the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 27 May 2026
| 27 May 2026
Limited iron isotope variation among tissues of a marine fish: a case study of wild chub mackerel (Scomber japonicus)
Nanako Hasegawa
Yoshio Takahashi
Takaaki Itai
Iron homeostasis in marine organisms operates under chronically low iron bioavailability, which may shape the strategies of iron uptake and storage in fish. Stable iron isotope ratios (δ56Fe) have emerged as tracers of iron storage and uptake in terrestrial mammals, yet the physiological drivers of isotope fractionation in marine organisms remain poorly understood. Here, we investigated δ56Fe variation and iron speciation across eight tissues of wild chub mackerel (Scomber japonicus), along with pool size estimation of key Fe species, including ferritin-bound (ferric) and heme-bound (mainly ferrous) Fe, using Fe K-edge X-ray Absorption Near Edge Structure (XANES) spectroscopy. In all the specimens, the liver δ56Fe values were higher than the average value of all tissues, with an apparent isotopic shift between ferritin- and heme-bound Fe (Δ56Fe) in the liver averaging 2.72 ‰ ± 3.03 ‰ (2 S.D.). In contrast to the liver, no enrichment of heavy Fe isotope was observed in the ovary and red muscle despite their high ferritin-Fe contribution, suggesting a high interconversion rate between ferritin- and heme-bound Fe pools in these tissues. The overall range of δ56Fe variation among tissues was smaller than that reported in mammals. Our results suggested that muscular δ56Fe in marine teleost is primarily governed by source signatures and intestinal uptake efficiency, while tissue heterogeneity due to heavy Fe storage by ferritin exerts only a minor influence. These findings highlight the potential of δ56Fe as a proxy for intestinal iron acquisition in fish and provide new geochemical perspectives on iron cycling through marine food webs.
- Article
(2744 KB) - Full-text XML
-
Supplement
(1150 KB) - BibTeX
- EndNote




Iron is an essential element for life, functioning in electron transfer through the redox cycling between Fe2+ and Fe3+ (Georgiadis et al., 1992; Hoffbrand et al., 1976; Robbins and Pederson, 1970). While soluble Fe2+ is highly bioavailable, insoluble Fe3+ dominates under atmospheric conditions (Crichton and Boelaert, 2001), resulting in dissolved iron concentrations below 2 nmol L−1 in the open ocean (Hatta et al., 2015; Johnson et al., 1997; Martin and Gordon, 1988; Nishioka and Obata, 2017). This scarcity constitutes a major factor limiting primary production by phytoplankton (Boyd et al., 2000; Martin et al., 1989; Martin and Gordon, 1988).
Although iron limitation in primary producers is well established, its implications for higher organisms such as fish remain less understood. Unlike phytoplankton, tissue iron concentrations in fish are not directly constrained by seawater iron availability (Galbraith et al., 2019). This is based on long-term iron retention strategies via ferritin (iron storage protein) and efficient intestinal absorption mechanisms that specifically exist in marine fish (Bury et al., 2001; Carriquiriborde et al., 2004; Kwong and Niyogi, 2008). However, direct quantification of intestinal iron uptake has been largely restricted to laboratory animals using enriched isotopes (van den Heuvel et al., 1998; Fiorito et al., 2012), an approach that is not feasible for wild species especially marine fish.
Natural iron stable isotope ratios (δ56Fe) offer a broadly applicable, non-lethal proxy for assessing iron metabolism. δ56Fe values are sensitive to intestinal absorption processes (Albalat et al., 2021; Flórez et al., 2017; Van Heghe et al., 2013; Hotz and Walczyk, 2013; Krayenbuehl et al., 2005; Stenberg et al., 2003) as well as to variation in iron storage and homeostasis, with ferritin-bound iron often implicated as a key driver of tissue-specific fractionation in mammals (Van Heghe et al., 2014; Hotz et al., 2012; Jaouen and Balter, 2014). Extending this framework to fish requires evaluating both intestinal absorption and ferritin-associated storage effects.
In this study, we examined δ56Fe distributions across tissues of marine fish chub mackerel (Scomber japonicus). Additionally, X-ray Absorption Near-edge Structure (XANES) spectroscopy provided a non-destructive and oxidation state–sensitive approach to quantify ferritin-bound iron, allowing direct estimation of the proportion of Fe3+ species in tissues without chemical extraction. Using this method, we quantified the relative contributions of ferritin- and heme-bound iron and assessed their roles in isotopic variability. By further considering physiological factors such as sex and reproductive status, our study provides new insights into the mechanisms governing δ56Fe fractionation in fish and its potential as a tracer of intestinal iron acquisition in marine ecosystems.
2.1 Sample Collection and Preparation
Six adult chub mackerel (females: Mk-1 to Mk-3, males: Mk-4 to Mk-6) were collected by longline fishing around Tsushima Island, East China Sea, on 27 April 2022. Fish were frozen onboard at −30 °C, stored at −20 to −25 °C for eight days, and subsequently transported to the laboratory at −15 °C. Body weight and fork length were recorded prior to dissection. Total blood volume was estimated as 60 mL kg−1 according to teleost values (Brill et al., 1998; Itazawa et al., 1983). Each individual was dissected into eight tissues (red and white muscle, liver, gonad, spleen, heart, gills, and blood). All tissue samples were stored at −80 °C until the analysis. Wet weight was measured, and hepatosomatic index (HSI) and gonadosomatic index (GSI) were calculated as follows (Allaya et al., 2013; Shiraishi et al., 2008):
Tissues with high iron content (red muscle, liver, gonad, blood, and heart) were selected for Fe K-edge XANES analysis. Small frozen fragments were immediately sealed in oxygen-impermeable polycarbonate film (ASONE, Japan) after dissection and stored until measurement.
2.2 Iron Concentration and Stable Isotope Measurement
Analytical procedures followed Hasegawa et al. (2023). Tissues were freeze-dried, and lipids removed from liver and gonad using chloroform : methanol mixture (2:1, ). Tests with certified reference materials confirmed no significant isotopic alteration during lipid removal (<0.07 ‰, n=4). Dried tissues (0.3–2.5 g) were digested in perfluoroalkoxy (PFA) vials using 68 % HNO3 and 35 % H2O2 (both Tamapure AA-100, Tama Chemical Corp., Japan) at 120 °C. The digests were evaporated to dryness and re-dissolved in 7 M HCl containing 0.001 % H2O2. Each sample was split for iron concentration analysis by inductively-coupled plasma mass spectroscopy (ICP-MS; Agilent 7700) and another aliquot for isotope analysis. Iron concentrations were determined from a single aliquot of each tissue sample.
Iron purification was performed using anion-exchange chromatography following Maréchal et al. (1999). Interfering elements (Ni, Cr, Cu) were removed with 7 M HCl, and Fe was subsequently eluted with 2 M HCl. The purified Fe fraction was dissolved in 2 % HNO3 at a concentration of 1 µg mL−1, with Cu added for mass bias correction.
Stable isotope ratios were determined by multicollector-ICP-MS (Neptune Plus, ThermoFisher Scientific) at The University of Tokyo. Instrumental settings were: RF power 1200 W; Ar gas flow 15 L min−1 (cool), 0.7 L min−1 (auxiliary), 1.0 L min−1 (nebulizer). Signal intensity was 5–8 V for 1 µg mL−1 56Fe+, with blanks below 0.004 V. Each tissue sample was measured once for iron isotope composition with 60 analytical cycles. The δ56Fe value was expressed relative to IRMM-014 as:
Whole-body δ56Fe (δWB) was calculated as:
where i represents the ith tissue, and Ti, Ci, and Mi represent the Fe burden, Fe concentration, and tissue mass, respectively.
Analytical accuracy was verified using certified reference materials (BCR-414, DORM-4, DOLT-5, ERM-CE464). Results were consistent with reported values: BCR-414 (−0.10 ‰ ± 0.03 ‰), DORM-4 (−0.24 ‰ ± 0.06 ‰), DOLT-5 (−2.32 ‰ ± 0.02 ‰), ERM-CE464 (−0.55 ‰ ± 0.02 ‰).
2.3 XANES Spectroscopy
Iron K-edge XANES was analyzed at BL-12C, Photon Factory (KEK, Japan) following the protocol of Hasegawa et al. (2023). Frozen tissue fragments were analyzed in fluorescence mode using a Si detector, with energy calibration against hematite (7110.2 eV). The number of independent spectral components was determined by principal component analysis (PCA).
The dominant Fe species in animal tissues include heme (Fe–porphyrin), ferritin (storage), and transferrin (Fe transport). Transferrin-bound Fe was excluded from further analysis due to minor abundance and negligible isotope fractionation (Walczyk and von Blanckenburg, 2005). Thus, subsequent analyses focused on ferritin- and heme-bound Fe species. Linear combination fitting (LCF) was performed to quantify their relative proportions in each tissue.
To account for possible post-mortem oxidation of hemoproteins, spectra of oxyHb, deoxyHb, and metHb were included in the fitting. Horse spleen ferritin (Hs-Ft) and bovine blood metHb (Bb-metHb) were used as reference standards, while oxyHb and deoxyHb were prepared from metHb following Di Iorio (1981) and Wilson et al. (2013). Pre-edge features were extracted and fitted using pseudo-Voigt functions implemented in Python. The quality of spectra fits was evaluated using the R-factor, defined as:
2.4 Statistical Analysis
Statistical analyses were conducted in Python (ver. 3.9.19). Sex differences in physiological traits, Fe concentration, and isotope ratios were tested using the Mann–Whitney U test.
3.1 Physiological Parameters of Chub Mackerel
The six chub mackerel analyzed had fork lengths (mean ± 2 S.D.) of 37.7 ± 2.41 cm in females and 40.9 ± 4.00 cm in males, and corresponding body weights of 0.71 ± 0.30 kg and 0.89 ± 0.24 kg, respectively (Table 1). Although females tended to be slightly smaller, the difference in body length was not significant (p=0.06). Based on the growth curve of Shiraishi et al. (2008), all individuals were estimated to be at least four years old and sexually mature. The hepatosomatic index (HSI) ranged from 1.05–2.14 in females and 0.74–1.26 in males, and the gonadosomatic index (GSI) ranged from 4.26–10.1 in females and 7.25–10.0 in males, consistent with the spawning season in the East China Sea (Shiraishi et al., 2008). Among tissues, white muscle represented the largest biomass fraction (31 %–52 % of total body weight; Fig. S1 in the Supplement), followed by gonads (3.8 %–8.9 %), red muscle (4.5 %–7.7 %), gills (2.1 %–3.9 %), liver (0.4 %–1.9 %), heart (0.2 %–0.6 %), and spleen (0.2 %–0.4 %). No significant sex-related differences were observed (p>0.05). Red and white muscles, as well as gills, exhibited strong positive correlations with body weight (r>0.82, p<0.04), whereas gonads showed a positive but non-significant trend (r=0.77, p=0.07; Fig. S2).
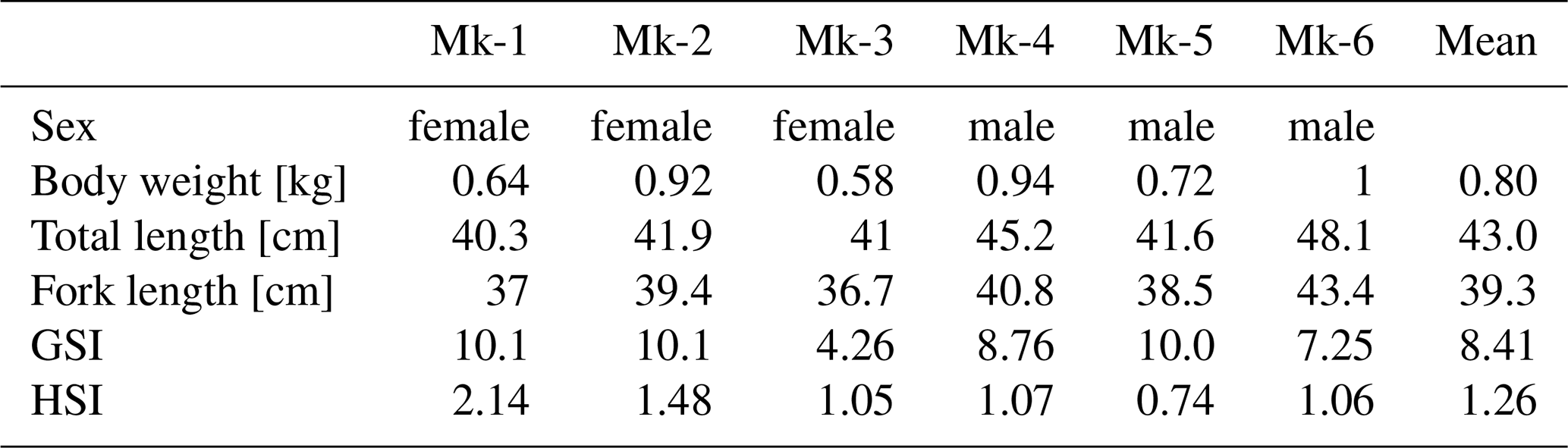
3.2 Tissue Iron Concentrations and Stable Isotope Ratios
Mean iron concentrations (±2 S.D.) were highest in the spleen (3100 ± 860 ), followed by blood (1300 ± 480 ), heart (720 ± 580 ), liver (630 ± 510 ), gills (290 ± 90 ), red muscle (250 ± 84 ), gonads (37 ± 24 ), and white muscle (15 ± 10 ) (Fig. 1A). Male spleens tended to have higher Fe concentrations than those of females, and liver and gonads also tended to be higher in males.
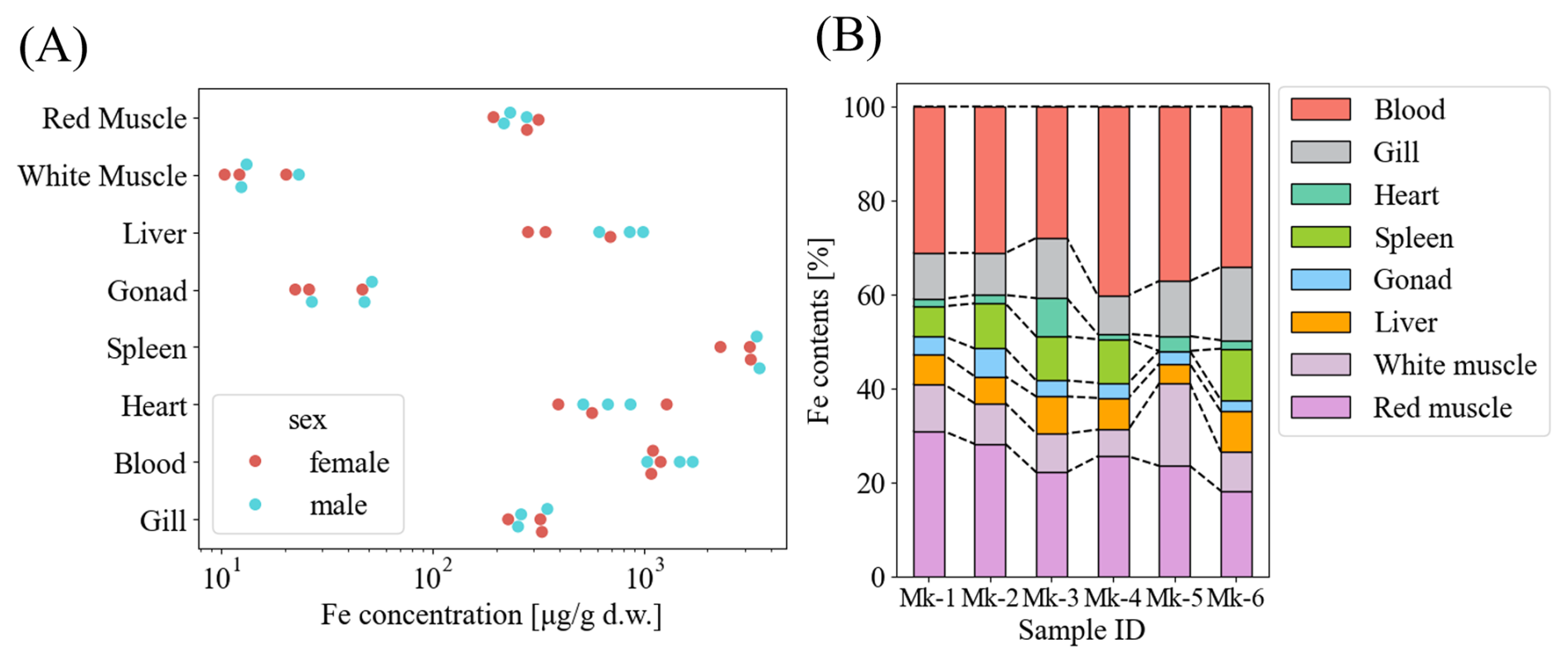
Figure 1(A) Iron concentration, and (B) Tissue iron burden of the chub mackerels (Mk-1 to Mk-3: female, Mk-4 to Mk-6: male). Total blood contents were assumed to be 30 mL kg−1 with a specific gravity of 1.05. No uncertainty is reported as multiple sample replicates were not measured. The measurement uncertainty was assumed to be zero.
Assuming a blood volume of 30 mL kg−1 and 1.05 as the specific gravity of fish blood (Davison, 2011), the total iron content in blood was estimated to be between 3.2 and 10.3 mg (Table S1 in the Supplement). Red muscle contributed the next largest Fe pool (2.3–6.1 mg), followed by gills (1.3–3.7 mg), white muscle (0.9–2.0 mg), spleen (0.8–2.6 mg), liver (0.5–2.0 mg), gonads (0.3–1.0 mg), and heart (0.2–0.9 mg). The total body iron inventory was therefore estimated to be approximately 17–26 mg kg−1. Although the liver is the principal iron storage tissue, it accounted for only 4 %–8 % of total body iron. In contrast, blood and red muscle together comprised 51 %–67 % of the total iron burden.
Mean iron isotope compositions (δ56Fe ± 2 S.D.) of each tissue across individuals were as follows: red muscle, −1.54 ‰ ± 0.20 ‰; white muscle, −1.46 ‰ ± 0.21 ‰; liver, −1.19 ‰ ± 0.16 ‰; gonads, −1.23 ‰ ± 0.42 ‰; spleen, −1.37 ‰ ± 0.15 ‰; heart, −1.43 ‰ ± 0.19 ‰; gills, −1.37 ‰ ± 0.22 ‰; and blood, −1.39 ‰ ± 0.20 ‰ (Fig. 2). The net δ56Fe in whole mackerel bodies weighted by tissue Fe contents ranged from −1.50 ‰ to −1.35 ‰ with the liver in all individuals showing higher values than the net values. No sex differences were detected, although ovarian δ56Fe was lower than testicular values.
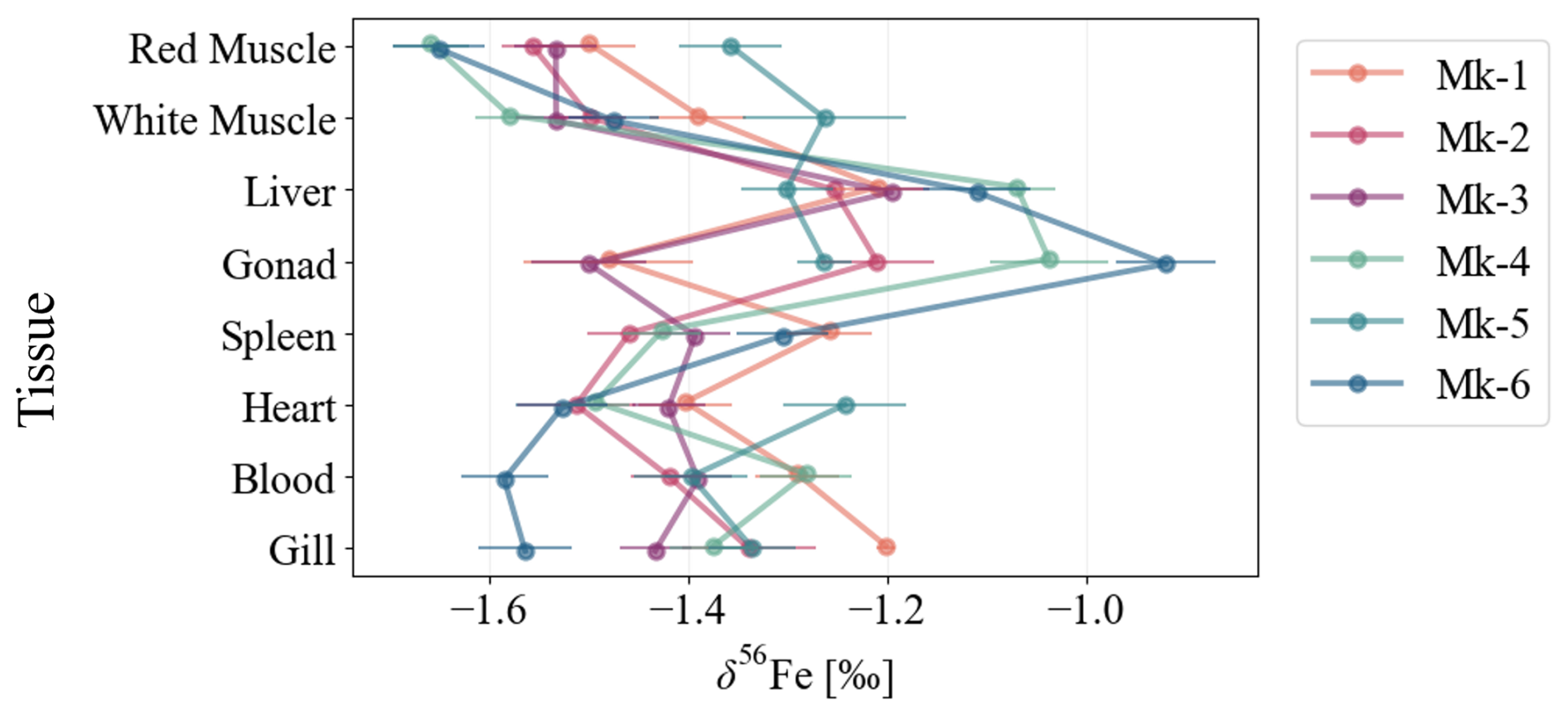
Figure 2Iron stable isotope ratio in tissues of the six Scomber japonicus individuals (Mk1–3: females, Mk4–6: males). Error bars represent 2 S.E. during a single measurement (60 analytical cycles).
3.3 Proportion of Ferritin-Bound Iron
Iron K-edge XANES spectra identified the dominant chemical forms of iron in each tissue (Wilke et al., 2001). Pre-edge peak positions for all samples were distributed between ferritin (most oxidized) and hemoglobin (reduced) reference standards (Fig. 3A). We note that the XANES spectrum of MK-2 liver was unavailable because its quality was not good.
Figure 3(A) Fe K-edge XANES spectra of Scomber japonicus tissues (Mk-1 and Mk-3: female, Mk-4 to Mk-6: male). Mk-1 and Mk-3 represent females, and Mk-4 to Mk-6 represent males. The standard spectra were obtained from ferritin from horse spleen and hemoglobin derivatives from bovine blood. Blue lines indicate the spectra of the standard materials, while black lines represent the tissue samples. Red dashed lines show the results of linear combination fitting (LCF) using the four standard spectra. Gray solid lines correspond to the first derivative of the normalized XANES spectra, which highlights the peak top energies, and gray dashed lines mark the peak top energy of Ferritin at 7129 eV. (B) Estimated proportions of each standard materials with linear combination fitting (LCF) of XANES spectra. R value is a goodness-of-fit parameter described in Eq. (4). Each number on the pie charts represents proportions (%) of each component.
Linear combination fitting (LCF) using Hs-Ft and Bb-Hb derivatives revealed that red muscle and heart were dominated by heme-bound Fe, whereas ferritin accounted for 8 %–31 % of total Fe in the liver (Fig. 3B). One individual exhibited extremely low liver ferritin iron (8 %). The liver ferritin fraction was comparable to those reported for chub mackerel from other regions (28 %; Hasegawa et al., 2023) but substantially lower than those of mouse liver (66 %; Chen and Chen, 2018).
Sex-related differences were also apparent: females exhibited lower ferritin-bound Fe proportions in the liver, while higher proportions in the red muscle and gonads than males. In ovaries, ferritin represented the predominant Fe form (70 %–80 %), whereas testes showed highly variable proportions (3 %–59 %).
4.1 Elevated δ56Fe in Liver and Contribution of Ferritin-Bound Iron
First, we focused on the liver because it is a central tissue for iron storage in vertebrates and is expected to exhibit iron isotope fractionation through the synthesis of ferritin. All the chub mackerel individuals showed higher δ56Fe values in the liver than δWB across eight tissues. Because ferritin preferentially incorporates heavier Fe isotopes during the oxidation in the storage process according to human studies (Albarède et al., 2011), this enrichment most likely reflects the contribution of ferritin-bound iron within the liver. It should be noted that human ferritin consists of H and L subunits responsible for iron oxidation and mineralization (Plays et al., 2021). While teleost ferritin includes an additional M-type subunit with intermediate functional properties, its effect on iron oxidation and isotope fractionation remains unclear and is not considered further here (Dickey et al., 1987; Andersen et al., 1995).
The proportion of ferritin-bound Fe in mackerel liver (8 %–31 %) was notably lower than that reported for mammalian liver (∼60 %; Chakrabarti et al., 2015; Chen and Chen, 2018), suggesting that a smaller fraction of hepatic Fe is stored as ferritin in teleost. Consequently, the δ56Fe enrichment in the mackerel liver appears more moderate than in mammals, possibly due to a greater influence of isotopically lighter Fe pools such as circulating hemoglobin iron.
Because blood contains little ferritin-bound iron, the elevated liver δ56Fe cannot be explained by residual blood contamination. Given that iron cycling within the body is a semi-closed system, in which the liver acts as the central exchange site between storage (ferritin) and transport (hemoglobin and transferrin) pools (Slusarczyk and Mleczko-Sanecka, 2021), hepatic δ56Fe can be interpreted as a mixture of these isotopically distinct components. This relationship can be expressed as:
where p is the fraction of ferritin-bound Fe in the liver, and δFt and δHm represent the isotopic compositions of ferritin- and heme-bound Fe, respectively.
By assuming that the lowest δ56Fe observed here in red muscle represents δHm (−1.66 ‰), the isotopic offset between ferritin- and heme-bound Fe in the liver () was estimated to be 2.72 ‰ ± 3.03 ‰ (2 S.D.) in all specimens, whereas it was 4.15 ‰ ± 2.84 ‰ (2 S.D.) in females and 1.76 ‰ ± 0.88 ‰ in males. These estimated values are higher than previous observations in skipjack tuna (Katsuwonus pelamis, female, Δ=1.52 ‰, n=1; Hasegawa et al., 2023) and another chub mackerel specimen (Δ=1.41 ‰, male, n=1; Hasegawa et al., 2023), and, in females, appear higher than the equilibrium isotope fractionation between Fe(II) and Fe(III) (∼2.8 ‰, Johnson et al., 2002; Welch et al., 2003). The two females exhibited a lower ferritin-bound iron fraction than the three males, while their liver δ56Fe values were comparable to those of the males, resulting in a tendency for their estimated Δ values to be higher. Because of the limited sample size, it cannot be definitively concluded that a sex difference exists. However, one possible explanation is that the release of isotopically lighter Fe from hepatic ferritin following Rayleigh fractionation leads to significant enrichment of heavy Fe in the residual ferritin and results in Δ values larger than those expected under isotopic equilibrium. As the sample consists of mature individuals during the spawning season, it cannot be ruled out that physiological changes relating to spawning may have temporarily altered iron metabolism, particularly in females.
4.2 δ56Fe Variation Across Tissues
Outside the liver, some tissues did not exhibit elevated δ56Fe values despite high ferritin content. In particular, ovaries and red muscle in females exhibited high ferritin-to-heme ratios but δ56Fe values comparable to other tissues. This pattern suggests that ferritin-bound Fe in these tissues is not retained over long periods but is actively mobilized and recycled in response to metabolic activity and oxygen consumption. For example, ferritin in skeletal muscle can be quickly mobilized to meet oxygen demand (Robach et al., 2007; Ryan et al., 2021), preventing the accumulation of a heavy isotope signature and maintaining δ56Fe values close to those of the whole-body iron pool dominated by heme iron. Moreover, δ56Fe has been associated with oxidative stress markers including superoxide dismutase and glutathione peroxidase (Sauzéat et al., 2025), indicating that tissues prone to oxidative stress, such as gonads, may display isotopic variability due to redox-related iron turnover.
These observations suggest that ferritin-bound (storage) iron in chub mackerel exhibits a high turnover rate, resulting in small δ56Fe differences among tissues. In contrast, heme iron likely turns over more slowly. Fish erythrocyte lifespans range from ∼80 to 500 d (Götting and Nikinmaa, 2017; Avery et al., 1992), comparable or longer than those of mice (∼60 d, Dholakia et al., 2015) and humans (∼120 d; Shemin and Rittenberg, 1946). Marine mammals, by contrast, display extended erythrocyte lifespans that scale with diving capacity (Pearson et al., 2024), highlighting the link between oxygen storage and iron utilization. In migratory fish such as mackerel, the high and continuous demand for heme to support aerobic activity may promote rapid redistribution of heme iron, which may result in reduced δ56Fe variability among organs.
4.3 Features of δ56Fe Variation Among Marine Species
The range of δ56Fe variation among tissues in chub mackerel was clearly narrower than that reported for mammals. For example, the differences in δ56Fe values between muscle and liver range from 0.72 ‰ to 1.71 ‰ in mice (Balter et al., 2013), 1.64 ‰ to 2.72 ‰ in sheep (Balter et al., 2013), and 0.83 ‰ to 1.75 ‰ in humans (Walczyk and von Blanckenburg, 2002), whereas the maximum isotopic difference among tissues in the present chub mackerel specimens was 0.73 ‰. Despite this relatively small internal fractionation, δ56Fe values in marine organisms often show species-specific ranges (Fig. 4). For example, muscular δ56Fe values in skipjack tuna are high regardless of regional differences (−1.46 ‰ to −0.71 ‰; Hasegawa et al., 2022), whereas mammals generally show lower values (−3.79 ‰ to −1.5 ‰; Walczyk and von Blanckenburg, 2002; Balter et al., 2013). This pattern does not appear to arise from the enrichment of lighter isotopes in specific tissues, but rather from the limited influence of internal isotopic fractionation on whole-body δ56Fe. This interpretation is consistent with our previous study on a single individual (Hasegawa et al., 2023), and the limited inter-tissue δ56Fe variation observed across multiple individuals in the present work supports that this characteristic is generally applicable to chub mackerel.
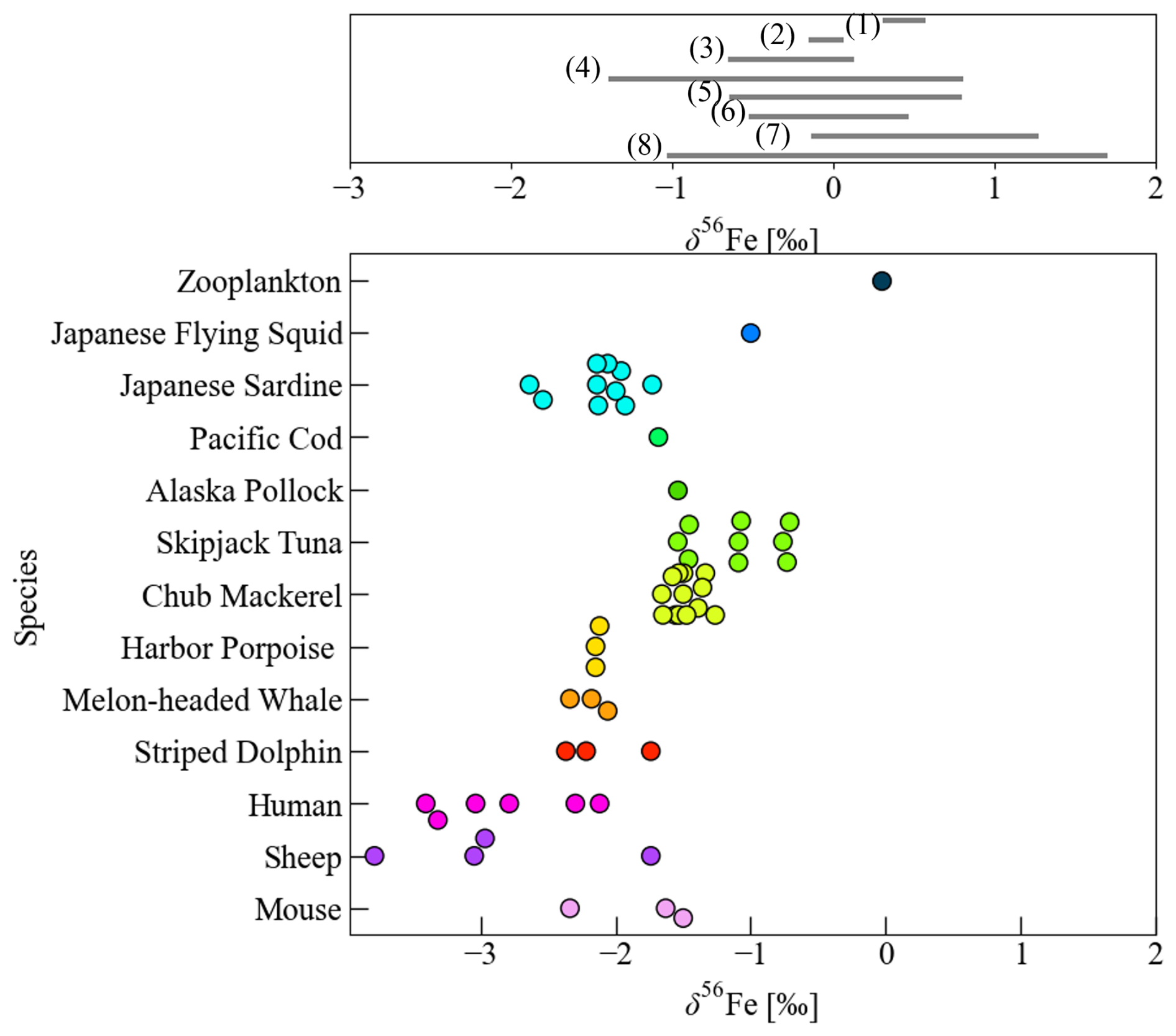
Figure 4Comparison of δ56Fe values in seawater (upper figure), and marine organisms and mammals (lower figure). Literature data in the upper figure are indicated by reference numbers: seawater from the Pacific Ocean (1–4: Radic et al., 2011; Conway and John, 2015; Pinedo-González et al., 2020; Kurisu et al., 2024), North Atlantic Ocean (5: Conway and John, 2014), and South Atlantic–Southern Ocean (6–8: Abadie et al., 2017; Ellwood et al., 2020; Sieber et al., 2021). The data of chub mackerel includes this study and Hasegawa et al. (2023), while values for zooplankton (whole body), squid (mantle), and mammals (harbor porpoise, melon-headed whale, striped dolphin, human, sheep, and mouse) are compiled from previous studies (Walczyk and von Blanckenburg, 2002; Balter et al., 2013; Hasegawa et al., 2022).
As discussed in the previous section, hepatic ferritin iron likely undergoes isotopic fractionation on the order of approximately 1.5 ‰–3 ‰, based on the Δ values estimated in this study and Hasegawa et al. (2023). However, because ferritin constitutes only a small fraction of total iron, its contribution to whole-body δ56Fe variation is limited. Consequently, heterogeneous distribution of δ56Fe across tissues is likely to have only a minor influence on the δ56Fe of indicator tissues representing whole-body δ56Fe values (e.g. muscle with a large Fe pool size). The primary determinant of tissue δ56Fe in chub mackerel therefore appears to be iron uptake process from external sources rather than internal redistribution.
The relatively high mean δ56Fe values in mackerel tissues may primarily reflect more efficient intestinal iron absorption than in mammals, resulting in a stronger reflection of environmental δ56Fe values. Additionally, selective ingestion of prey with intrinsically higher δ56Fe values could also contribute to their isotopic composition. Various prey taxa of marine fish, including zooplankton, squid, and crustaceans, are known to exhibit relatively high δ56Fe values (−1.00 ‰ to −0.03 ‰; von Blanckenburg et al., 2013; Hasegawa et al., 2022, 2023). The δ56Fe differences observed among fish groups such as tuna and mackerel (−1.58 ‰ to −0.71 ‰) versus sardine and herring (−2.64 ‰ to −1.73 ‰; Hasegawa et al., 2022) seemed difficult to explain solely by isotopic differences in their prey since all prey species analyzed so far showed higher δ56Fe values than sardine and herring (Hasegawa et al., 2022). However, recent studies have suggested species-specific δ56Fe variability even among zooplankton (Hasegawa et al., 2026) and therefore prey-derived δ56Fe may also partially contribute to the overall isotopic composition of fish such as sardines and herrings that feed selectively on specific plankton groups. Further expansion of marine biological δ56Fe databases may enable a more comprehensive understanding of Fe physiology in wild marine organisms.
In contrast, terrestrial mammals such as humans, mice, and sheep consistently show large δ56Fe differences between liver and muscle (Walczyk and von Blanckenburg, 2002; Balter et al., 2013), reflecting a well-developed iron recycling system based on the long-term ferritin storage. This mechanism reduces daily iron requirements and promotes preferential uptake of lighter isotopes, thereby lowering the overall δ56Fe values. In contrast to mammals that rely on ferritin-based iron storage and recycling, the isotopic pattern observed in fish may indicate that long-term iron storage plays a less dominant role in iron homeostasis, despite inhabiting chronically iron-limited environments. They may instead have evolved a “recycle dominant” mode of iron homeostasis that relies on intensive recycling of iron to sustain metabolic demands.
4.4 Implications for biological iron cycling in marine environments
The results of this study suggest that the δ56Fe variability in marine fish reflects dietary characteristics and uptake efficiency more strongly than internal physiological variations. The distinctly lighter isotopic signatures of animals compared to seawater may ultimately influence the isotopic composition of iron recycled through marine food webs. Therefore, whole-body δ56Fe values could serve as an integrated tracer of biologically available iron within marine ecosystems. Our analyses also indicate that in fish, δ56Fe values in non-destructive tissues such as blood can represent the whole-body isotopic composition, highlighting their potential as key species for assessing iron supply and cycling in higher marine ecosystems.
In the future, applying a Rayleigh-type model may allow quantitative evaluation of the relative contributions of absorption and retention to isotopic fractionation in fish. Combined with XAFS analyses that provide species-level insights into ferritin- and heme-bound iron pools, these approaches could offer a powerful framework for linking biological iron metabolism to marine biogeochemical iron cycling. In addition, further biochemical studies examining Fe isotope fractionation during enzymatic incorporation into iron-binding proteins such as ferritin or heme in fish would help to better constrain the mechanisms controlling Fe isotope distribution in marine organisms.
This study demonstrates that δ56Fe variations among tissues in chub mackerel are smaller than those observed in mammals. Although isotopic fractionation of around 1.5 ‰–3 ‰ may occur in liver ferritin, its effect on whole-body δ56Fe is minimal due to the limited ferritin fraction and overall liver iron content. These results suggest that the high δ56Fe of mackerel primarily reflects dietary iron sources and intestinal uptake rather than internal isotopic fractionation. The maximum δ56Fe range among major tissues was 0.73 ‰, indicating relatively small tissue isotopic variability. The small variation in δ56Fe among major tissues such as muscle and liver compared with the potential isotopic fractionation associated with ferritin formation, suggests that iron isotope pools in fish remain relatively stable. Although the number of individuals analysed in this study is limited, these δ56Fe patterns across multiple individuals provide robust constraints on iron isotope behaviour in wild marine fish. These findings provide fundamental information for studies of iron cycling and metabolism in marine organisms and have potential applications in food-web modeling and ecological assessments.
The data underlying this study are available in the article itself and its Supplement.
The supplement related to this article is available online at https://doi.org/10.5194/bg-23-3605-2026-supplement.
NH and TI contributed conceptualization, Methodology, and Project Administration; NH conducted Formal analysis, Software, Investigation, Visualization, and Writing (original draft preparation); YT and TI contributed Writing (review and editing); NH, YT and TI contributed Funding Acquisition.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
The authors appreciate Mr. Kei Zenimoto and Prof. Kotaro Shirai for collecting and transporting mackerel samples. We also thank Mr. Kotaro Hirayama and Mr. Yuma Sato for helping with the dissection of fish samples. We used ChatGPT to improve the English clarity and readability of the manuscript. This work was performed under the approval of the Photon Factory Program Advisory Committee (Proposal No. 2023G119, 2023S2-001, and 2022G126). This work was supported by the Japan Science and Technology Agency (JST) Support for Pioneering Research Initiated by the Next Generation (SPRING) program, Grant Number JPMJSP2108, and Asahi Group Foundation Grant Number A24-0057.
This research has been supported by the Japan Science and Technology Agency (grant no. JPMJSP2108) and the Asahi Group Foundation (grant no. A24-0057).
This paper was edited by Yuan Shen and reviewed by three anonymous referees.
Abadie, C., Lacan, F., Radic, A., Pradoux, C., and Poitrasson, F.: Iron isotopes reveal distinct dissolved iron sources and pathways in the intermediate versus deep Southern Ocean, P. Natl. Acad. Sci. USA, 114, 858–863, https://doi.org/10.1073/pnas.1603107114, 2017.
Albalat, E., Cavey, T., Leroyer, P., Ropert, M., Balter, V., and Loréal, O.: Hfe Gene Knock-Out in a Mouse Model of Hereditary Hemochromatosis Affects Bodily Iron Isotope Compositions, Front. Med., 8, 711822, https://doi.org/10.3389/fmed.2021.711822, 2021.
Albarède, F., Telouk, P., Lamboux, A., Jaouen, K., and Balter, V.: Isotopic evidence of unaccounted for Fe and Cu erythropoietic pathways, Metallomics, 3, 926–933, https://doi.org/10.1039/c1mt00025j, 2011.
Allaya, H., Hattour, A., Hajjej, G., and Trabelsi, M.: Biologic characteristics of Scomber japonicus (Houttuyn, 1782) in Tunisian waters (Central Mediterranean Sea), Afr. J. Biotechnol., 12, 3040, 2013.
Andersen, O., Dehli, A., Standal, H., Giskegjerde, T. A., and Karstensen, R., and Rørvik, K. A.: Two ferritin subunits of Atlantic salmon (Salmo salar): cloning of the liver cDNAs and antibody preparation, Mol. Mar. Biol. Biotech., 4, 164–170, 1995.
Avery, E. H., Lee, B. L., Freedland, R. A., and Cornelius, C. E.: Bile pigments in gallbladder and freshly-secreted hepatic duct bile from fed and fasted rainbow trout, Oncorhynchus mykiss, Comp. Biochem. Physiol. Comp. Physiol., 101, 857–861, https://doi.org/10.1016/0300-9629(92)90370-6, 1992.
Balter, V., Lamboux, A., Zazzo, A., Télouk, P., Leverrier, Y., Marvel, J., Moloney, A. P., Monahan, F. J., Schmidt, O., and Albarède, F.: Contrasting Cu, Fe, and Zn isotopic patterns in organs and body fluids of mice and sheep, with emphasis on cellular fractionation, Metallomics, 5, 1470–1482, https://doi.org/10.1039/C3MT00151B, 2013.
Boyd, P. W., Watson, A. J., Law, C. S., Abraham, E. R., Trull, T., Murdoch, R., Bakker, D. C. E., Bowie, A. R., Buesseler, K. O., Chang, H., Charette, M., Croot, P., Downing, K., Frew, R., Gall, M., Hadfield, M., Hall, J., Harvey, M., Jameson, G., Laroche, J., Liddicoat, M., Ling, R., Maldonado, M. T., Mckay, R. M., Nodder, S., Pickmere, S., Pridmore, R., Rintoul, S., Safi, K., Sutton, P., Strzepek, R., Tanneberger, K., Turner, S., Waite, A., and Zeldis, J.: A mesoscale phytoplankton bloom in the polar Southern Ocean stimulated by iron fertilization, Nature, 407, 695–702, https://doi.org/10.1038/35037500, 2000.
Brill, R. W., Cousins, K. L., Jones, D. R., Bushnell, P. G., and Steffensen, J. F.: Blood volume, plasma volume and circulation time in a high-energy-demand teleost, the yellowfin tuna (Thunnus albacares), J. Exp. Biol., 201, 647–654, https://doi.org/10.1242/jeb.201.5.647, 1998.
Bury, N. R., Grosell, M., Wood, C. M., Hogstrand, C., Wilson, R. W., Rankin, J. C., Busk, M., Lecklin, T., and Jensen, F. B.: Intestinal iron uptake in the European flounder (Platichthys flesus), J. Exp. Biol., 204, 3779–3787, https://doi.org/10.1242/jeb.204.21.3779, 2001.
Carriquiriborde, P., Handy, R. D., and Davies, S. J.: Physiological modulation of iron metabolism in rainbow trout (Oncorhynchus mykiss) fed low and high iron diets, J. Exp. Biol., 207, 75–86, https://doi.org/10.1242/jeb.00712, 2004.
Chakrabarti, M., Cockrell, A. L., Park, J., McCormick, S. P., Lindahl, L. S., and Lindahl, P. A.: Speciation of iron in mouse liver during development, iron deficiency, IRP2 deletion and inflammatory hepatitis, Metallomics, 7, 93–101, https://doi.org/10.1039/c4mt00215f, 2015.
Chen, R. and Chen, G.: Tumor-induced disorder of iron metabolism in major organs: A new insight from chemical speciation of iron, J. Int. Med. Res., 46, 70–78, https://doi.org/10.1177/0300060517718711, 2018.
Conway, T. M. and John, S. G.: Quantification of Dissolved Iron Sources to the North Atlantic Ocean, Nature, 511, 212–215, https://doi.org/10.1038/nature13482, 2014.
Conway, T. M. and John, S. G.: The Cycling of Iron, Zinc and Cadmium in the North East Pacific Ocean – Insights from Stable Isotopes, Geochim. Cosmochim. Ac., 164, 262–283, https://doi.org/10.1016/j.gca.2015.05.023, 2015.
Crichton, R. R. and Boelaert, J. R.: Inorganic biochemistry of iron metabolism: from molecular mechanisms to clinical consequences, John Wiley & Sons, https://doi.org/10.1002/0470845791, 2001.
Davison, P.: The specific gravity of mesopelagic fish from the northeastern Pacific Ocean and its implications for acoustic backscatter, ICES J. Mar. Sci., 68, 2064–2074, https://doi.org/10.1093/icesjms/fsr140, 2011.
Dholakia, U., Bandyopadhyay, S., Hod, E. A., and Prestia, K. A.: Determination of RBC survival in C57BL/6 and C57BL/6-Tg (UBC–GFP) mice, Comp. Med., 65, 196–201, 2015.
Di Iorio, E. E.: Preparation of derivatives of ferrous and ferric hemoglobin, Method. Enzymol., 76, 57–72, https://doi.org/10.1016/0076-6879(81)76114-7, 1981.
Dickey, L. F., Sreedharan, S., Theil, E. C., Didsbury, J. R., Wang, Y. H., and Kaufman, R. E.: Differences in the regulation of messenger RNA for housekeeping and specialized-cell ferritin. A comparison of three distinct ferritin complementary DNAs, the corresponding subunits, and identification of the first processed in amphibia, J. Biol. Chem., 262, 7901–7907, https://doi.org/10.1016/S0021-9258(18)47653-3, 1987.
Ellwood, M. J., Strzepek, R. F., Strutton, P. G., Trull, T. W., Fourquez, M., and Boyd, P. W.: Distinct iron cycling in a Southern Ocean eddy, Nat. Commun., 11, 825, https://doi.org/10.1038/s41467-020-14464-0, 2020.
Fiorito, V., Crich, S. G., Silengo, L., Altruda, F., Aime, S., and Tolosano, E.: Assessment of iron absorption in mice by ICP-MS measurements of 57Fe levels, Eur. J. Nutr., 51, 783–789, https://doi.org/10.1007/s00394-011-0256-6, 2012.
Flórez, M. R., Anoshkina, Y., Costas-Rodríguez, M., Grootaert, C., Van Camp, J., Delanghe, J., and Vanhaecke, F.: Natural Fe isotope fractionation in an intestinal Caco-2 cell line model, J. Anal. Atom. Spectrom., 32, 1713–1720, https://doi.org/10.1039/c7ja00090a, 2017.
Galbraith, E. D., Le Mézo, P., Hernandez, G. S., Bianchi, D., and Kroodsma, D.: Growth limitation of marine fish by low iron availability in the open ocean, Front. Mar. Sci., 6, 509, https://doi.org/10.3389/fmars.2019.00509, 2019.
Georgiadis, M. M., Komiya, H., Chakrabarti, P., Woo, D., Kornuc, J. J., and Rees, D. C.: Crystallographic Structure of the Nitrogenase Iron Protein from Azotobacter vinelandii, Science, 257, 1653–1659, https://doi.org/10.1126/science.1529353, 1992.
Götting, M. and Nikinmaa, M. J.: Transcriptomic analysis of young and old erythrocytes of fish, Front. Physiol., 8, 1046, https://doi.org/10.3389/fphys.2017.01046, 2017.
Hasegawa, N., Itai, T., Kunisue, T., and Takahashi, Y.: Variation of Iron Stable Isotopes in a Marine Ecosystem from the Northwest Pacific Ocean, Chem. Lett., 51, 556–560, https://doi.org/10.1246/cl.220099, 2022.
Hasegawa, N., Takahashi, Y., and Itai, T.: Tissue-variation of iron stable isotopes in marine fish coupled with speciation analysis using X-ray absorption fine structure, Sci. Total Environ., 881, 163449, https://doi.org/10.1016/j.scitotenv.2023.163449, 2023.
Hasegawa, N., Kurisu, M., Takahashi, K., Kodama, T., Hirayama, K., Ijichi, Y., Kashiwabara, T., Takahashi, Y., and Itai, T.: Bulk Net Zooplankton Samples Mask Species-specific Iron Isotope Signatures, ACS ES&T Water, accepted, 2026.
Hatta, M., Measures, C. I., Wu, J., Roshan, S., Fitzsimmons, J. N., Sedwick, P., and Morton, P.: An overview of dissolved Fe and Mn distributions during the 2010–2011 U. S. GEOTRACES north Atlantic cruises: GEOTRACES GA03, Deep-Sea Res. Pt. II, 116, 117–129, https://doi.org/10.1016/j.dsr2.2014.07.005, 2015.
Hoffbrand, A. V., Ganeshaguru, K., Hooton, J. W. L., and Tattersall, M. H. N.: Effect of Iron Deficiency and Desferrioxamine on DNA Synthesis in Human Cells, Brit. J. Haematol., 33, 517–526, https://doi.org/10.1111/j.1365-2141.1976.tb03570.x, 1976.
Hotz, K. and Walczyk, T.: Natural iron isotopic composition of blood is an indicator of dietary iron absorption efficiency in humans, J. Biol. Inorg. Chem., 18, 1–7, https://doi.org/10.1007/s00775-012-0943-7, 2013.
Hotz, K., Krayenbuehl, P. A., and Walczyk, T.: Mobilization of storage iron is reflected in the iron isotopic composition of blood in humans, J. Biol. Inorg. Chem., 17, 301–309, https://doi.org/10.1007/s00775-011-0851-2, 2012.
Itazawa, Y., Takeda, T., Yamamoto, K., and Azuma, T.: Determination of Circulating Blood Volume in Three Teleosts,Carp,Yellowtail and Porgy, Jpn. J. Ichthyol., 30, 94–101, https://doi.org/10.11369/jji1950.30.94, 1983.
Jaouen, K. and Balter, V.: Menopause effect on blood Fe and Cu isotope compositions, Am. J. Phys. Anthropol., 153, 280–285, https://doi.org/10.1002/ajpa.22430, 2014.
Johnson, C. M., Skulan, J. L., Beard, B. L., Sun, H., Nealson, K. H., and Braterman, P. S.: Isotopic fractionation between Fe(III) and Fe(II) in aqueous solutions, Earth Planet. Sc. Lett., 195, 141–153, https://doi.org/10.1016/S0012-821X(01)00581-7, 2002.
Johnson, K. S., Gordon, R. M., and Coale, K. H.: What controls dissolved iron concentrations in the world ocean?, Mar. Chem., 57, 137–161, https://doi.org/10.1016/S0304-4203(97)00043-1, 1997.
Krayenbuehl, P. A., Walczyk, T., Schoenberg, R., von Blanckenburg, F., and Schulthess, G.: Hereditary hemochromatosis is reflected in the iron isotope composition of blood, Blood, 105, 3812–3816, https://doi.org/10.1182/blood-2004-07-2807, 2005.
Kurisu, M., Sakata, K., Nishioka, J., Obata, H., Conway, T. M., Hunt, H. R., Sieber, M., Suzuki, K., Kashiwabara, T., Kubo, S., Takada, M., and Takahashi, Y.: Source and Fate of Atmospheric Iron Supplied to the Subarctic North Pacific Traced by Stable Iron Isotope Ratios, Geochim. Cosmochim. Ac., 378, 168–185, https://doi.org/10.1016/j.gca.2024.06.009, 2024.
Kwong, R. W. M. and Niyogi, S.: An in vitro examination of intestinal iron absorption in a freshwater teleost, rainbow trout (Oncorhynchus mykiss), J. Comp. Physiol. B, 178, 963–975, https://doi.org/10.1007/s00360-008-0279-3, 2008.
Maréchal, C. N., Télouk, P., and Albarède, F.: Precise analysis of copper and zinc isotopic compositions by plasma-source mass spectrometry, Chem. Geol., 156, 251–273, https://doi.org/10.1016/S0009-2541(98)00191-0, 1999.
Martin, J. H. and Gordon, R. M.: Northeast Pacific iron distributions in relation to phytoplankton productivity, Deep-Sea Res., 35, 177–196, https://doi.org/10.1016/0198-0149(88)90035-0, 1988.
Martin, J. H., Gordon, R. M., Fitzwater, S., and Broenkow, W. W.: VERTEX: phytoplankton/iron studies in the Gulf of Alaska, Deep-Sea Res., 36, 649–680, https://doi.org/10.1016/0198-0149(89)90144-1, 1989.
Nishioka, J. and Obata, H.: Dissolved iron distribution in the western and central subarctic Pacific: HNLC water formation and biogeochemical processes, Limnol. Oceanogr., 62, 2004–2022, https://doi.org/10.1002/lno.10548, 2017.
Pearson, A. B., Hückstädt, L. A., Kinsey, S. T., Schmitt, T. L., Robeck, T. R., Leger, J. St., Ponganis, P. J., and Tift, M. S.: Relationship between red blood cell lifespan and endogenous carbon monoxide in the common bottlenose dolphin and beluga, Am. J. Physiol. Regul. Integr. Comp. Physiol., 326, 134–146, https://doi.org/10.1152/ajpregu.00172.2023, 2024.
Pinedo-González, P., Hawco, N. J., and Bundy, R. M.: Anthropogenic Asian aerosols provide Fe to the North Pacific Ocean, P. Natl. Acad. Sci. USA, 117, 27862–27868, https://doi.org/10.1073/pnas.2010315117, 2020.
Plays, M., Müller, S., and Rodriguez, R.: Chemistry and biology of ferritin, Metallomics, 13, mfab021, https://doi.org/10.1093/mtomcs/mfab021, 2021.
Radic, A., Lacan, F., and Murray, J. W.: Iron isotopes in the seawater of the equatorial Pacific Ocean: New constraints for the oceanic iron cycle, Earth Planet. Sc. Lett., 306, 1–10, https://doi.org/10.1016/j.epsl.2011.03.015, 2011.
Robach, P., Cairo, G., Gelfi, C., Bernuzzi, F., Pilegaard, H., Viganò, A., Santambrogio, P., Cerretelli, P., Calbet, J. A. L., Phane Moutereau, S., and Lundby, C.: Strong iron demand during hypoxia-induced erythropoiesis is associated with down-regulation of iron-related proteins and myoglobin in human skeletal muscle, Blood, 109, 4724–4731, https://doi.org/10.1182/blood-2006-08-040006, 2007.
Robbins, E. and Pederson, T.: Iron: its intracellular localization and possible role in cell division, P. Natl. Acad. Sci. USA, 66, 1244–1251, https://doi.org/10.1073/pnas.66.4.1244, 1970.
Ryan, B. J., Foug, K. L., Gioscia-Ryan, R. A., Varshney, P., Ludzki, A. C., Ahn, C., Schleh, M. W., Gillen, J. B., Chenevert, T. L., and Horowitz, J. F.: Exercise training decreases whole-body and tissue iron storage in adults with obesity, Exp. Physiol., 106, 820–827, https://doi.org/10.1113/EP089272, 2021.
Sauzéat, L., Moreira, M., Holota, H., Beaudoin, C., and Volle, D. H.: Unveiling the hidden impact of long-term metal-rich volcanic pollution on male reproductive functions using isotope metallomics, Environ. Int., 198, 109388, https://doi.org/10.1016/j.envint.2025.109388, 2025.
Shemin, D. and Rittenberg, D.: The life span of the human red blood cell, J. Biol. Chem., 166, 627–636, 1946.
Shiraishi, T., Okamoto, K., Yoneda, M., Sakai, T., Ohshimo, S., Onoe, S., Yamaguchi, A., and Matsuyama, M.: Age validation, growth and annual reproductive cycle of chub mackerel Scomber japonicus off the waters of northern Kyushu and in the East China Sea, Fisheries Sci., 74, 947–954, https://doi.org/10.1111/j.1444-2906.2008.01612.x, 2008.
Sieber, M., Conway, T. M., de Souza, G. F., Hassler, C. S., Ellwood, M. J., and Vance, D.: Isotopic fingerprinting of biogeochemical processes and iron sources in the iron-limited surface Southern Ocean, Earth Planet. Sc. Lett., 567, 116967, https://doi.org/10.1016/j.epsl.2021.116967, 2021.
Slusarczyk, P. and Mleczko-Sanecka, K.: The multiple facets of iron recycling, Genes-Basel, 12, 1364, https://doi.org/10.3390/genes12091364, 2021.
Stenberg, A., Malinovsky, D., Rodushkin, I., Andrén, H., Pontér, C., Öhlander, B., and Baxter, D. C.: Separation of Fe from whole blood matrix for precise isotopic ratio measurements by MC-ICP-MS: A comparison of different approaches, J. Anal. Atom. Spectrom., 18, 23–28, https://doi.org/10.1039/b210482b, 2003.
van den Heuvel, E. G., Muys, T., Pellegrom, H., Bruyntjes, J. P., Van Dokkum, W., Spanhaak, S., and Schaafsma, G.: A new method to measure iron absorption from the enrichment of 57Fe and 58Fe in young erythroid cells, Clin. Chem., 44, 649–654, https://doi.org/10.1093/clinchem/44.3.649,1998.
van Heghe, L., Delanghe, J., Van Vlierberghe, H., and Vanhaecke, F.: The relationship between the iron isotopic composition of human whole blood and iron status parameters, Metallomics, 5, 1503–1509, https://doi.org/10.1039/c3mt00054k, 2013.
van Heghe, L., Deltombe, O., Delanghe, J., Depypere, H., and Vanhaecke, F.: The influence of menstrual blood loss and age on the isotopic composition of Cu, Fe and Zn in human whole blood, J. Anal. Atom. Spectrom., 29, 478–482, https://doi.org/10.1039/c3ja50269d, 2014.
von Blanckenburg, F., Noordmann, J., and Guelke-Stelling, M.: The iron stable isotope fingerprint of the human diet, J. Agr. Food Chem., 61, 11893–11899, https://doi.org/10.1021/jf402358n, 2013.
Walczyk, T. and von Blanckenburg, F.: Natural iron isotope variations in human blood, Science, 29, 2065–2066, https://doi.org/10.1126/science.1069389, 2002.
Walczyk, T. and von Blanckenburg, F.: Deciphering the iron isotope message of the human body, Int. J. Mass Spectrom., 242, 117–134, https://doi.org/10.1016/j.ijms.2004.12.028, 2005.
Welch, S. A., Beard, B. L., Johnson, C. M., and Braterman, P. S.: Kinetic and equilibrium Fe isotope fractionation between aqueous Fe(II) and Fe(III), Geochim. Cosmochim. Ac., 67, 4231–4250, https://doi.org/10.1016/S0016-7037(03)00266-7, 2003.
Wilke, M., Farges, F., Petit, P. E., Brown, G. E., and Martin, F.: Oxidation state and coordination of Fe in minerals: An Fe K-XANES spectroscopic study, Am. Mineral., 86, 714–730, https://doi.org/10.2138/am-2001-5-612, 2001.
Wilson, S. A., Green, E., Mathews, I. I., Benfatto, M., Hodgson, K. O., Hedman, B., and Sarangi, R.: X-ray absorption spectroscopic investigation of the electronic structure differences in solution and crystalline oxyhemoglobin, P. Natl. Acad. Sci. USA, 110, 16333–16338, https://doi.org/10.1073/pnas.1315734110, 2013.