the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 04 Mar 2026
| 04 Mar 2026
Tropical wet season runoff mobilises younger carbon in rainforest streams but older carbon in agricultural streams
Vanessa Solano
Dioni I. Cendón
Francesco Ulloa-Cedamanos
Liza K. McDonough
Robert G. M. Spencer
Niels C. Munksgaard
Lindsay B. Hutley
Jean-Sébastien Moquet
David E. Butman
Knowledge of the age of organic carbon (C) that is leached from soils to streams is key to understanding how C is mobilised within ecosystems. The tropics are characterised by significant C fluxes through streams, yet the time scales of organic C sequestration and export remain uncertain in these regions. Here we examined the concentration, composition and age of dissolved organic C (DOC) in 18 small mountainous catchments of the Australian humid tropics, including six rainforest and 12 agricultural catchments, sampled during both the dry and wet seasons. We found that DOC ages varied widely across sites but were generally centuries to millennia old (median ± standard deviation 1553±848 years BP), with no consistent differences between rainforest and agricultural catchments. However, the two land use categories diverged in their responses to high flow conditions, with DOC age in rainforest streams tending to decrease from 1878±604 years BP in the dry season to 708±791 years BP in the wet season, whereas agricultural streams mobilised similarly aged or older DOC in the wet season (1728±641 years BP) than in the dry season (1303±1036 years BP). A subset of dissolved inorganic C (DIC) samples collected from three of the catchments (both rainforest and agricultural) indicated that DIC was mostly modern (123±136 years BP) and always younger than DOC. These differences in DIC and DOC ages suggest a partial decoupling between DOC and DIC export pathways, with DOC derived from older soil C pools, while DIC reflected recent C inputs from vegetation uptake and decomposition. Our results highlight the importance of seasonal shifts in the age of C export and the need to conduct sampling that encompasses seasonality in human-impact studies to better constrain C pools and sinks.
- Article
(4661 KB) - Full-text XML
-
Supplement
(2129 KB) - BibTeX
- EndNote




Soils represent one of the largest reservoirs of organic carbon (C) and play a crucial role in the global terrestrial C sink. Yet the stability of the soil C pool is highly vulnerable to anthropogenic disturbance, including urban and agricultural development (Don et al., 2011; Schlesinger, 1986; Xenopoulos et al., 2021; Hobley et al., 2017). Soil C loss not only occurs during the process of land use change, when carbon dioxide (CO2) is released to the atmosphere, but also with changes in the leakage of organic C into streams and rivers as dissolved organic C (DOC) (e.g. Coble et al., 2022; Wilson and Xenopoulos, 2008; Drake et al., 2020). While most riverine DOC is modern, sourced from recent assimilation of CO2 by vegetation (e.g. Mayorga et al., 2005; Marwick et al., 2015; Billett et al., 2007), land conversion to agriculture can cause old soil organic C to re-enter the contemporary C cycle and be exported via streams and rivers (Butman et al., 2015; Evans et al., 2014; Moore et al., 2013). Conversion from natural to agricultural land use has been linked to increased soil erosion and to the alteration of surface and subsurface flow paths and water residence times following disturbance (Barnes et al., 2018; Butman et al., 2015; Evans et al., 2022; Drake et al., 2020, 2019).
Tropical streams and rivers are a globally significant hotspot for terrestrial C export (Liu et al., 2022, 2024; Rocher-Ros et al., 2023; Battin et al., 2023), yet knowledge of the origin and age of C export from tropical ecosystems remains limited (Dean et al., 2025). Early work in the Amazon (Mayorga et al., 2005) and Congo (Spencer et al., 2012) revealed that the DOC transported by these two major rivers was predominantly modern. Mayorga et al. (2005) showed that this young DOC was the dominant source of dissolved inorganic C (DIC) and CO2 degassing. Marwick et al. (2015) also consistently found modern DOC in a range of African rivers. The dominance of young DOC has also been highlighted in smaller river systems, including streams draining undisturbed peatland forest in Borneo (Moore et al., 2013; Müller et al., 2015) and pristine rainforest in the upper Congo Basin (Drake et al., 2019). However, anthropogenic disturbance has emerged as a key control on DOC age in small tropical systems. Studies have shown that in catchments where land has been cleared for agriculture, exported DOC can be centuries to millennia old (Moyer et al., 2013; Drake et al., 2019; Moore et al., 2013; Waldron et al., 2019).
Despite improved understanding of the role of land use change on DOC age in tropical streams and rivers, observations remain limited in both space and time, and the extent to which the age of riverine DOC is affected by changes in flow conditions remains unknown. The tropics are affected by strong rainfall seasonality, and the bulk of C export is likely to occur during the wetter months, yet few studies have examined DOC age under high flow conditions and across varying flow regimes. Marwick et al. (2015) reported younger DOC during the wet season in the Athi River (Kenya), attributing this to the flushing of young organic C accumulated during the dry season. Likewise, Chen et al. (2024) found younger DOC values during a large flood of the subtropical Yangtze River compared to lower flow conditions. Similar patterns have been reported in northern high-latitude and temperate regions, where high hydrological connectivity and shallow subsurface flow paths tend to mobilise younger DOC (Tittel et al., 2022; Campeau et al., 2019; Barnes et al., 2018; Aiken et al., 2014). In contrast, Moore et al. (2013) found increasing DOC ages with wet conditions in disturbed peatland catchments of Borneo. These contrasting findings highlight the remaining uncertainties regarding the combined effects of land use and flow conditions on the age of transported DOC in tropical streams. Also unknown is the relationship between DOC and DIC age in these systems. Studies have reported a decoupling between DOC and DIC ages, with generally modern compared to older and more variable DOC (Campeau et al., 2019; Moyer et al., 2013). Such patterns have been attributed to being directly derived from recent soil CO2 inputs from root respiration rather than from the decomposition of older DOC (Campeau et al., 2019), a hypothesis that needs to be further tested.
The humid tropics of Australia is a mountainous region that drains to the Great Barrier Reef lagoon. Agricultural development in the region began in the 1870s, with rainforest cleared for sugarcane and pasture development (Kemp et al., 2007). As a result, coastal ecosystems have been impacted by poor water quality, which has intensified in the past few decades (Kroon et al., 2016, 2012; Guo et al., 2025). While suspended sediment and nutrient loads have been particularly scrutinised, few studies have investigated C export from rivers of the region (Rosentreter and Eyre, 2020), and none have investigated the age of exported C. In this paper, we aim to assess the impact of land disturbance and flow conditions on the age of DOC exported by streams across the Australian humid tropics. We examined the characteristics, concentration and isotopic composition (carbon-13; δ13C and radiocarbon; 14C) of bulk DOC in 18 small catchments (six relatively pristine rainforest and 12 dominated by agriculture) during both the dry and wet seasons. Additionally, we measured the age of soil organic C in two shallow soil cores and the age of DIC in a small subset (three) of the catchments. We addressed the following questions:
-
Is there a significant difference between the age of DOC transported in streams draining rainforest catchments versus that of agricultural catchments? We hypothesised that natural land conversion to agricultural results in the export of older DOC, as increased erosion reduces the contribution of modern soil organic C to exported DOC.
-
Does the age of DOC transported in streams alter with changing flow conditions? We hypothesised that DOC is younger at high flow than under baseflow conditions, as C from shallower soil layers tends to be mobilised when water tables rise.
-
Is the age of DIC comparable to the age of DOC? We hypothesised that DIC is younger than DOC, due to external inputs of recent soil CO2 and the preferential mineralisation of biolabile organic matter (while recognising that DOC age and biolability do not always covary).
2.1 Study area
The 18 stream sites are located within the humid tropics of Far North Queensland, Australia (Fig. 1; Table 1). The region spans a range of climate types, from Af (tropical rainforest) and Am (tropical monsoon) in the lowlands (0–500 m a.s.l.) to Cfa (humid subtropical) at higher elevations (500–1000 m a.s.l.) (Beck et al., 2023). Annual rainfall in the region ranges between approximately 1500 and 4000 mm and follows a strong seasonal pattern, with 65 %–75 % falling during the wetter months (November to April) and 25 %–35 % during the drier months (May to October) (Bureau of Meteorology, 2025). The geology of the region is diverse, with the coastal areas to the East dominated by Quaternary alluvial deposits, while the mountain range and Atherton Tableland to the West are characterised by Cenozoic and Palaeozoic basaltic lava flows and metamorphic rocks. The area does not feature any significant organic-rich or carbonate-rich sedimentary formations (Jell, 2013).

Figure 1Location of the 18 study catchments. Catchments delineated in green represent rainforest (R) while those shaded in yellow to red represent agricultural catchments, dominated by either pasture (P) or cropland (C). Grey lines denote elevation contours. The background imagery is derived from Sentinel-2 satellite data.
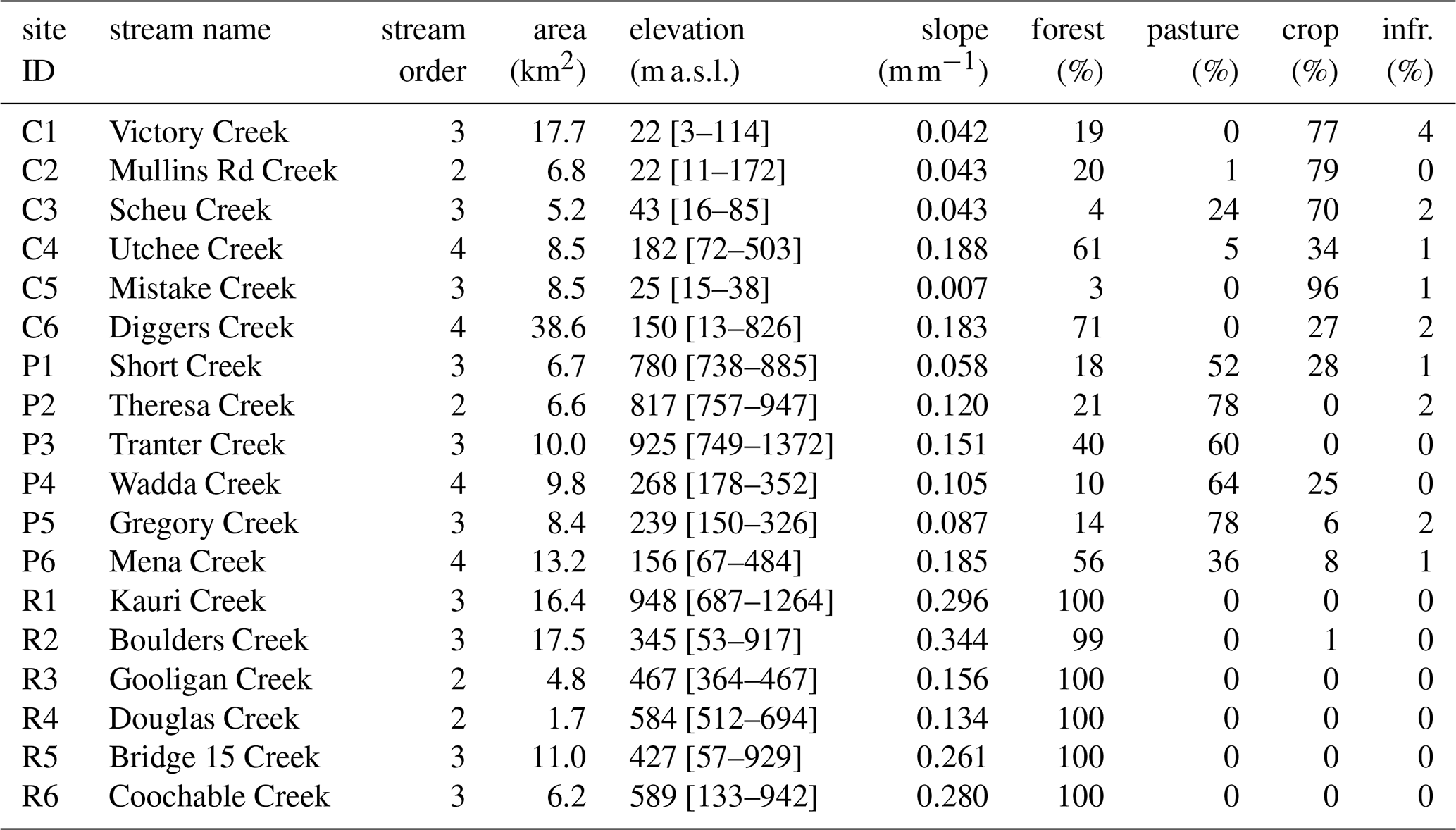
Samples were collected in six streams that drain relatively pristine rainforest remnants (R1 to R6) and 12 streams that drain some degree of agriculturally disturbed land, including six with land converted to pasture (P1 to P6) and six with cropland (mostly sugarcane and banana plantations; C1 to C6) (Fig. 1). Some of the agricultural sites drain a mixture of different land uses (i.e., forested area, agricultural area (pasture/crops) and urban area; Table 1). However, urban land use remains below 4 % at all sites. The six rainforest sites are generally located in more rugged terrain, resulting in higher mean catchment slopes (0.25 m m−1) compared to the 12 disturbed sites (0.11 m m−1) (p=0.005; Mann-Whitney U-test). Streams range from second to fourth order, and while the extent of groundwater connection is unknown, it is likely that all streams receive some level of shallow groundwater inputs.
Table 1Landscape features for the 18 stream sites. Elevations are reported as mean [min–max]. Infr.: infrastructure/urban. Site coordinates are available in Table S4.
2.2 Sample collection
Stream water samples were collected once during the dry season from 25 to 28 August 2020, and a second time at the end of the wet season from 21 to 26 March 2021 (Fig. S1). At each site, electrical conductivity and temperature were measured using a portable conductivity meter (WTW), while the partial pressure of CO2 (pCO2) was measured only during the wet season campaign using a pre-calibrated sensor (eosGP, Eosense) that was allowed to equilibrate for at least 30 min prior to measurement. At each site, samples were taken for later analysis of DOC concentration, δ13C-DOC, 14C-DOC, dissolved organic matter (DOM) composition, and water stable isotopes (deuterium, δ2H and oxygen-18, δ18O). In addition, at three sites (i.e. R1, P1 and C4; randomly selected from each land use type), samples were collected for 14C-DIC analyses. Two samples (P1 and C3 in the wet season) were damaged during analysis and therefore could not be analysed for 14C-DOC.
We collected samples in 1 L high density polyethylene (HDPE) bottles (14C-DIC), 2 L HDPE bottles (14C-DOC), 125 mL HDPE bottles (DOM composition), 40 mL pre-acidified borosilicate amber glass vials (DOC and δ13C-DOC), and 60 mL polyethylene centrifuge vials (δ2H and δ18O). All HDPE bottles were acid-leached with hydrochloric acid and Milli-Q rinsed prior to sampling. Samples for 14C-DIC, 14C-DOC, and DOM composition analyses were filtered in the field using a peristaltic pump (Geopump, GEOTECH) connected to Tygon tubing and high-capacity 0.45 µm polyethersulfone filters (FHT-45, Waterra). The Tygon tubing was pre-rinsed in 10 % hydrochloric acid and Milli-Q water between samples, and both the tubing and the high-capacity filters were then pre-conditioned with the water to be sampled for at least 30 s before sampling.
We also extracted two shallow soil cores in two rainforest sites (R3 and R6) using a hand auger. We sampled soil organic C (SOC) at three depths along these cores (0–5, 10–15, 20–30 cm) for later analysis of 14C-SOC. For each depth, approximately 100 g of soil were collected, double Ziploc bagged, and frozen upon return to the laboratory.
2.3 Laboratory analyses
Both 14C-DOC and 14C-DIC analyses were conducted at the Australian Nuclear Science and Technology Organisation (ANSTO) in Sydney (NSW, Australia). 14C-DOC samples were acidified to pH 2, sparged with N2 gas and neutralised to just below pH 7, rotary evaporated to a concentrate, and freeze-dried to a powder. 14C-DIC samples were acidified on a water line under vacuum, with CO2 released using phosphoric acid. The liberated CO2 was then captured in a vial using liquid nitrogen. Samples for 14C-DOC and 14C-DIC were then combusted using CuO, Ag and Cu wire. CO2 was cryogenically purified before being converted to graphite, as outlined in Hua et al. (2001). The graphite was analysed by accelerator mass spectrometry (Wilcken et al., 2015). We report all 14C data as normalised percent Modern Carbon (pMC), calculated according to Stuiver and Polach (1977). Mean standard errors were ±0.41 pMC for 14C-DOC and ±0.36 pMC for 14C-DIC.
All soil samples were sent to the Radiocarbon Laboratory at the Australian National University (Canberra, ACT, Australia) for 14C-SOC analysis. Prior to analysis, rootlet fragments were sieved out and samples were pretreated with hydrochloric acid to remove any carbonates. The analyses were conducted on a single stage accelerator mass spectrometer following the method described in Fallon et al. (2010). Results are reported as pMC, calculated according to Stuiver and Polach (1977). The mean standard error for 14C-SOC based on all six measurements was ±0.28 pMC.
DOM composition was analysed on an Aqualog optical spectrometer (HORIBA) at ANSTO, with excitation wavelengths of 240–600 nm and emission wavelengths of 250–800 nm. Data were blank corrected, interpolated, corrected for inner filter effects and for Raman scattering and first and second order Rayleigh scattering, and normalised using the Aqualog software (Hansen et al., 2018). We derived peak intensities following Coble (1996), and calculated the humification index (HIX) as per Ohno (2002) and the biological index (BIX) as per Huguet et al. (2009) and Fellman et al. (2010). Peak intensities, HIX and BIX were determined using the R package staRdom (Pucher et al., 2019).
DOC concentrations and δ13C-DOC were measured at the Environmental Analysis Laboratory in Southern Cross University (Lismore, NSW, Australia) using a Flash elemental analyser (Thermo Fisher) connected to a Delta V Plus isotope ratio mass spectrometer (Thermo Fisher), as per Carvalho (2023). δ2H and δ18O were analysed at Charles Darwin University (Darwin, NT, Australia) on a L2130-i cavity ring-down spectrometer (Picarro) connected to a diffusion sampler as described in Munksgaard et al. (2011).
2.4 Catchment delineation and land use classification
We conducted all spatial analyses in QGIS 3.4 (QGIS Development Team, 2019). Catchment boundaries and average catchment slopes were estimated using LiDAR-based elevation data at 5 m resolution (Geoscience Australia, 2015). In areas where the LiDAR data were incomplete or unavailable, we used elevation information from the SRTM 30 (Shuttle Radar Topography Mission, NASA). Land use proportions were derived from Sentinel-2 satellite imagery captured in October 2020. The classification between rainforest, pasture, cropland, and infrastructure was conducted using the Semi-Automatic Classification Plugin (Congedo, 2021) in QGIS 3.4.
2.5 Data analysis
There were very few differences between agricultural sites dominated by pasture and those dominated by cropland, with the only significant differences observed in elevation and stable isotopic values during the dry season (Table S1). Therefore, we grouped these two land uses into a single “agricultural” category. To assess seasonal differences within the same category (rainforest or agricultural), we used the Wilcoxon signed-rank test (signrank function in MATLAB). To compare the two land use categories (rainforest and agricultural) within the same season (wet or dry), we used the Mann-Whitney U-test (ranksum function in MATLAB). Given the small sample sizes, p values lower than 0.1 were viewed as being indicative of differences.
To explore the variability in 14C-DOC and identify potential drivers of these variations, we ran generalised additive models (GAMs) using the fitrgam function in MATLAB, as most relationships between 14C-DOC and predictors were nonlinear (Table S2). The fitrgam function uses cubic B-spline functions by default for continuous predictors. We developed separate GAMs for the dry and wet seasons as we expected the drivers to differ between our two sampling campaigns. All predictor variables were standardised (centred to the mean and scaled to the standard deviation) prior to analysis, and predictors that were strongly collinear were excluded. To assess collinearity, we calculated Pearson correlation coefficients and removed variables with coefficients greater than 0.5 (Fig. S2). This led us to remove δ18O and δ2H (strongly correlated with each other and with temperature), elevation (strongly anticorrelated with temperature), δ13C-DOC (strongly anticorrelated with DOC), HIX (strongly correlated with DOC and anticorrelated with BIX), conductivity (strongly anticorrelated with slope), and the fraction of forest (strongly correlated with slope). Because slope significantly differed between the two land use categories (p=0.005; Mann-Whitney U-test), we added an interaction term (“land use ⋅ slope”) to account for the non-independence between these two predictors. We assessed model performance by comparing Akaike Information Criterion (AIC) values, testing models with and without this interaction term to determine the best fit. The resulting GAM structure was as follows:
where s() indicates a smooth term and ε is the error term. We then used partial dependence plots to visualise the marginal effect of each predictor on 14C-DOC based on the fitted GAMs.
For the three sites where we collected 14C-DIC data, we used a two-endmember mixing approach to estimate the relative contributions of soil CO2 inputs and DOC mineralisation to stream DIC. The expected absence of any significant geogenic DIC source in the study catchments (Jell, 2013) allowed us to undertake this analysis. To represent the soil CO2 endmember (Csoil), we used the mean ± standard deviation of 14C-SOC from the four shallowest soil samples (0–15 cm), giving 104.3±4.3 pMC. To characterise the DOC endmember (CDOC), we used the 14C-DOC value measured at the site on the same day. The fraction of DOC mineralisation (fDOC) to stream DIC (CDIC) was then estimated as follows:
Uncertainty in fDOC was estimated for each site and season by propagating the uncertainty in Csoil, using its upper and lower bounds to generate corresponding bounds in fDOC. This simple approach provided only first-order estimates of the sources of stream DIC, because 14C-DOC likely represents an older, less labile fraction of DOC, which could result in underestimations of fDOC. However, the low HIX values (0.4–4.1) measured across all sites (see Results) indicate a predominance of highly biolabile DOM, suggesting that our mixing model provided coarse yet reliable results.
The two soil cores yielded contrasting results (Table S3), with modern SOC at all three depths at site R6 (all values >104.6 pMC), whereas site R3 had modern (102.8 pMC) SOC only at the shallowest (0–5 cm) depth, and increasingly old SOC at greater depths (99.1 pMC at 10–15 cm and 83.4 pMC at 20–30 cm). All values above 100 pMC indicate samples affected by the nuclear bomb pulse (e.g. Graven et al., 2020).
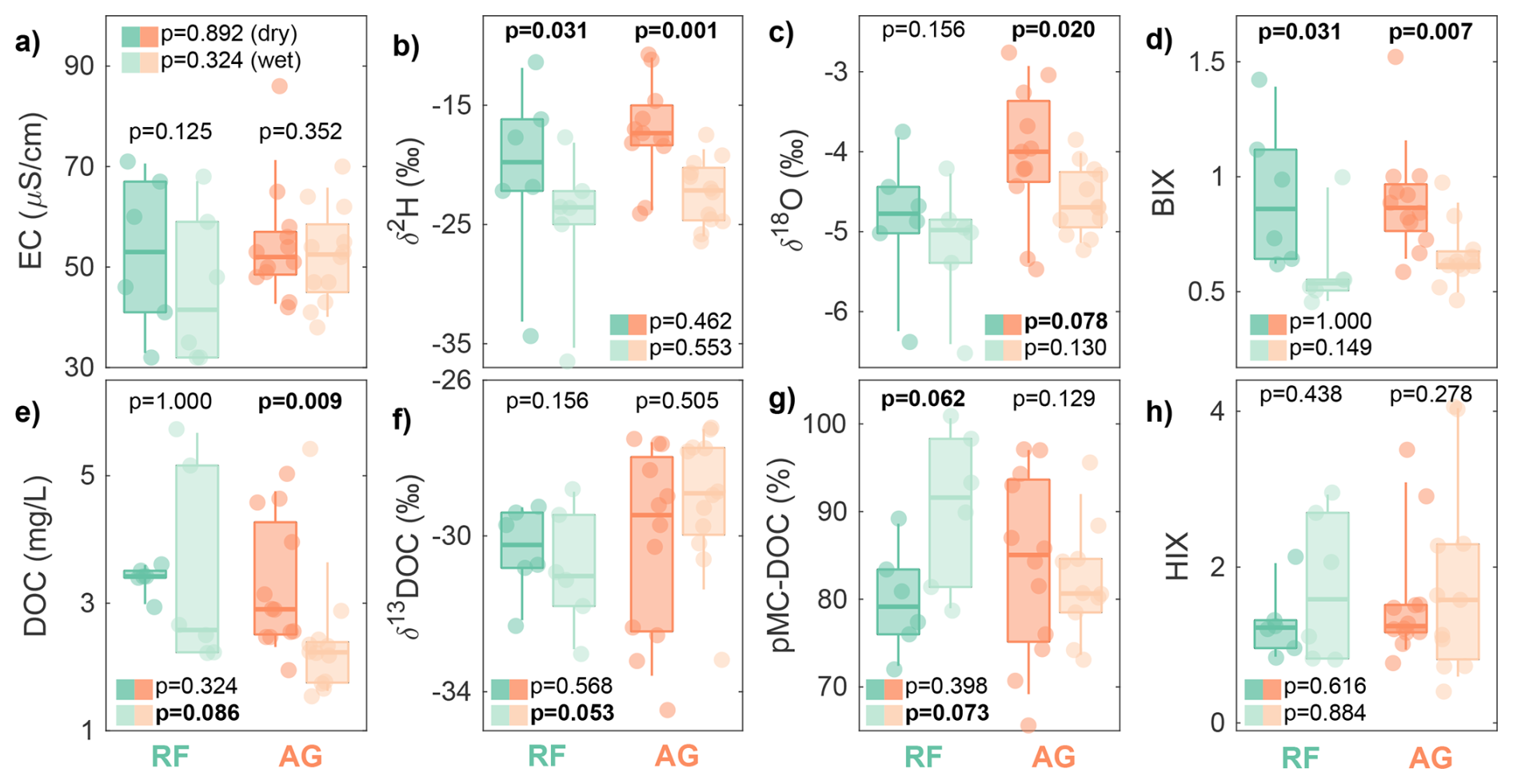
Figure 2Distribution of variables between the dry (dark) and wet (light) seasons across rainforest (RF; green) and agricultural (AG; orange) sites. Variables are (a) electrical conductivity (EC), (b, c) stable isotopes of hydrogen and oxygen in water (δ2H and δ18O), (d) biological index (BIX), (e) DOC, (f) δ13C-DOC, (g) 14C-DOC in percent Modern C (pMC), and (h) humification index (HIX). The p values at the top of each boxplot pair represent the seasonal differences within the same land use category based on Wilcoxon signed-rank tests. The p values next to the coloured boxes represent the differences between land use categories for the dry (dark) and wet (light) seasons based on Mann-Whitney U-tests. Given the small sample sizes, p values lower than 0.1 were considered indicative of a significant difference and are shown in bold.
We also observed large variations in measured stream water parameters across land use categories and seasons (Fig. 2). Differences between the two land uses were significant for more parameters in the wet season (DOC, δ13C-DOC, 14C-DOC) than in the dry season (δ18O). In addition, seasonal differences were significant for more parameters at agricultural sites (δ2H, δ18O, BIX, DOC) than at rainforest sites (δ2H, BIX, 14C-DOC). Water stable isotopes tended to decrease from dry (median −17.7 ‰ for δ2H and −4.2 ‰ for δ18O) to wet seasons (median −22.9 ‰ for δ2H and −4.8 ‰ for δ18O), reflecting a change in rainfall sources and reduced residence times in soils in the wet season, with less soil water subject to evaporation. This seasonal decrease in δ2H and δ18O did not coincide with a decrease in EC (median from 52 to 50 µS cm−1), likely due to the already very low EC levels in the dry season in these small headwater catchments, indicative of fast, shallow flow paths and low weathering rates all year round. DOC concentrations decreased from dry (median 2.9 mg L−1) to wet seasons (median 2.2 mg L−1) in the agricultural sites (p=0.009), whereas there was high cross-site variability and no significant seasonal change in the rainforest sites. Importantly, DOC concentrations varied in response to local flow conditions during the wet season. The highest three concentrations (>5 mg L−1) observed at sites C2, R5, and R6 coincided with the rising limb of a small high-flow event on 25 March (Fig. S1), whereas all other sites were sampled under flow recession, resulting in depleted DOC sources and lower concentrations.
The δ13C-DOC values were always lower than −27 ‰ and relatively stable between seasons and across land use categories, although there was a significant difference (p=0.053) between rainforest sites (median −31.0 ‰) and agricultural sites (median −28.9 ‰) in the wet season. BIX values significantly decreased from dry (median 0.86) to wet seasons (median 0.61), while HIX values were generally low (<4.1) and did not vary consistently with season. Neither BIX nor HIX values were significantly different between land use categories.
We found substantial variability in the age of exported DOC among streams draining the same land use category, resulting in no significant differences between the two land use categories in the dry season (p=0.398) and when both seasons were combined (p=0.589), but a significant difference during the wet season (p=0.073). In addition, the response of individual sites to high flow conditions diverged between the two land use categories (Fig. 3a). In rainforest catchments, high flows mobilised significantly younger DOC compared to baseflow conditions (p=0.062); this pattern was observed at five out of six sites (Fig. 3a). In contrast, in agricultural catchments, high flows tended to mobilise DOC of similar age or older compared to dry season flows, although the difference between seasons was not significant (p=0.129), likely due to the high variability during the dry season. This pattern of similar or increasing DOC age in the wet season was observed at eight of the ten agricultural sites with repeated measurements (Fig. 3a). The fraction of rainforest cover explained 29 % of the variability in these seasonal changes, with sites with more forest cover having a more positive change in DOC age between seasons, i.e. younger DOC in the wet season (Fig. 3b).
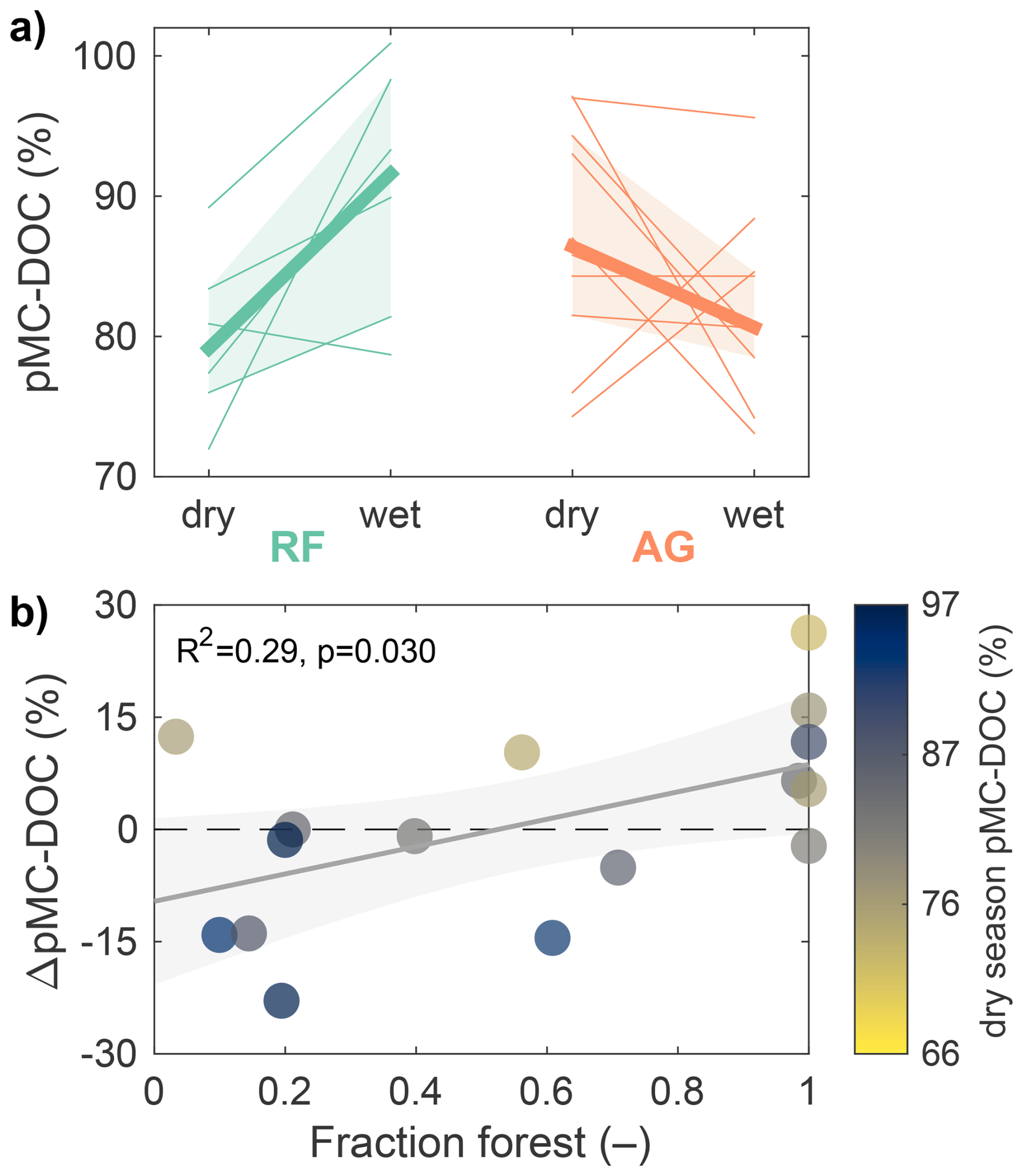
Figure 3Seasonal changes in percent modern C (pMC) in dissolved organic carbon (DOC). (a) Changes in pMC-DOC for the six rainforest (RF) sites and ten agricultural (AG) sites. The thick lines and shaded areas represent the median and interquartile range across all sites. (b) Relationship between seasonal change in pMC from dry to wet seasons (ΔpMC-DOC; positive = younger DOC in the wet season) and fraction of forest cover. All six rainforest sites plot to the right around 1. In the upper plot, the seasonal change in rainforest sites (p=0.062) and the difference between the two land use categories during the wet season (p=0.073; Fig. 2) are significant at a 90 % confidence level, whereas the other temporal and cross-category changes are not.
Examining the linear relationships between DOC age and environmental variables (Table S2), few predictors showed strong correlations with DOC age. In the wet season, both slope and DOC concentration were positively correlated with DOC age (Table S2). The slope–DOC age relationship was particularly strong for rainforest sites in the wet season (R2=0.53; Fig. S3). In contrast, slope had no significant linear relationship with DOC age at agricultural sites in either season (Fig. S3).
The GAM approach provided additional insight that was not detectable using simple linear relationships. We found that the drivers of variation and relationships with DOC age differed strongly between seasons (Fig. 4). In the dry season, catchment area, water temperature, DOC concentration and BIX were positively correlated with pMC-DOC, indicating that higher values of these variables were associated with younger DOC, while land use ⋅ slope was negatively correlated with pMC-DOC. In the wet season, the most significant drivers were land use ⋅ slope and DOC concentration, all positively correlated with pMC-DOC, while catchment area was negatively correlated with pMC-DOC. While the two land use categories explained significant differences in DOC age in the wet season only (Fig. 2), the GAM results indicate that the interaction between slope and land use was a strong driver of pMC-DOC in both seasons. Models that treated land use and slope as separate predictors resulted in higher AIC values (−392 and −341 for the dry and wet seasons, respectively) compared to GAMs that included the land use ⋅ slope interaction (AICs of −395 and −345 for the dry and wet seasons, respectively), suggesting that the interaction term improved model performance.
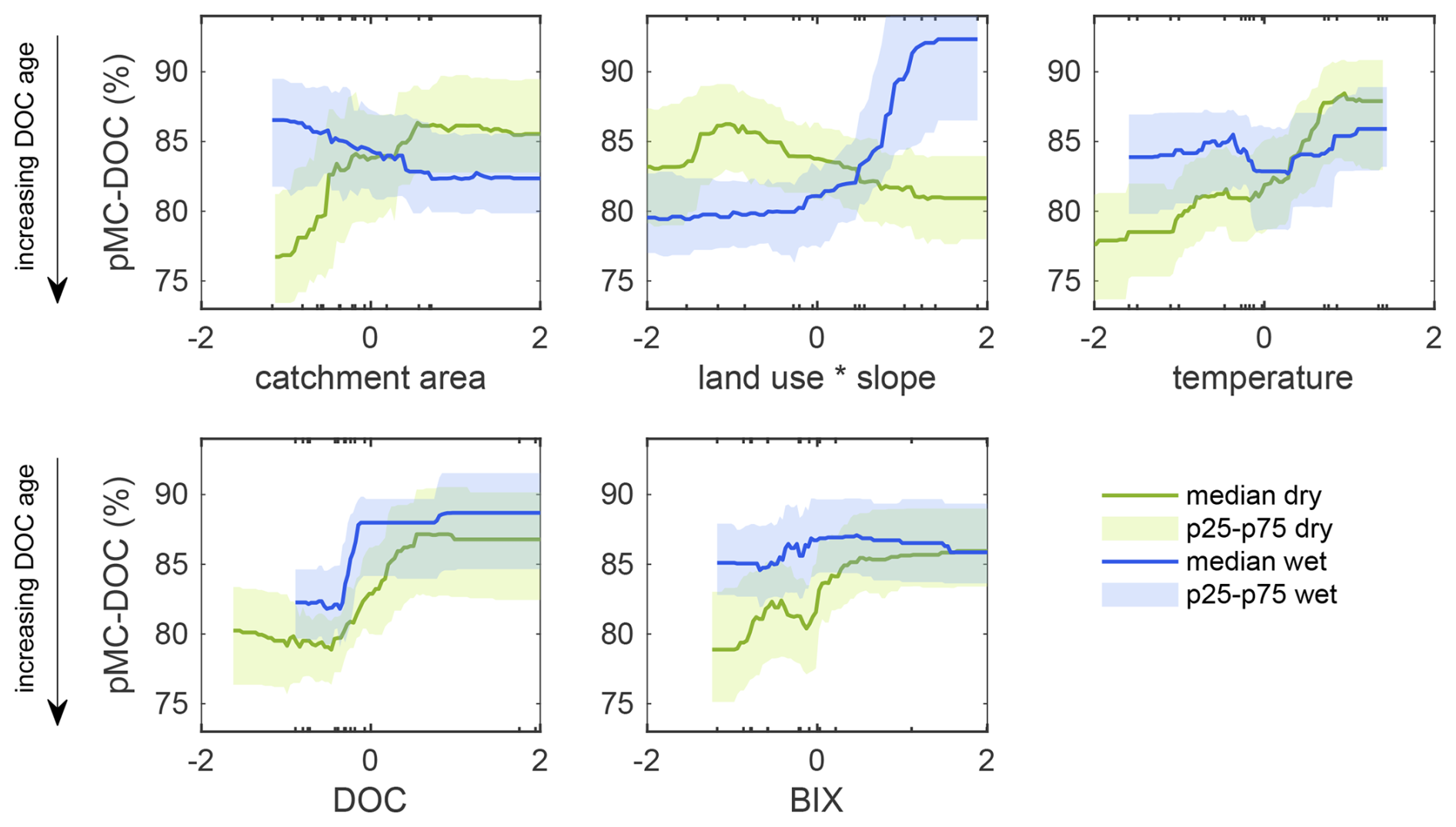
Figure 4Partial dependence plots of the marginal effects of each predictor on 14C-DOC based on the best GAMs developed for the dry and wet seasons. All predictors were standardised prior to analysis, so that negative values on the x axes indicate below-mean values, while positive values indicate above-mean values. The small dashes on the horizontal axes indicate the distribution of predictor values and are duplicated on both axes. Note: higher pMC-DOC corresponds to lower DOC age.
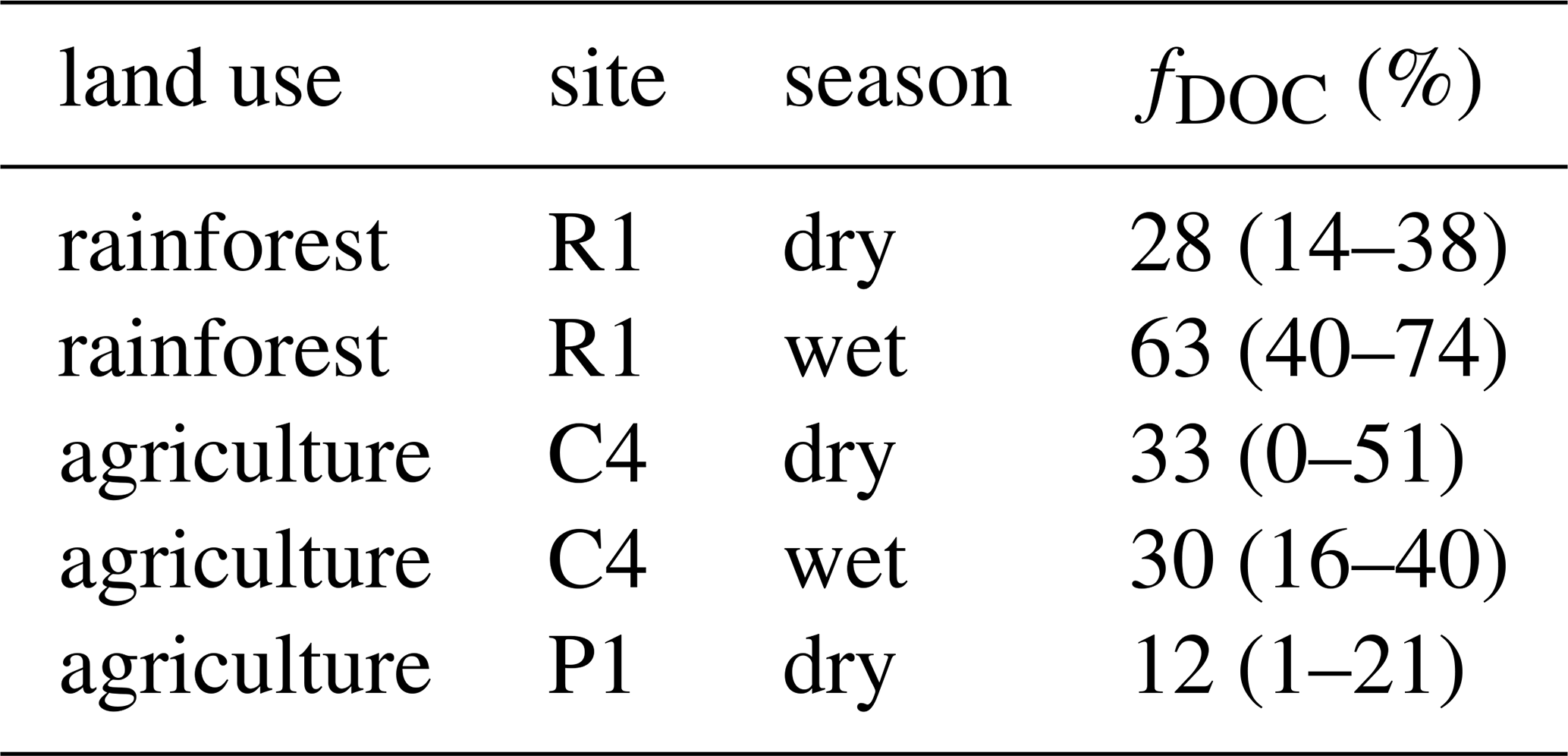
The DIC samples collected at three sites were modern or close to modern, with mean values of 97.1 pMC (R1), 98.5 pMC (C4) and 100.4 pMC (P1). Values slightly increased (i.e. DIC became younger) in the wet season at R1 and P1, whereas they decreased at C4. DIC was always younger than DOC, and at each site the DIC and DOC ages shifted in the same direction between seasons, although the direction of change differed between sites (Fig. 5). At the rainforest site (R1), both DIC and DOC were younger in the wet season, whereas at the agricultural site (C4), both were older in the wet season. Unfortunately, the wet season 14C-DOC sample from the third site (P1) was damaged during the analysis, so we have no DOC age data to compare the wet season 14C-DIC sample to. Using our simple endmember analysis, we estimated that between 12 % (1–21) and 63 % (40–74) (mean across sites and seasons: 33 %) of DIC might have originated from in-stream mineralisation of DOC, with the rest likely originating from soil CO2 (Table 2).

Figure 5Concomitant changes in percent modern C (pMC) in DIC and DOC for streams representative of the two broad land use categories (R1: rainforest; P1 and C4: agricultural). The dashed line represents the 1:1 line, along which DIC and DOC would have equivalent ages.
Table 2Mean estimates of the fraction of DOC mineralised to DIC (fDOC, %) in one rainforest stream (R1) and two agricultural streams (C4 and P1) during the dry and wet seasons. Uncertainty ranges are given in parentheses.
4.1 Controls on DOC age in rainforest versus agricultural streams
We observed high variability in DOC age across sites, with no consistent differences between rainforest and agricultural sites; a result that allows us to reject our first hypothesis that older DOC is exported after land conversion to agriculture. A range of factors may explain this cross-site variability. First, our two soil cores had very distinct age profiles (Table S3), with SOC at >20 cm depth being either modern or centuries old. Such discrepancy may be linked to geomorphic controls: the R6 core, which contained modern SOC at depth, was taken from a low point near the stream, where organic matter likely accumulates more rapidly. In contrast, the R3 core, which contained old SOC at depth, was taken from a gentle slope. Such spatial variability in SOC age is not uncommon in Australian landscapes (Hobley et al., 2017; Bowman et al., 2004) and might have contributed to some of the differences in DOC age observed across sites. Shallow (0–30 cm) soil C in tropical forests has been reported in the range 95–104 pMC (Shi et al., 2020), i.e. from modern to ∼400 years BP (5th–95th percentiles). This range broadly encompasses the integrated age of our two soil profiles, with R3 at the older end and R6 at the younger end of the range. Given its location on a gentle slope, we believe the R3 core may be more representative of broader landscape conditions where DOC is produced and exported.
Hydrological flow paths also appeared to play an important role. The negative effect of DOC concentration on DOC age (years BP) in both seasons (Fig. 4) (despite this trend being weak in the raw data during the dry season; Table S2) suggests that some streams may be fed by slightly deeper subsurface contributions, with older and more processed, lower concentration DOC, whereas others were fed by shallower flow paths, which tend to contain younger and more concentrated DOC (Barnes et al., 2018; Campeau et al., 2019; Sanderman et al., 2009) as natural soil formation processes allow for a decreasing age with depth. Various factors can influence flow path depth, including seasonal changes in flow regime and catchment morphology. Steep catchments tend to have shallower flow paths and shorter residence times (Remondi et al., 2019; Tetzlaff et al., 2009), leading to the export of younger DOC. Steep forested catchments are also likely to generate high runoff containing freshly leached organic material from both throughfall (Van Stan and Stubbins, 2018; Behnke et al., 2023) and leaf litter (Janeau et al., 2014; Miranda and Avelar, 2022). In our study, slope was an important predictor of DOC age, particularly in the wet season and for rainforest sites, where steeper catchments tended to export younger DOC. A similar influence of catchment morphology on DOC age has been observed in subtropical mountainous catchments (Chen et al., 2023). Interestingly, DOC ages (years BP) in the agricultural catchments had no significant relationship with slope in either season, contrasting with the negative relationship observed in rainforest catchments in the wet season (Fig. S3). This finding suggests that in agricultural catchments, the rate and magnitude of soil erosion, rather than flow path depth, may be a key control on DOC ages. In these systems, it is possible that steeper catchments might even increase the mobilisation of aged soil C, likely exposed by tillage and/or cattle trampling (Sickman et al., 2010; Drake et al., 2019).
The sources of DOC were relatively similar between rainforest and agricultural catchments, with δ13C-DOC compositions indicating similarly depleted, likely C3 plant-derived biomass as the dominant source. This is despite the agricultural catchments having available C4 pools (sugar cane and some pasture grass species), and suggests that most DOC in disturbed catchments originated from older, pre-clearing soil C pools – a result similar to that of Moyer et al. (2013). Lower contributions of C4-derived C sources than would be expected based on land use have been related to potentially large contributions from riparian forests and to differences in the biolability of different DOC sources (Bouillon et al., 2012, 2007). While we lack δ13C-SOC data to confirm the importance of older soil C sources to the stream DOC pool in disturbed catchments, the generally older wet-season DOC ages at these sites tend to support our conclusion (see following section). The relatively low HIX values across sites and seasons indicate that regardless of DOC age, highly biolabile compounds, which are typically low molecular weight and less aromatic in nature (Berggren et al., 2009; Li et al., 2023), may be dominating the DOC pool. This is not unexpected, as previous studies have shown that aged C can also be relatively enriched in energy-rich organic matter at the molecular level and be highly biolabile and readily oxidised by microbes (Mann et al., 2015; Drake et al., 2019; Spencer et al., 2015).
Despite differences across sites and no clear distinction between DOC ages in rainforest versus agricultural catchments, our data revealed some unexpected findings. In contrast to most previous tropical studies, we found that DOC in small, headwater streams of humid tropical regions can be centuries to millennia old (with a median of 82.5 pMC or 1553 years BP across sites and seasons), even in pristine rainforest areas. Recent studies have reported similar occurrences of old DOC at low flow in small, relatively undisturbed catchments in temperate regions (Tittel et al., 2022; Rhyner et al., 2023). Our findings suggest that this pattern may also apply to tropical regions, challenging the view of faster DOC turnover in the tropics (Spencer et al., 2012; Mayorga et al., 2005; Müller et al., 2015; Marwick et al., 2015). One possible explanation for this result is the higher elevations (hence lower temperatures) of our six pristine sites (mean elevations 345–948 m a.s.l.; Cfa climate class), which may have led to relatively slower decomposition rates and older SOC accumulating over time, compared to what would be expected in warmer, lowland tropical areas. Another potential factor is the contribution of old groundwater-derived DOC to streams (McDonough et al., 2020), a possibility we discuss further in Sect. 4.3.
4.2 Seasonal shifts in DOC age and pathways
Our results show that DOC age varied significantly between dry and wet seasons, with marked differences between rainforest catchments, where DOC tended to be younger at high flow, and agricultural catchments, where DOC was similar or older at high flow (Fig. 6). These findings support our second hypothesis that DOC is younger at high flow for rainforest sites, but contradict it for agricultural sites. We interpret these differences between land use categories as reflecting fundamental differences in system dynamics. Rainforest sites were in an “equilibrium” state, where DOC age was a direct function of flow path depth and reflected the hydrological shift between dry and wet seasons. This pattern is well documented in temperate and northern high-latitude catchments, where during high flow conditions the shallower riparian or organic-rich surface soil and litter layers become connected to streams and mobilise younger DOC (e.g. Tittel et al., 2015; Barnes et al., 2018; Evans et al., 2022; Campeau et al., 2019; Aiken et al., 2014). Additionally, the younger DOC observed in the wet season may also be attributed to increased leaching of fresh leaf litter (abundant in the rainforest catchments) through surface flow pathways, as reported in similar steep tropical rainforest regions (Miranda and Avelar, 2022). Leaf litter is a source of young, unprocessed, low molecular weight, biolabile C (e.g. Hudson et al., 2018; Harfmann et al., 2019), and the low HIX and BIX values observed at our rainforest sites during the wet season are a strong indication that leaf litter leaching contributed to DOC in these streams.
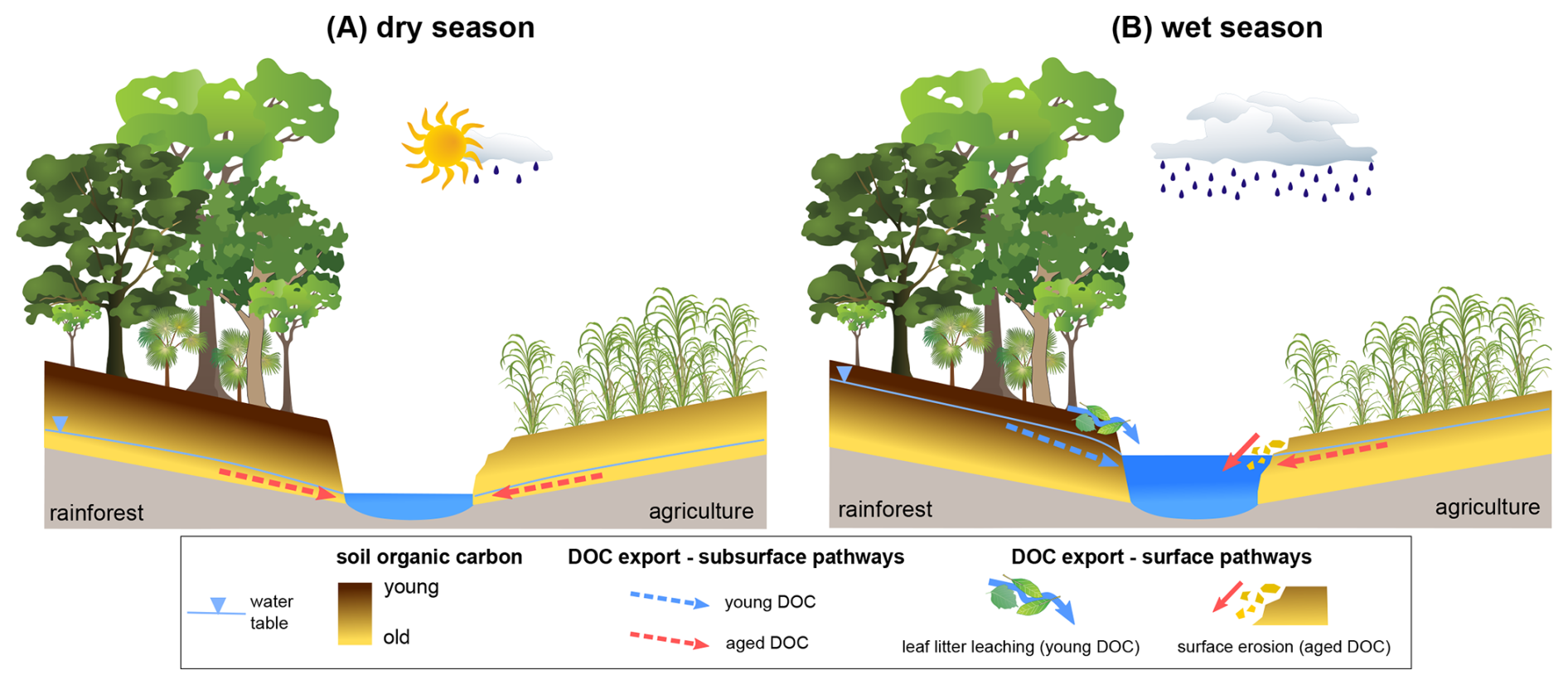
Figure 6Conceptual diagram showing the shift in DOC sources between the (A) dry and (B) wet seasons in rainforest versus agricultural catchments.
In contrast, agricultural sites operated under unsteady-state conditions, where old DOC was exported during both wet and dry periods, although flow conditions still influenced DOC age. This likely results from anthropogenic disturbance removing the bulk of modern C from organic-rich surface soil and litter layers (Drake et al., 2019; Moore et al., 2013; Lee et al., 2021). At some agricultural sites, higher flow conditions even led to the export of older DOC, a result that aligns with Moore et al. (2013) who observed the export of older DOC during the wet season from two disturbed peat catchments in Borneo. We propose that at these sites, processes such as tillage, cattle trampling and bank erosion may have exposed deeper, older C at the surface, increasing the leaching and transport of older DOC via surface rather than subsurface pathways during the wet season (Fig. 6). Although we do not present direct evidence linking older soil C to disturbance, all our agricultural sites were formerly rainforest (Kemp et al., 2007; Vanclay, 1996) and there is a long history of erosion in the Great Barrier Reef region (e.g. Kroon et al., 2016; McKergow et al., 2005), suggesting that substantial loss of young SOC due to anthropogenic disturbance is likely. While the role of surface erosion on DOC export is complex and largely unclear, studies have reported enhanced DOC mobilisation with increased erosion (Van Gaelen et al., 2014; Chen et al., 2023). Alternatively, older DOC at higher flow could also originate from 14C-dead organic chemicals transported via runoff (Sickman et al., 2010), such as synthetic fertilisers commonly used in sugarcane cultivation – however, we suggest this explanation is likely not a major control on DOC age across our sites, as it would not apply to pasture sites where fertilisers are not used.
In the wet season, the two land use categories had significantly different DOC sources (p=0.053), with more depleted values (median ‰) in rainforest sites and slightly more enriched values (median ‰) in agricultural sites. These differences may be related to increased contributions of young, C3 plant-derived organic C in the rainforest sites compared to older SOC in agricultural sites, as the latter is typically more 13C enriched than C3 plant-derived biomass (Moyer et al., 2013; Drake et al., 2019). These apparent differences in DOC sources were not reflected in the BIX and HIX indices, where seasonal variations were more pronounced than the differences between land use categories (Fig. 2). The decrease in BIX values from dry to wet seasons suggests a shift from a mixture of aquatic/microbial and terrestrial DOM sources to mostly terrestrial sources (Huguet et al., 2009; Fellman et al., 2010). This shift is consistent with increasing hydrological connectivity between soil C pools and streams, and in some cases, with higher surface erosion and runoff. The low HIX values across all sites indicate low humification, which is relatively common in tropical streams and rivers (Ji et al., 2024; Iles et al., 2022; Lambert et al., 2016). This suggests that even older SOC pools underwent limited humification before being mobilised into streams as DOC. Additionally, at sites with gentler slopes, slower streamflow and greater evaporation, older C might have been photodegraded over time, transforming aromatic-rich organic matter to lower molecular weight, more biolabile moeities (Stubbins et al., 2010). Alternatively, the dominance of low molecular weight DOM could suggest potential connection to regional groundwater systems at certain sites, as groundwater has been shown to contain old C with low molecular weight (McDonough et al., 2020). This explanation is explored further in the following section.
4.3 Decoupling between DIC and DOC cycling
Unlike DOC, DIC was mostly modern (96.5 < pMC-DIC < 101.2) and always younger than DOC, consistent with our third hypothesis that DIC is younger than DOC in the study region. At the three sampled sites (R1, P1 and C4), DIC was likely to be primarily sourced from young, soil-derived CO2 inputs to streams (37 %–88 %) rather than from internal respiration of older DOC (12 %–63 %). While these estimates should be interpreted cautiously given the limited number of DIC measurements, our results suggest a decoupling between DIC and DOC cycling, regardless of land use. Consistent with our work, previous studies have also observed a disconnect between the sources and controls of DIC and DOC in streams, as reported in both boreal and tropical areas (Campeau et al., 2019; Moyer et al., 2013). Younger CO2/DIC has been associated with the lateral export of fresh soil CO2 (Campeau et al., 2019) or to direct inputs of atmospheric CO2 (Moyer et al., 2013). The dominance of soil-derived CO2 inputs to the stream DIC pool is well-documented in Australian tropical streams (Solano et al., 2024; Duvert et al., 2020) and, more generally, in low-order streams (Hotchkiss et al., 2015; Battin et al., 2023). However, in-stream DOC respiration can also be a substantial source of stream DIC (e.g. Solano et al., 2023), and it is interesting to note that even in these relatively steep headwater streams characterised by short residence times, stream DOC mineralisation contributed a non-negligible fraction of the stream DIC pool. This may have been facilitated by potentially high DOM biolability across sites, as indicated by the low HIX values, and relatively high BIX values, particularly in the dry season.
Groundwater is increasingly recognised as a potentially significant source of old, biolabile DOC to surface systems (McDonough et al., 2022, 2020). There is a possibility that at our sites, some old DOC could have entered streams via groundwater rather than solely through the mobilisation of old soil organic C. However, the DIC data suggest this is unlikely, as DIC remained modern or near-modern in both the wet and dry seasons. Any groundwater contributions to streams likely originated from shallow flow paths composed of recently recharged waters, as further corroborated by the low EC values across all sites and seasons and the significant seasonal changes in water stable isotopes, which suggest minimal regional groundwater influence and short residence times in the subsurface. Our DIC dataset includes only three of the 18 sites, so additional DIC age data would be necessary to definitively rule out groundwater as a source of old DOC. This would be particularly relevant in larger, flatter catchments where older, regional groundwater contributions may be more prevalent.
Our study shows that DOC exported from tropical headwater streams can be old, particularly during drier periods, challenging the common view of fast SOC turnover in tropical systems. We also highlight the influence of seasonal variations on DOC age, with flow path depth and leaf litter layer leaching through wet-season runoff being the dominant factors in pristine rainforest catchments, with DOC becoming younger at high flow. In contrast, in agricultural catchments, DOC age was primarily influenced by the loss of modern C due to land conversion and contemporary soil erosion, particularly at high flow. Lastly, our findings suggest that the stream DIC and DOC pools are largely decoupled in these systems, with DIC being younger than DOC and in-stream respiration of DOC representing a minor fraction of the DIC pool.
While our findings provide valuable new insights, there is still much to explore. In particular, our wet season sampling occurred three weeks after the last major rainfall event (Fig. S1) and during flow recession conditions, when soil DOC sources may have already been depleted. Clearly, more frequent sampling will be required to provide a clearer picture of the temporal dynamics of DOC age at different stages of the wet season. Similarly, while our GAM analysis provided useful insights into the drivers of DOC age, it is based on a limited number of observations, so these results should be interpreted cautiously and verified with larger datasets.
Climate change is expected to intensify hydrological extremes, potentially leading to increased erosion and depletion of soil C stocks in agricultural areas. Understanding the extent to which old soil C is reintroduced into the modern C cycle under these changing conditions will be crucial for predicting long-term C fluxes and ecosystem responses, particularly in disturbed tropical landscapes.
The code used in this study is available upon request from the corresponding author.
The dataset used in this study is available at https://www.hydroshare.org/resource/c5e20d5ffe5441e2bf54ba0561fb4dd4/ (Duvert, 2025).
The supplement related to this article is available online at https://doi.org/10.5194/bg-23-1755-2026-supplement.
CD, DEB, DIC and RGMS designed the study. CD conducted fieldwork with assistance from NCM for site selection. VS performed the geospatial analyses. LMD performed the DOM composition analyses. CD analysed and interpreted the data with inputs from LMD, VS and FUC. CD wrote the initial draft which all authors reviewed and edited.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We acknowledge financial support from an AINSE 2019 Early Career Researcher Grant and a CDU Covid Supplementary Funding Project, both awarded to CD. Our thanks go to Peter Freeman, Vladimir Levchenko (ANSTO), Matheus Carvalho (Southern Cross University) and Stewart Fallon (Australian National University) for assistance with laboratory work, to Rainy Comley and Michael Bird (James Cook University) for support with field logistics, and to Paula Martínez for fieldwork assistance. We also thank the two anonymous reviewers and Editor for helping us improve the paper. Clément Duvert is supported by a fellowship from the Australian Research Council (grant no. DE220100852).
This research has been supported by the Australian Institute of Nuclear Science and Engineering (grant no. ECRG 2019) and Charles Darwin University (Covid Supplementary Funding Project 001).
This paper was edited by Gabriel Singer and reviewed by two anonymous referees.
Aiken, G. R., Spencer, R. G. M., Striegl, R. G., Schuster, P. F., and Raymond, P. A.: Influences of glacier melt and permafrost thaw on the age of dissolved organic carbon in the Yukon River basin, Global Biogeochem. Cy., 28, 525–537, https://doi.org/10.1002/2013GB004764, 2014.
Barnes, R. T., Butman, D. E., Wilson, H., and Raymond, P. A.: Riverine export of aged carbon driven by flow path depth and residence time, Environ. Sci. Technol., 52, 1028–1035, https://doi.org/10.1021/acs.est.7b04717, 2018.
Battin, T. J., Lauerwald, R., Bernhardt, E. S., Bertuzzo, E., Gener, L. G., Hall, R. O., Hotchkiss, E. R., Maavara, T., Pavelsky, T. M., Ran, L., Raymond, P., Rosentreter, J. A., and Regnier, P.: River ecosystem metabolism and carbon biogeochemistry in a changing world, Nature, 613, 449–459, https://doi.org/10.1038/s41586-022-05500-8, 2023.
Beck, H. E., McVicar, T. R., Vergopolan, N., Berg, A., Lutsko, N. J., Dufour, A., Zeng, Z., Jiang, X., van Dijk, A. I. J. M., and Miralles, D. G.: High-resolution (1 km) Köppen-Geiger maps for 1901–2099 based on constrained CMIP6 projections, Sci. Data, 10, 724, https://doi.org/10.1038/s41597-023-02549-6, 2023.
Behnke, M. I., Fellman, J. B., D'Amore, D. V., Gomez, S. M., and Spencer, R. G. M.: From canopy to consumer: what makes and modifies terrestrial DOM in a temperate forest, Biogeochemistry, 164, 185–205, https://doi.org/10.1007/s10533-022-00906-y, 2023.
Berggren, M., Laudon, H., Haei, M., Ström, L., and Jansson, M.: Efficient aquatic bacterial metabolism of dissolved low-molecular-weight compounds from terrestrial sources, The ISME Journal, 4, 408–416, https://doi.org/10.1038/ismej.2009.120, 2009.
Billett, M. F., Garnett, M. H., and Harvey, F.: UK peatland streams release old carbon dioxide to the atmosphere and young dissolved organic carbon to rivers, Geophys. Res. Lett., 34, https://doi.org/10.1029/2007GL031797, 2007.
Bouillon, S., Dehairs, F., Schiettecatte, L.-S., and Borges, A. V.: Biogeochemistry of the Tana estuary and delta (northern Kenya), Limnol. Oceanogr., 52, 46–59, https://doi.org/10.4319/lo.2007.52.1.0046, 2007.
Bouillon, S., Yambélé, A., Spencer, R. G. M., Gillikin, D. P., Hernes, P. J., Six, J., Merckx, R., and Borges, A. V.: Organic matter sources, fluxes and greenhouse gas exchange in the Oubangui River (Congo River basin), Biogeosciences, 9, 2045–2062, https://doi.org/10.5194/bg-9-2045-2012, 2012.
Bowman, D. M. J. S., Cook, G. D., and Zoppi, U.: Holocene boundary dynamics of a northern Australian monsoon rainforest patch inferred from isotopic analysis of carbon (14C and δ13C) and nitrogen (δ15N) in soil organic matter, Austral Ecol., 29, 605–612, https://doi.org/10.1111/j.1442-9993.2004.01394.x, 2004.
Bureau of Meteorology: Climate Data Online: http://www.bom.gov.au/climate/data, last access: 21 February 2025.
Butman, D. E., Wilson, H. F., Barnes, R. T., Xenopoulos, M. A., and Raymond, P. A.: Increased mobilization of aged carbon to rivers by human disturbance, Nat. Geosci., 8, 112–116, https://doi.org/10.1038/ngeo2322, 2015.
Campeau, A., Bishop, K., Amvrosiadi, N., Billett, M. F., Garnett, M. H., Laudon, H., Öquist, M. G., and Wallin, M. B.: Current forest carbon fixation fuels stream CO2 emissions, Nat. Commun., 10, 1876, https://doi.org/10.1038/s41467-019-09922-3, 2019.
Carvalho, M. C.: Adapting an elemental analyser to perform high-temperature catalytic oxidation for dissolved organic carbon measurements in water, Rapid Commun. Mass Sp., 37, e9451, https://doi.org/10.1002/rcm.9451, 2023.
Chen, S., Zhong, J., Ran, L., Yi, Y., Wang, W., Yan, Z., Li, S., and Mostofa, K. M. G.: Geomorphologic controls and anthropogenic impacts on dissolved organic carbon from mountainous rivers: insights from optical properties and carbon isotopes, Biogeosciences, 20, 4949–4967, https://doi.org/10.5194/bg-20-4949-2023, 2023.
Chen, Z. L., Yi, Y., Fu, W., Liang, W., Li, P., Wang, K., Zhang, L., Dong, K., Li, S.-L., Xu, S., and He, D.: Severe flood modulates the sources and age of dissolved organic carbon in the Yangtze River Estuary, Environ. Res., 252, 119040, https://doi.org/10.1016/j.envres.2024.119040, 2024.
Coble, A. A., Wymore, A. S., Potter, J. D., and McDowell, W. H.: Land use overrides stream order and season in driving dissolved organic matter dynamics throughout the year in a river network, Environ. Sci. Technol., 56, 2009–2020, https://doi.org/10.1021/acs.est.1c06305, 2022.
Coble, P. G.: Characterization of marine and terrestrial DOM in seawater using excitation-emission matrix spectroscopy, Mar. Chem., 51, 325–346, https://doi.org/10.1016/0304-4203(95)00062-3, 1996.
Congedo, L.: Semi-Automatic Classification Plugin: A Python tool for the download and processing of remote sensing images in QGIS., J. Open Source Softw., 6, 3172, https://doi.org/10.21105/joss.03172, 2021.
Dean, J. F., Coxon, G., Zheng, Y., Bishop, J., Garnett, M. H., Bastviken, D., Galy, V., Spencer, R. G. M., Tank, S. E., Tipper, E. T., Vonk, J. E., Wallin, M. B., Zhang, L., Evans, C. D., and Hilton, R. G.: Old carbon routed from land to the atmosphere by global river systems, Nature, 642, 105–111, https://doi.org/10.1038/s41586-025-09023-w, 2025.
Don, A., Schumacher, J., and Freibauer, A.: Impact of tropical land-use change on soil organic carbon stocks – a meta-analysis, Glob. Change Biol., 17, 1658–1670, https://doi.org/10.1111/j.1365-2486.2010.02336.x, 2011.
Drake, T. W., Van Oost, K., Barthel, M., Bauters, M., Hoyt, A. M., Podgorski, D. C., Six, J., Boeckx, P., Trumbore, S. E., Cizungu Ntaboba, L., and Spencer, R. G. M.: Mobilization of aged and biolabile soil carbon by tropical deforestation, Nat. Geosci., 12, 541–546, https://doi.org/10.1038/s41561-019-0384-9, 2019.
Drake, T. W., Podgorski, D. C., Dinga, B., Chanton, J. P., Six, J., and Spencer, R. G. M.: Land-use controls on carbon biogeochemistry in lowland streams of the Congo Basin, Glob. Change Biol., 26, 1374–1389, https://doi.org/10.1111/gcb.14889, 2020.
Duvert, C.: Dataset: stream carbon age in the Australian humid tropics, HydroShare [data set], http://www.hydroshare.org/resource/c5e20d5ffe5441e2bf54ba0561fb4dd4 (last access: 4 March 2026), 2025.
Duvert, C., Hutley, L. B., Birkel, C., Rudge, M., Munksgaard, N. C., Wynn, J. G., Setterfield, S. A., Cendón, D. I., and Bird, M. I.: Seasonal shift from biogenic to geogenic fluvial carbon caused by changing water sources in the wet-dry tropics, J. Geophys. Res.-Biogeo., 125, e2019JG005384, https://doi.org/10.1029/2019JG005384, 2020.
Evans, C. D., Page, S. E., Jones, T., Moore, S., Gauci, V., Laiho, R., Hruška, J., Allott, T. E. H., Billett, M. F., Tipping, E., Freeman, C., and Garnett, M. H.: Contrasting vulnerability of drained tropical and high-latitude peatlands to fluvial loss of stored carbon, Global Biogeochem. Cy., 28, 1215–1234, https://doi.org/10.1002/2013GB004782, 2014.
Evans, M. G., Alderson, D. M., Evans, C. D., Stimson, A., Allott, T. E. H., Goulsbra, C., Worrall, F., Crouch, T., Walker, J., Garnett, M. H., and Rowson, J.: Carbon loss pathways in degraded peatlands: New insights from radiocarbon measurements of peatland waters, J. Geophys. Res.-Biogeo., 127, e2021JG006344, https://doi.org/10.1029/2021JG006344, 2022.
Fallon, S. J., Fifield, L. K., and Chappell, J. M.: The next chapter in radiocarbon dating at the Australian National University: Status report on the single stage AMS, Nuclear Instruments and Methods in Physics Research Section B: Beam Interactions with Materials and Atoms, 268, 898–901, https://doi.org/10.1016/j.nimb.2009.10.059, 2010.
Fellman, J. B., Hood, E., and Spencer, R. G. M.: Fluorescence spectroscopy opens new windows into dissolved organic matter dynamics in freshwater ecosystems: A review, Limnol. Oceanogr., 55, 2452–2462, https://doi.org/10.4319/lo.2010.55.6.2452, 2010.
Geoscience Australia: Digital Elevation Model (DEM) of Queensland derived from LiDAR 5 Metre Grid, Geoscience Australia [data set], https://doi.org/10.26186/89644, 2015.
Graven, H., Keeling, R. F., and Rogelj, J.: Changes to carbon isotopes in atmospheric CO2 over the industrial era and into the future, Global Biogeochem. Cy., 34, e2019GB006170, https://doi.org/10.1029/2019GB006170, 2020.
Guo, D., Zhang, Q., Minaudo, C., Dupas, R., Duvert, C., Liu, S., Zhang, K., Bende-Michl, U., and Lintern, A.: Australian water quality trends over two decades show deterioration in the Great Barrier Reef region and recovery in the Murray-Darling Basin, Commun. Earth Environ., 6, 67, https://doi.org/10.1038/s43247-025-02044-3, 2025.
Hansen, A. M., Fleck, J., Kraus, T. E. C., Downing, B. D., von Dessonneck, T., and Bergamaschi, B. A.: Procedures for using the Horiba Scientific Aqualog® fluorometer to measure absorbance and fluorescence from dissolved organic matter, Reston, VA, Report 2018-1096, https://doi.org/10.3133/ofr20181096, 2018.
Harfmann, J. L., Guillemette, F., Kaiser, K., Spencer, R. G. M., Chuang, C.-Y., and Hernes, P. J.: Convergence of terrestrial dissolved organic matter composition and the role of microbial buffering in aquatic ecosystems, J. Geophys. Res.-Biogeo., 124, 3125–3142, https://doi.org/10.1029/2018JG004997, 2019.
Hobley, E., Baldock, J., Hua, Q., and Wilson, B.: Land-use contrasts reveal instability of subsoil organic carbon, Glob. Change Biol., 23, 955–965, https://doi.org/10.1111/gcb.13379, 2017.
Hotchkiss, E. R., Hall Jr., R. O., Sponseller, R. A., Butman, D., Klaminder, J., Laudon, H., Rosvall, M., and Karlsson, J.: Sources of and processes controlling CO2 emissions change with the size of streams and rivers, Nat. Geosci., 8, 696–699, https://doi.org/10.1038/ngeo2507, 2015.
Hua, Q., Jacobsen, G. E., Zoppi, U., Lawson, E. M., Williams, A. A., Smith, A. M., and McGann, M. J.: Progress in radiocarbon target preparation at the Antares AMS Centre, Radiocarbon, 43, 275–282, https://doi.org/10.1017/S003382220003811X, 2001.
Hudson, J. E., Levia, D. F., Wheeler, K. I., Winters, C. G., Vaughan, M. C. H., Chace, J. F., and Sleeper, R.: American beech leaf-litter leachate chemistry: Effects of geography and phenophase, J. Plant Nutr. Soil Sc., 181, 287–295, https://doi.org/10.1002/jpln.201700074, 2018.
Huguet, A., Vacher, L., Relexans, S., Saubusse, S., Froidefond, J. M., and Parlanti, E.: Properties of fluorescent dissolved organic matter in the Gironde Estuary, Org. Geochem., 40, 706–719, https://doi.org/10.1016/j.orggeochem.2009.03.002, 2009.
Iles, J. A., Pettit, N. E., Donn, M. J., and Grierson, P. F.: Phosphorus sorption characteristics and interactions with leaf litter-derived dissolved organic matter leachate in iron-rich sediments of a sub-tropical ephemeral stream, Aquat. Sci., 84, 56, https://doi.org/10.1007/s00027-022-00888-x, 2022.
Janeau, J. L., Gillard, L. C., Grellier, S., Jouquet, P., Le, T. P. Q., Luu, T. N. M., Ngo, Q. A., Orange, D., Pham, D. R., Tran, D. T., Tran, S. H., Trinh, A. D., Valentin, C., and Rochelle-Newall, E.: Soil erosion, dissolved organic carbon and nutrient losses under different land use systems in a small catchment in northern Vietnam, Agric. Water Manage., 146, 314–323, https://doi.org/10.1016/j.agwat.2014.09.006, 2014.
Jell, P. A. (Ed.): Geology of Queensland, Geological Survey of Queensland, Brisbane, QLD, Australia, ISBN 978-1-921489-76-1, https://geoscience.data.qld.gov.au/dataset/cr145752 (last access: 4 March 2026), 2013.
Ji, H., Wang, H., Wu, Z., Wang, D., Wang, X., Fu, P., Li, C., and Deng, W.: Source, composition and molecular diversity of dissolved and particulate organic matter varied with riparian land use in tropical coastal headstreams, Sci. Total Environ., 908, 168577, https://doi.org/10.1016/j.scitotenv.2023.168577, 2024.
Kemp, J. E., Lovatt, R. J., Bahr, J. C., Kahler, C. P., and Appelman, C. N.: Pre-clearing vegetation of the coastal lowlands of the Wet Tropics Bioregion, North Queensland, Cunninghamia, 10, 285–329, 2007.
Kroon, F. J., Thorburn, P., Schaffelke, B., and Whitten, S.: Towards protecting the Great Barrier Reef from land-based pollution, Glob. Change Biol., 22, 1985–2002, https://doi.org/10.1111/gcb.13262, 2016.
Kroon, F. J., Kuhnert, P. M., Henderson, B. L., Wilkinson, S. N., Kinsey-Henderson, A., Abbott, B., Brodie, J. E., and Turner, R. D. R.: River loads of suspended solids, nitrogen, phosphorus and herbicides delivered to the Great Barrier Reef lagoon, Mar. Pollut. Bull., 65, 167–181, https://doi.org/10.1016/j.marpolbul.2011.10.018, 2012.
Lambert, T., Teodoru, C. R., Nyoni, F. C., Bouillon, S., Darchambeau, F., Massicotte, P., and Borges, A. V.: Along-stream transport and transformation of dissolved organic matter in a large tropical river, Biogeosciences, 13, 2727–2741, https://doi.org/10.5194/bg-13-2727-2016, 2016.
Lee, S.-C., Shin, Y., Jeon, Y.-J., Lee, E.-J., Eom, J.-S., Kim, B., and Oh, N.-H.: Optical properties and 14C ages of stream DOM from agricultural and forest watersheds during storms, Environ. Pollut., 272, 116412, https://doi.org/10.1016/j.envpol.2020.116412, 2021.
Li, P., Wu, M., Li, T., Dumbrell, A. J., Saleem, M., Kuang, L., Luan, L., Wang, S., Li, Z., and Jiang, J.: Molecular weight of dissolved organic matter determines its interactions with microbes and its assembly processes in soils, Soil Biol. Biochem., 184, 109117, https://doi.org/10.1016/j.soilbio.2023.109117, 2023.
Liu, M., Raymond, P. A., Lauerwald, R., Zhang, Q., Trapp-Müller, G., Davis, K. L., Moosdorf, N., Xiao, C., Middelburg, J. J., Bouwman, A. F., Beusen, A. H. W., Peng, C., Lacroix, F., Tian, H., Wang, J., Li, M., Zhu, Q., Cohen, S., van Hoek, W. J., Li, Y., Li, Y., Yao, Y., and Regnier, P.: Global riverine land-to-ocean carbon export constrained by observations and multi-model assessment, Nat. Geosci., 17, 896–904, https://doi.org/10.1038/s41561-024-01524-z, 2024.
Liu, S., Kuhn, C., Amatulli, G., Aho, K., Butman, D. E., Allen, G. H., Lin, P., Pan, M., Yamazaki, D., Brinkerhoff, C., Gleason, C., Xia, X., and Raymond, P. A.: The importance of hydrology in routing terrestrial carbon to the atmosphere via global streams and rivers, P. Natl. Acad. Sci. USA, 119, e2106322119, https://doi.org/10.1073/pnas.2106322119, 2022.
Mann, P. J., Eglinton, T. I., McIntyre, C. P., Zimov, N., Davydova, A., Vonk, J. E., Holmes, R. M., and Spencer, R. G. M.: Utilization of ancient permafrost carbon in headwaters of Arctic fluvial networks, Nat. Commun., 6, 7856, https://doi.org/10.1038/ncomms8856, 2015.
Marwick, T. R., Tamooh, F., Teodoru, C. R., Borges, A. V., Darchambeau, F., and Bouillon, S.: The age of river-transported carbon: A global perspective, Global Biogeochem. Cy., 29, 122–137, https://doi.org/10.1002/2014GB004911, 2015.
Mayorga, E., Aufdenkampe, A. K., Masiello, C. A., Krusche, A. V., Hedges, J. I., Quay, P. D., Richey, J. E., and Brown, T. A.: Young organic matter as a source of carbon dioxide outgassing from Amazonian rivers, Nature, 436, 538–541, https://doi.org/10.1038/nature03880, 2005.
McDonough, L. K., Rutlidge, H., O'Carroll, D. M., Andersen, M. S., Meredith, K., Behnke, M. I., Spencer, R. G. M., McKenna, A. M., Marjo, C. E., Oudone, P., and Baker, A.: Characterisation of shallow groundwater dissolved organic matter in aeolian, alluvial and fractured rock aquifers, Geochim. Cosmochim. Ac., 273, 163–176, https://doi.org/10.1016/j.gca.2020.01.022, 2020.
McDonough, L. K., Andersen, M. S., Behnke, M. I., Rutlidge, H., Oudone, P., Meredith, K., O'Carroll, D. M., Santos, I. R., Marjo, C. E., Spencer, R. G. M., McKenna, A. M., and Baker, A.: A new conceptual framework for the transformation of groundwater dissolved organic matter, Nat. Commun., 13, 2153, https://doi.org/10.1038/s41467-022-29711-9, 2022.
McKergow, L. A., Prosser, I. P., Hughes, A. O., and Brodie, J.: Sources of sediment to the Great Barrier Reef World Heritage Area, Mar. Pollut. Bull., 51, 200–211, https://doi.org/10.1016/j.marpolbul.2004.11.029, 2005.
Miranda, F. S. D. M. E. and Avelar, A. D. S.: Dissolved organic carbon from the forest floor with different decomposition rates in a rainforest in south-eastern Brazil, Soil Res., 60, 50–64, https://doi.org/10.1071/SR21008, 2022.
Moore, S., Evans, C. D., Page, S. E., Garnett, M. H., Jones, T. G., Freeman, C., Hooijer, A., Wiltshire, A. J., Limin, S. H., and Gauci, V.: Deep instability of deforested tropical peatlands revealed by fluvial organic carbon fluxes, Nature, 493, 660–663, https://doi.org/10.1038/nature11818, 2013.
Moyer, R. P., Bauer, J. E., and Grottoli, A. G.: Carbon isotope biogeochemistry of tropical small mountainous river, estuarine, and coastal systems of Puerto Rico, Biogeochemistry, 112, 589–612, https://doi.org/10.1007/s10533-012-9751-y, 2013.
Müller, D., Warneke, T., Rixen, T., Müller, M., Jamahari, S., Denis, N., Mujahid, A., and Notholt, J.: Lateral carbon fluxes and CO2 outgassing from a tropical peat-draining river, Biogeosciences, 12, 5967–5979, https://doi.org/10.5194/bg-12-5967-2015, 2015.
Munksgaard, N. C., Wurster, C. M., and Bird, M. I.: Continuous analysis of δ18O and δD values of water by diffusion sampling cavity ring-down spectrometry: a novel sampling device for unattended field monitoring of precipitation, ground and surface waters, Rapid Commun. Mass Sp., 25, 3706–3712, https://doi.org/10.1002/rcm.5282, 2011.
Ohno, T.: Fluorescence Inner-Filtering Correction for Determining the Humification Index of Dissolved Organic Matter, Environ. Sci. Technol., 36, 742–746, https://doi.org/10.1021/es0155276, 2002.
Pucher, M., Wünsch, U., Weigelhofer, G., Murphy, K., Hein, T., and Graeber, D.: staRdom: Versatile software for analyzing spectroscopic data of dissolved organic matter in R, Water, 11, 2366, https://doi.org/10.3390/w11112366, 2019.
QGIS Development Team: QGIS Geographic Information System, Open Source Geospatial Foundation Project [code], https://github.com/qgis/QGIS (last access: 4 March 2026), 2019.
Remondi, F., Botter, M., Burlando, P., and Fatichi, S.: Variability of transit time distributions with climate and topography: A modelling approach, J. Hydrol., 569, 37–50, https://doi.org/10.1016/j.jhydrol.2018.11.011, 2019.
Rhyner, T. M. Y., Bröder, L., White, M. E., Mittelbach, B. V. A., Brunmayr, A., Hagedorn, F., Storck, F. R., Passera, L., Haghipour, N., Zobrist, J., and Eglinton, T. I.: Radiocarbon signatures of carbon phases exported by Swiss rivers in the Anthropocene, Philos. T. R. Soc. A, 381, 20220326, https://doi.org/10.1098/rsta.2022.0326, 2023.
Rocher-Ros, G., Stanley, E. H., Loken, L. C., Casson, N. J., Raymond, P. A., Liu, S., Amatulli, G., and Sponseller, R. A.: Global methane emissions from rivers and streams, Nature, 621, 530–535, https://doi.org/10.1038/s41586-023-06344-6, 2023.
Rosentreter, J. A. and Eyre, B. D.: Alkalinity and dissolved inorganic carbon exports from tropical and subtropical river catchments discharging to the Great Barrier Reef, Australia, Hydrol. Process., 34, 1530–1544, https://doi.org/10.1002/hyp.13679, 2020.
Sanderman, J., Lohse, K. A., Baldock, J. A., and Amundson, R.: Linking soils and streams: Sources and chemistry of dissolved organic matter in a small coastal watershed, Water Resour. Res., 45, https://doi.org/10.1029/2008WR006977, 2009.
Schlesinger, W. H.: Changes in soil carbon storage and associated properties with disturbance and recovery, in: The Changing Carbon Cycle: A Global Analysis, edited by: Trabalka, J. R., and Reichle, D. E., Springer New York, New York, NY, 194–220, https://doi.org/10.1007/978-1-4757-1915-4_11, 1986.
Shi, Z., Allison, S. D., He, Y., Levine, P. A., Hoyt, A. M., Beem-Miller, J., Zhu, Q., Wieder, W. R., Trumbore, S., and Randerson, J. T.: The age distribution of global soil carbon inferred from radiocarbon measurements, Nat. Geosci., 13, 555–559, https://doi.org/10.1038/s41561-020-0596-z, 2020.
Sickman, J. O., DiGiorgio, C. L., Lee Davisson, M., Lucero, D. M., and Bergamaschi, B.: Identifying sources of dissolved organic carbon in agriculturally dominated rivers using radiocarbon age dating: Sacramento–San Joaquin River Basin, California, Biogeochemistry, 99, 79–96, https://doi.org/10.1007/s10533-009-9391-z, 2010.
Solano, V., Duvert, C., Birkel, C., Maher, D. T., García, E. A., and Hutley, L. B.: Stream respiration exceeds CO2 evasion in a low-energy, oligotrophic tropical stream, Limnol. Oceanogr., 68, 1132–1146, https://doi.org/10.1002/lno.12334, 2023.
Solano, V., Duvert, C., Hutley, L. B., Cendón, D. I., Maher, D. T., and Birkel, C.: Seasonal wetlands make a relatively limited contribution to the dissolved carbon pool of a lowland headwater tropical stream, J. Geophys. Res.-Biogeo., 129, e2023JG007556, https://doi.org/10.1029/2023JG007556, 2024.
Spencer, R. G. M., Hernes, P. J., Aufdenkampe, A. K., Baker, A., Gulliver, P., Stubbins, A., Aiken, G. R., Dyda, R. Y., Butler, K. D., Mwamba, V. L., Mangangu, A. M., Wabakanghanzi, J. N., and Six, J.: An initial investigation into the organic matter biogeochemistry of the Congo River, Geochim. Cosmochim. Ac., 84, 614–627, https://doi.org/10.1016/j.gca.2012.01.013, 2012.
Spencer, R. G. M., Mann, P. J., Dittmar, T., Eglinton, T. I., McIntyre, C., Holmes, R. M., Zimov, N., and Stubbins, A.: Detecting the signature of permafrost thaw in Arctic rivers, Geophys. Res. Lett., 42, 2830–2835, https://doi.org/10.1002/2015GL063498, 2015.
Stubbins, A., Spencer, R. G. M., Chen, H., Hatcher, P. G., Mopper, K., Hernes, P. J., Mwamba, V. L., Mangangu, A. M., Wabakanghanzi, J. N., and Six, J.: Illuminated darkness: Molecular signatures of Congo River dissolved organic matter and its photochemical alteration as revealed by ultrahigh precision mass spectrometry, Limnol. Oceanogr., 55, 1467–1477, https://doi.org/10.4319/lo.2010.55.4.1467, 2010.
Stuiver, M. and Polach, H. A.: Discussion Reporting of 14C Data, Radiocarbon, 19, 355–363, https://doi.org/10.1017/S0033822200003672, 1977.
Tetzlaff, D., Seibert, J., McGuire, K. J., Laudon, H., Burns, D. A., Dunn, S. M., and Soulsby, C.: How does landscape structure influence catchment transit time across different geomorphic provinces?, Hydrol. Process., 23, 945–953, https://doi.org/10.1002/hyp.7240, 2009.
Tittel, J., Müller, C., Schultze, M., Musolff, A., and Knöller, K.: Fluvial radiocarbon and its temporal variability during contrasting hydrological conditions, Biogeochemistry, 126, 57–69, https://doi.org/10.1007/s10533-015-0137-9, 2015.
Tittel, J., Musolff, A., Rinke, K., and Büttner, O.: Anthropogenic transformation disconnects a lowland river from contemporary carbon stores in its catchment, Ecosystems, 25, 618–632, https://doi.org/10.1007/s10021-021-00675-z, 2022.
Vanclay, J. K.: Lessons from the Queensland Rainforests, J. Sustain. Forest., 3, 1–27, https://doi.org/10.1300/J091v03n02_01, 1996.
Van Gaelen, N., Verschoren, V., Clymans, W., Poesen, J., Govers, G., Vanderborght, J., and Diels, J.: Controls on dissolved organic carbon export through surface runoff from loamy agricultural soils, Geoderma, 226–227, 387–396, https://doi.org/10.1016/j.geoderma.2014.03.018, 2014.
Van Stan, J. T. and Stubbins, A.: Tree-DOM: Dissolved organic matter in throughfall and stemflow, Limnol. Oceanogr. Lett., 3, 199–214, https://doi.org/10.1002/lol2.10059, 2018.
Waldron, S., Vihermaa, L., Evers, S., Garnett, M. H., Newton, J., and Henderson, A. C. G.: C mobilisation in disturbed tropical peat swamps: old DOC can fuel the fluvial efflux of old carbon dioxide, but site recovery can occur, Sci. Rep., 9, 11429, https://doi.org/10.1038/s41598-019-46534-9, 2019.
Wilcken, K. M., Hotchkis, M., Levchenko, V., Fink, D., Hauser, T., and Kitchen, R.: From carbon to actinides: A new universal 1MV accelerator mass spectrometer at ANSTO, Nuclear Instruments and Methods in Physics Research Section B: Beam Interactions with Materials and Atoms, 361, 133–138, https://doi.org/10.1016/j.nimb.2015.04.054, 2015.
Wilson, H. F. and Xenopoulos, M. A.: Ecosystem and seasonal control of stream dissolved organic carbon along a gradient of land use, Ecosystems, 11, 555–568, https://doi.org/10.1007/s10021-008-9142-3, 2008.
Xenopoulos, M. A., Barnes, R. T., Boodoo, K. S., Butman, D., Catalán, N., D'Amario, S. C., Fasching, C., Kothawala, D. N., Pisani, O., Solomon, C. T., Spencer, R. G. M., Williams, C. J., and Wilson, H. F.: How humans alter dissolved organic matter composition in freshwater: relevance for the Earth's biogeochemistry, Biogeochemistry, 154, 323–348, https://doi.org/10.1007/s10533-021-00753-3, 2021.