrogen retention capacity of catchments, Hydrol. Earth Syst. Sci., 27, 303–318, https://doi.org/10.5194/hess-27-303-2023, 2023.
Winter, C., Müller, S., Kattenborn, T., Stahl, K., Szillat, K., Weiler, M., and Schnabel, F.: Forest Dieback in Drinking Water Protection Areas – A Hidden Threat to Water Quality, Earth's Future, 13, e2025EF006078, https://doi.org/10.1029/2025EF006078, 2025.
Woods, C., Hunt, S., Morris, D., and Gordon, A.: Epiphytes influence the transformation of nitrogen in coniferous forest canopies, Boreal Environ. Res., 17, 411–424, 2012.
Zsolnay, A., Baigar, E., Jimenez, M., Steinweg, B., and Saccomandi, F.: Differentiating with fluorescence spectroscopy the sources of dissolved organic matter in soils subjected to drying, Chemosphere, 38, 45–50, https://doi.org/10.1016/S0045-6535(98)00166-0, 1999.