the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 04 Jun 2026
| 04 Jun 2026
Temperature dependence of the contribution of soil water content to soil respiration in a monsoon influenced temperate deciduous forest
Dongmin Seo
Minyoung Lee
Jaeho Lee
Jaeseok Lee
Soil respiration (Rs) in forest soils is a key flux governing forest carbon balance and the global carbon cycle. Because this flux is expected to respond rapidly to climate warming, understanding the controls on Rs is essential for predicting changes in forest carbon balance induced by warming. In natural field conditions, soil temperature (Ts) and soil water content (SWC) often covary seasonally, which tends to limit our ability to isolate and quantify the independent contribution of SWC and to evaluate how its contribution varies with temperature. Although changes in the Rs response to temperature have been reported, few studies have quantitatively identified such patterns from field observations and examined whether the relative contribution of SWC becomes more evident across temperature conditions. Here, we used two years of continuous automated chamber measurements in a temperate deciduous forest to assess how the relative contribution of SWC varies with Ts by comparing models across temperature ranges, and to evaluate whether a breakpoint occurs in the Rs response to Ts and whether that breakpoint is observed near the temperature range where the contribution of SWC increases. At the annual scale, the explanatory power of SWC alone was limited, but the relationship between SWC and Rs was significant. In contrast, above 15 °C, the relationship between SWC and Rs strengthened consistently, indicating that the relative contribution of SWC became more evident under warm soil conditions. Piecewise regression of the relationship between Rs and Ts identified a breakpoint near 17 °C, and models including this breakpoint improved fit relative to an exponential model. These results suggest a possible difference in the relative importance of controls on Rs across temperature conditions. Therefore, projections of forest Rs may benefit from considering temperature dependent changes in the contribution of SWC, particularly near the temperature range where this contribution becomes more evident.
- Article
(1379 KB) - Full-text XML
-
Supplement
(493 KB) - BibTeX
- EndNote




Global warming is altering both air temperature and precipitation regimes, thereby affecting soil moisture conditions that influence carbon cycling in forest ecosystems. Forest ecosystems are known to play a central role in the global carbon cycle (IPCC, 2021). Forest soils store approximately 40 % of global soil organic carbon (Mayer et al., 2020), and forests have been reported to account for about 40 %–90 % of terrestrial soil respiration emissions (Rodtassana et al., 2021). These findings indicate that forests are important components of the carbon cycle because they function both as major carbon reservoirs and as sources of CO2 emissions (Win and Sato, 2024). In particular, soil respiration (Rs) is the CO2 flux from soil to the atmosphere, and the global annual magnitude is estimated to be about 70–100 Pg C yr−1 (Jian et al., 2018; Lei et al., 2021). Consequently, changes in Rs directly affect forest carbon balance and atmospheric CO2 variability, and variability in Rs strongly influences uncertainty in forest carbon balance estimates.
Under climate change, forest ecosystems are expected to be exposed not only to warming and rising atmospheric CO2 concentrations but also to shifts in precipitation seasonality and variability, yet substantial uncertainty remains regarding associated plant physiological responses and changes in forest soil microclimate (IPCC, 2021; Liu et al., 2025). Such changes may modify the relative contributions of soil temperature (Ts) and soil water content (SWC), thereby increasing uncertainty in predicting Rs responses (Liang et al., 2024). Nevertheless, quantitatively constraining the spatiotemporal variability of forest Rs and its controlling mechanisms remains a core challenge in carbon cycle research (Le Quéré et al., 2018).
Soil respiration is composed of root respiration from plant roots and microbial respiration associated with the decomposition of organic matter by soil microorganisms (Yan et al., 2025), and it responds strongly to seasonal biological activity and changes in meteorological conditions (Wang et al., 2021). Because Rs is observed as the combined flux of these two processes, their relative contributions may vary with phenological transitions and changes in soil water conditions, and the resulting sensitivities to Ts and SWC may therefore differ across seasons and temperature ranges (Yan et al., 2024). In general, increasing Ts enhances enzyme activity and metabolic rates, stimulating microbial decomposition and root physiological activity and thereby increasing Rs (Sáez-Sandino et al., 2023; Kengdo et al., 2023). Soil water content governs diffusion of oxygen and substrates in soil and can directly constrain or promote root water uptake and microbial physiological activity. Microbial activity is suppressed under dry conditions, whereas respiration can be limited by oxygen deficiency under wet conditions, a pattern that has long been recognized in studies of soil respiration responses to soil moisture and temperature (Cook and Orchard, 2008; Huang et al., 2023). Thus, Ts and SWC are key environmental factors controlling both root and microbial respiration.
To quantify these controls, previous studies have often simplified the influences of Ts and SWC using empirical functions. The Ts and Rs relationship has typically been represented using exponential or Q10 models, whereas the SWC and Rs relationship has commonly been described using polynomial functions that allow inhibition under both dry and wet extremes (Lloyd and Taylor, 1994; Davidson et al., 1998). Although previous studies have examined the combined effects of Ts and SWC on Rs, fewer field studies have explicitly quantified how the additional contribution of SWC changes across temperature ranges and whether this pattern is consistent with a breakpoint in the Rs response to Ts (Lai et al., 2012; Cui et al., 2020; Mao et al., 2024). In particular, at relatively low Ts, variations in Rs tend to be more strongly governed by temperature driven biochemical reaction rates, whereas at high Ts, changes in moisture supply and variability, or constraints associated with moisture deficit or excess, may jointly act to increase the sensitivity of Rs to changes in SWC (He et al., 2024). Even so, quantitative evidence from field observations remains limited regarding when the relative contribution of SWC begins to change systematically with Ts and whether a breakpoint in the Rs response to Ts occurs near the temperature range where this contribution becomes more evident.
It has been reported that the extent to which SWC explains variability in Rs depends strongly on time scale, phenological state, and temperature conditions (Kim et al., 2019; Podzikowski et al., 2025). Accordingly, analyses based on annual aggregation can mask short term and condition dependent effects of SWC by averaging across heterogeneous seasonal conditions. This averaging may also mask temperature dependent properties of SWC effects, limiting efforts to quantify the dependence of SWC contributions on Ts. Therefore, based on field observations, it remains uncertain how the effects of SWC change across temperature ranges (Liu et al., 2023; Bond-Lamberty et al., 2024). It is also important to evaluate whether a breakpoint in the Rs response to Ts occurs near the temperature range where the relative contribution of SWC becomes more evident, because this can help identify whether the two patterns occur over a similar temperature range and support interpretation of possible differences in the relative importance of controls on Rs.
Rather than changing monotonically with increasing Ts, Rs can exhibit nonlinearity when changes in moisture conditions and associated constraints modify the relative importance of controlling factors, leading to changes in response slopes. Breakpoint based interpretation may therefore help identify the temperature range where such changes in response become more evident, although breakpoint estimates should be interpreted in relation to vegetation type, climate, and site specific environmental conditions (Carey et al., 2016; Almagro et al., 2025; Li et al., 2025a).
Here, we used continuous chamber observations of Rs in a temperate deciduous forest to quantitatively assess whether the relative contribution of SWC varies systematically with Ts. By partitioning the analyses by temperature conditions to reduce the dilution that can arise from annual scale averaging, we aimed to provide field based evidence for the temperature dependence of SWC effects and to examine possible differences in the relative importance of controls on Rs across temperature conditions.
This study formulated and tested the following research questions.
-
At the annual scale, the explanatory power of SWC is often limited. We evaluate whether a statistically significant relationship between SWC and Rs exists despite this limitation.
-
If a relationship between SWC and Rs is observed, we examine whether its magnitude and statistical significance vary with Ts conditions.
-
Considering the seasonal covariation between Ts and SWC, we quantify how the relative contribution of SWC to Rs changes across temperature ranges after accounting for the temperature effect.
-
We assess whether the temperature range of the breakpoint in the Rs response to Ts is consistent with the temperature range where the relative contribution of SWC becomes more evident, and discuss what this convergence may suggest about possible differences in the relative importance of controlling factors governing Rs.
2.1 Study area
This study was conducted in a temperate deciduous forest in Gongju, Chungcheongnamdo, Republic of Korea (36°34′16′′ N, 127°00′34′′ E; 218 m above sea level). The site is influenced by the Asian monsoon climate, with mean annual precipitation of 1256.6 mm, of which more than 65 % occurs in summer from June–September. This pronounced precipitation seasonality results in warm, humid summers and cold, dry winters. Seasonal vegetation dynamics are distinct. Leaf development typically begins in April, and leaf senescence and abscission occur during October and November. Accordingly, leaf on and leaf off periods can be separated relatively clearly. The dominant tree species is konara oak (Quercus serrata), with oriental white oak (Quercus aliena), Japanese snowbell (Styrax japonicus), and Japanese cherry (Prunus serrulata) also present. During the study period, mean air temperature was 13.1 °C, and mean Ts measured at the site was 13.3 °C.
2.2 Measurement of environmental factors
Precipitation was continuously measured at 1.5 m above the ground surface within the forest using a tipping bucket rain gauge (S-RGB-M002, Onset, MA, USA). Ts was measured at 5 cm depth, and SWC was measured over the 0–15 cm depth using soil water content sensors (CS616, Campbell Scientific Inc., Logan, UT, USA). Precipitation, Ts, and SWC were recorded at 1 min intervals using a CR1000 datalogger (Campbell Scientific Inc.) and stored as 15 min means. The clocks of all devices were synchronized to a common reference time.
Periods flagged by quality control criteria, including rainfall, condensation, instrument maintenance, and sensor contamination, were excluded. For subsequent analyses, precipitation was aggregated to daily totals, and Ts and SWC were processed as daily means. Because rainfall can induce very brief increases in soil respiration immediately after rewetting, including Birch type pulses (Xu et al., 2004), daily aggregation was used to reduce the influence of these transient responses and to better evaluate the broader effect of SWC on Rs across temperature conditions.
2.3 Measurement of soil respiration
Soil respiration (Rs) was continuously measured from January 2022–December 2023 using five automated chambers operated in repeated open and closed cycles. The chambers were installed at least 1 m apart and were arranged to reflect the spatial heterogeneity of soil and surface environmental conditions within the plot, which included slope gradients ranging from 16 to 34°. To avoid spatial bias in chamber placement, the characteristics of each location were carefully assessed, and the five chambers were arranged in a roughly pentagonal configuration. In addition, the chambers were installed at least 50 cm away from tree stems to minimize the potential overestimation of root respiration associated with dense root distribution near the stems. Locations with substantial surface disturbance or conditions likely to reduce measurement stability were excluded. For each chamber, Ts and SWC sensors were installed together within 5 cm of the chamber so that both variables represented the same local environmental conditions. The Ts sensor was positioned adjacent to the SWC sensor, while avoiding any direct physical contact or interference that could affect SWC measurements. Each chamber operated on a 30 min cycle consisting of 25 min open and 5 min closed phases. During the closed period, chamber air was circulated through an infrared gas analyzer (IRGA; LI-840, LI-COR, Lincoln, NE, USA) using a pump. CO2 concentration was recorded at approximately 30 s intervals, and Rs was calculated from the rate of increase in CO2 concentration during the closed period. The IRGA inlet and outlet flow rates were regulated to 1.0 L min−1 using a flow meter. Chamber collars were inserted 10 cm into the soil and fixed in place, and measurements related to Rs were logged at 1 min intervals using a CR1000 datalogger. Aboveground vegetation inside the chambers was periodically removed to prevent CO2 interference from photosynthesis, while the litter layer was maintained to minimize disturbance to the soil surface. As with the Ts and SWC data, outliers and erroneous records were removed, and 30 min mean values were calculated. These were then used to derive daily mean soil respiration values (Eq. 1):
Where Rs is soil respiration (), is the rate of change in CO2 mass concentration in the chamber headspace (), V is chamber volume (m3), and A is chamber base area (m2).
2.4 Model fitting and evaluation
To quantify the combined effects of Ts and SWC on Rs, we applied nonlinear regression models. First, an exponential model describing the baseline temperature response was used as the Ts-only model (Eq. 2).
Ts-only model (Eq. 2):
To incorporate the effect of SWC and its temperature dependent influence, we defined an extended model in which the exponential temperature term was multiplied by a quadratic function of SWC, referred to as the Ts plus SWC model (Eq. 3).
Ts+SWC model (Eq. 3):
Model parameters , and e were estimated by fitting the model to the observations. For the full daily dataset, the Ts-only model yielded an R2 of 0.782, an RMSE of 274.94, and an AIC of 183 047, whereas the Ts+SWC model yielded an R2 of 0.846, an RMSE of 230.77, and an AIC of 177 346.
2.5 Statistical analysis
All analyses were conducted using SAS software version 9.4 (SAS Institute Inc., Cary, NC, USA). Equations (2) and (3) were fitted to the full daily dataset. We used fixed 5 °C Ts bins rather than a continuous sliding window to provide discrete and interpretable temperature ranges while maintaining sufficient observations within each range for model comparison. The daily data were then grouped into 5 °C Ts intervals for each year to further examine how the additional contribution of SWC changed across temperature conditions. Within each temperature range, the Ts-only model and the Ts+SWC model were compared using Adj. R2 and AIC. The contribution of SWC was quantified as the difference in Adj. R2 between the two models, and the degree of model improvement was quantified as the difference in AIC. We also evaluated the significance of the SWC term within each temperature range.
2.6 Breakpoint analysis of the soil respiration response to soil temperature
To identify a breakpoint in the temperature response of Rs, we applied segmented regression (Muggeo, 2003). Segmented regression estimates a breakpoint by fitting two or more linear segments with different slopes along a continuous predictor, here Ts. We first fitted an initial linear regression with Ts and then used iterative fitting to estimate the breakpoint and segment specific slopes. Analyses were performed in R (version 4.1.1) using the segmented package. The number of breakpoints was determined by comparing AIC among a model with no breakpoint and candidate models with one or more breakpoints (Table S1 in the Supplement). Fit between the segmented regression model and the exponential model was compared using ΔAIC, and the significance of the fit difference between models was evaluated using a bootstrap based test. The estimated breakpoint was interpreted as a reference temperature indicating a structural change in the Rs response to Ts, rather than as a direct threshold in SWC sensitivity. Temperature dependent changes in the relative contribution of SWC were evaluated separately using ΔAdj.R2 and ΔAIC from the Ts bin model comparisons. In addition, to visually examine whether unexplained variation in Rs became more dispersed across temperature conditions after removing the Ts effect, residuals were calculated as observed Rs minus Rs predicted by the Ts-only model and plotted against Ts with SWC indicated by color (Fig. S2 in the Supplement).
3.1 Environmental conditions and temporal variation in soil respiration
Mean annual Ts was 12.5 °C in 2022 and 13.5 °C in 2023, a difference of approximately 1 °C, and the difference between years was not statistically significant (p=0.08; Fig. 1a). Ts reached its maximum in July and August and its minimum in January and February, showing a typical seasonal pattern.
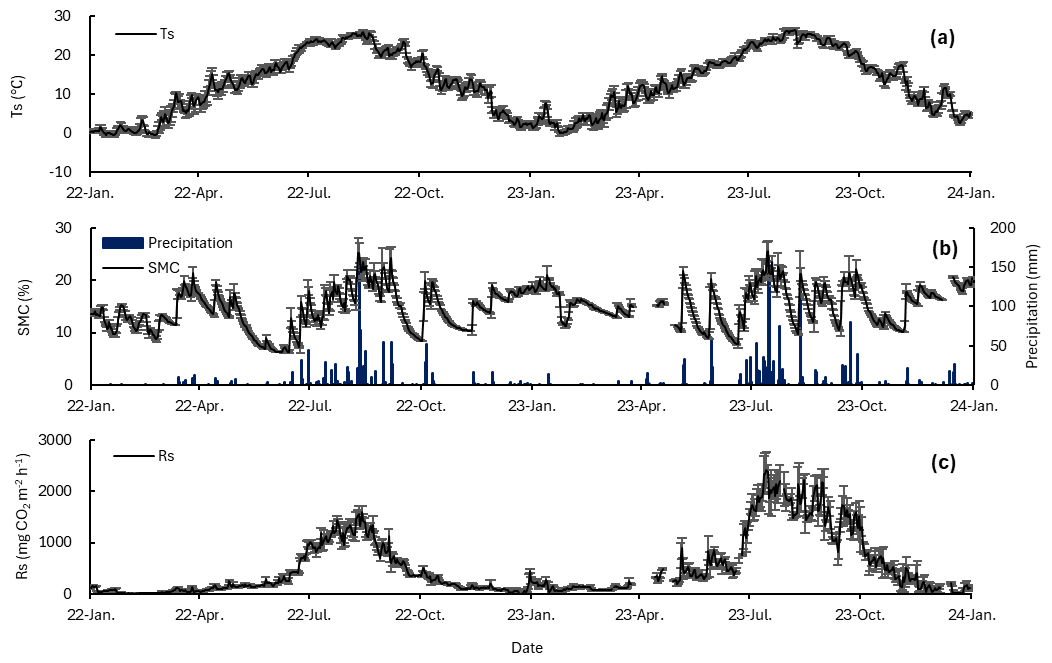
Figure 1Temporal variation in (a) Ts, (b) SWC and precipitation, and (c) Rs during the 2022–2023 observation period. Data gaps in SWC and Rs occurred from 26 March–13 April 2023 and from 21–29 April 2023 due to instrument malfunction. Error bars represent ±1 standard error (SE).
Annual precipitation increased from 1018.0 mm in 2022–1495.2 mm in 2023, an increase of 46.9 %, and the number of precipitation days increased from 90–100. Monthly precipitation peaked in August 2022 (362.3 mm) and July 2023 (504.0 mm), and precipitation was also high in September 2023 (204.3 mm).
SWC ranged from 6.2 %–25.3 % in 2022 and from 7.7 %–25.5 % in 2023, with mean annual values of 14.1 % in 2022 % and 15.6 % in 2023 (Fig. 1b). In 2022, monthly mean SWC was 9.6 % in May and 9.2 % in June, remaining below 10 %, and then increased after July to 20.1 % in August. In 2023, monthly mean SWC exceeded 12 % throughout the year and reached 20.3 % in July and 17.2 % in September. Additionally, short term increases in Rs were observed following increases in SWC after rainfall during the study period. Specifically, Rs tended to increase on the day of rainfall and on the following day, and a similar increasing pattern was also observed at 0, 1, and 2 h after rainfall at the hourly scale.
Mean annual Rs was 351.4 in 2022 and 701.9 in 2023 (Fig. 1c). Rs showed clear seasonality consistent with the seasonal variation in Ts, and cumulative Rs during July to September accounted for 68 % of annual cumulative Rs in 2022 % and 64 % in 2023.
3.2 Variability in soil respiration in relation to soil water content as a single factor
To examine the relationship between SWC and Rs, we first fitted a single variable regression with SWC as the sole predictor. In both years, Rs showed a consistent tendency to increase when SWC increased above a certain level. Based on the quadratic regression, the SWC value corresponding to the minimum of the quadratic function was estimated at approximately 10.8 % in 2022 % and 13.1 % in 2023 (Fig. 2). However, Rs was widely distributed even within the same SWC range, indicating substantial variability. When considering SWC alone, the coefficient of determination for explaining variability in Rs was low, with R2=0.29 in 2022 and R2=0.21 in 2023, but the relationship between SWC and Rs was statistically significant in both years (p<0.0001).
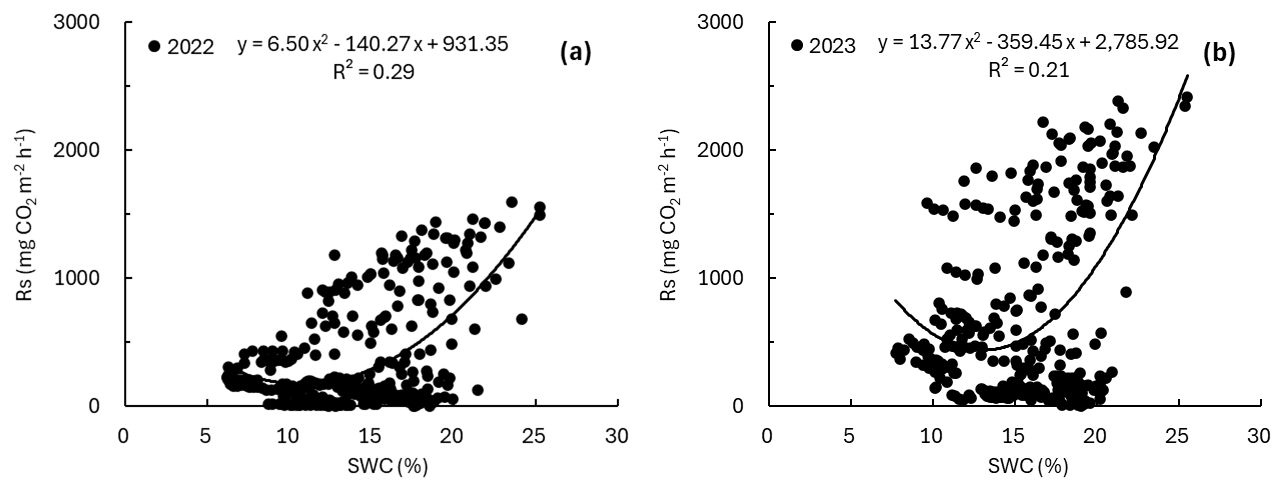
3.3 Soil respiration responses to soil water content across soil temperature ranges
Rs varied widely even within the same SWC range, and the single factor analysis did not show a consistent pattern between Rs and SWC. We therefore binned Ts and examined Rs responses to SWC under comparable temperature conditions (Fig. 3). The numbers of daily observations in each Ts bin were as follows: in 2022, n=20 for <0 °C, n=78 for 0–5 °C, n=39 for 5–10 °C, n=86 for 10–15 °C, n=56 for 15–20 °C, n=82 for 20–25 °C, and n=24 for 25–30 °C; in 2023, n=74for 0–5 °C, n=46 for 5–10 °C, n=44 for 10–15 °C, n=67 for 15–20 °C, n=72 for 20–25 °C, and n=31 for 25–30 °C. For Ts bins above 15 °C, R2 ranged from 0.39–0.76 in 2022 and from 0.48–0.61 in 2023, and R2 generally increased with higher temperature bins. In both years, the relationship between SWC and Rs was statistically significant for Ts bins above 15 °C (p<0.0001). In 2022, within the 20–25 °C Ts bin, Rs increased with increasing SWC, but the rate of increase declined at higher SWC, indicating a reduced slope at high SWC (Fig. 3). In contrast, for Ts bins below 15 °C, R2 was low, ranging from 0.07–0.13 in 2022 and from 0.00–0.06 in 2023, and the relationship between SWC and Rs was not statistically significant in some bins.
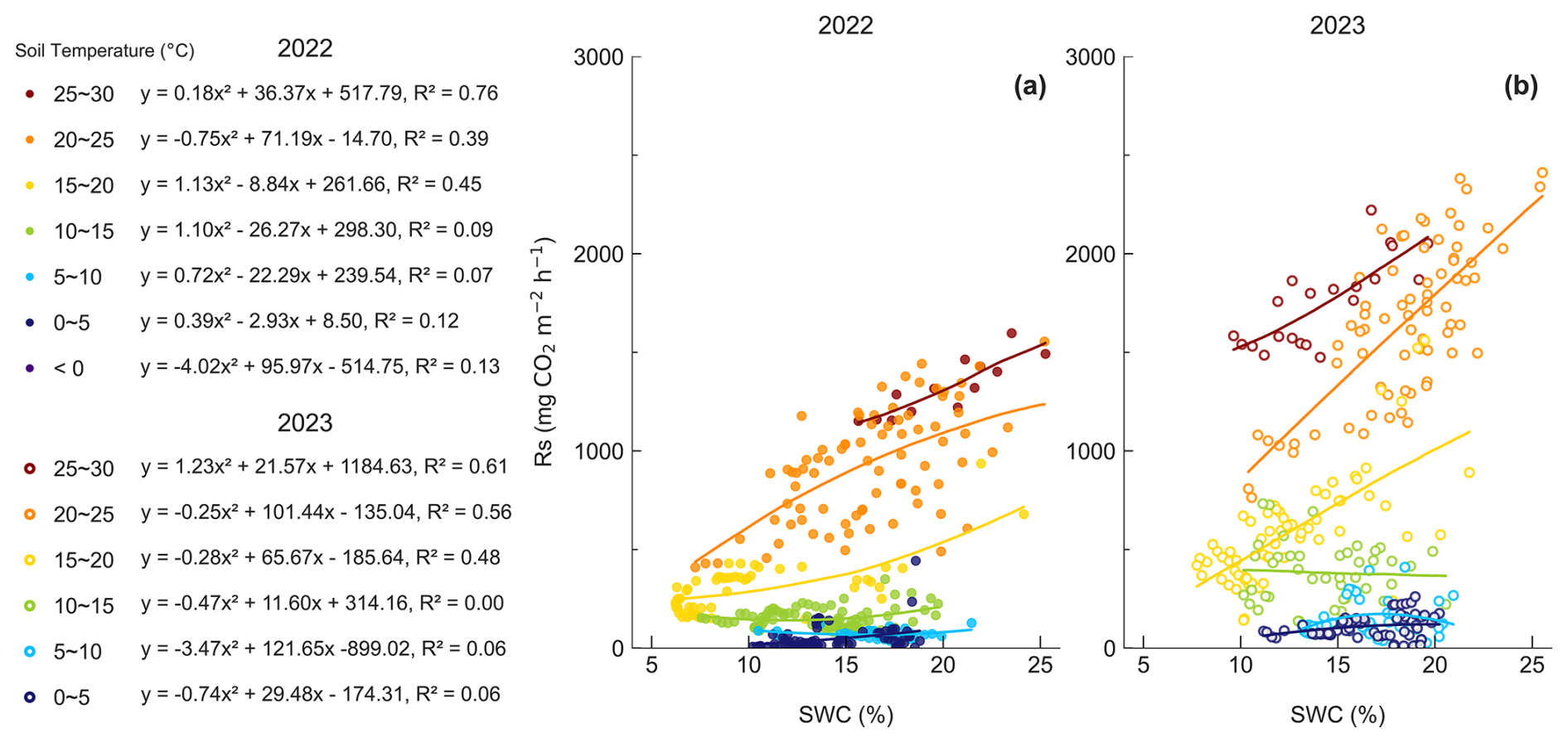
Figure 3Relationship between SWC and Rs across Ts bins in (a) 2022 and (b) 2023. In 2023, no days had Ts below 0 °C, so the Ts bins below 0 °C were excluded from the analysis.
3.4 Changes in the contribution of soil water content across soil temperature ranges
To directly test whether the contribution of SWC to Rs changed across temperature conditions after accounting for Ts, we compared the Ts-only model and the Ts+SWC model within each temperature bin and quantified the difference as the SWC contribution (ΔAdj.R2) (Fig. 4). Over the full period, the SWC contribution was 0.03 in 2022 and 0.09 in 2023. However, the SWC contribution varied substantially among temperature bins. Below 15 °C, the SWC contribution was generally close to zero, ranging from 0.00–0.07 in 2022 and from −0.04–0.05 in 2023, whereas above 15 °C it increased markedly, ranging from 0.21–0.43 in 2022 and from 0.44–0.62 in 2023. Consistent with this pattern, indicated improved fit of the Ts+SWC model above 15 °C, while the improvement was relatively small below 15 °C (Fig. S1 in the Supplement).
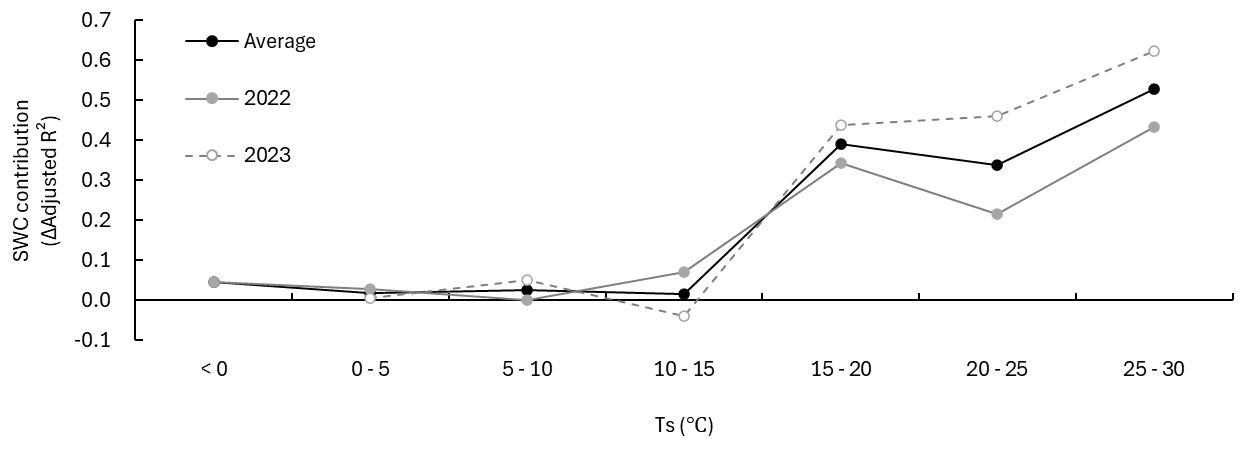
Figure 4SWC contribution (ΔAdj.R2) by Ts bin. The SWC contribution was calculated as the difference in adjusted R2 between the Ts-only model and the Ts+SWC model (ΔAdj.R2). The gray solid line indicates 2022, the gray dashed line indicates 2023, and the black solid line indicates the mean across both years.
3.5 Breakpoint in the soil respiration response to soil temperature
The contribution of SWC differed across temperature conditions and became more evident at higher Ts. We then applied segmented regression to the relationship between Rs and Ts to examine whether a breakpoint occurred near the temperature range where this pattern emerged (Fig. 5). In both years, a breakpoint was detected near 17 °C, with breakpoint estimates of 16.94±0.44 °C in 2022 and 16.77±0.75 °C in 2023, where ± indicates the 95 % confidence interval. To further examine the temperature response across the breakpoint, we analyzed the Ts range, slope, R2, p value, and mean SWC for the segments below and above the breakpoint (Table S2 in the Supplement). The number of breakpoints was optimized by comparing AIC among candidate models, and the segmented regression model with one breakpoint was selected as the best model (Table S1). The segmented regression model also showed improved fit relative to the exponential model, with lower AIC, and this improvement was significant based on a bootstrap test (2022 ΔAIC=20.4, p<0.0001; 2023 ΔAIC=112.4, p<0.0001). To provide a more direct visual assessment of the variation not explained by Ts alone, we additionally examined the residuals of the Ts-only model across the Ts range (Fig. S2). In both years, residuals appeared relatively more constrained at lower Ts, but became more widely dispersed in the warmer temperature range, where variation in SWC was also more evident. However, this wider residual dispersion does not by itself demonstrate that SWC explains the additional variation, because it may also reflect unmeasured seasonal covariates such as phenological changes in root activity or fine root biomass.
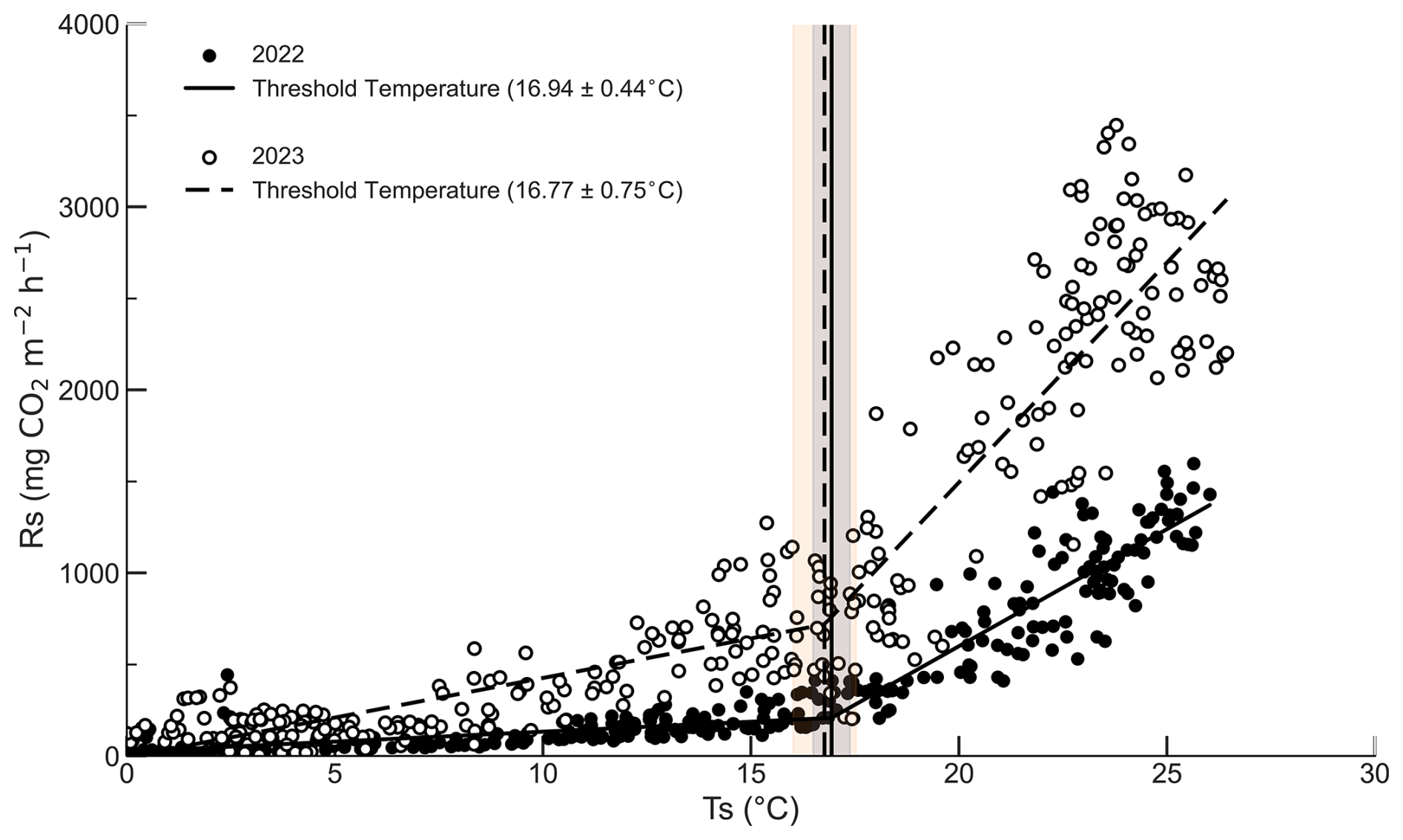
Figure 5Soil temperature breakpoint estimated by applying segmented regression to the relationship between Rs and Ts. Vertical lines indicate the estimated breakpoint temperatures (16.94 °C in 2022 and 16.77 °C in 2023), and shaded areas indicate the 95 % confidence intervals for each breakpoint.
Our results indicate that the effect of SWC on Rs depends on soil temperature, with SWC exerting a stronger influence under warmer conditions. This pattern suggests that the relative importance of SWC is not constant across temperature conditions and that the relative importance of controls on Rs may differ between cooler and warmer conditions. To examine temperature dependent changes in the contribution of SWC and whether a breakpoint in the Rs response to Ts occurs near the temperature range where this contribution becomes more evident, we quantified SWC effects within discrete temperature conditions and evaluated the presence of a breakpoint. However, this pattern should be interpreted cautiously because the breakpoint in the Rs response to Ts does not directly represent a threshold in SWC sensitivity.
4.1 Relationship between soil water content as a single factor and soil respiration
Although SWC was significantly associated with Rs, variation in SWC alone explained only a limited portion of Rs variability. The wide dispersion of Rs within similar SWC ranges indicates that the influence of SWC is strongly conditioned by other environmental drivers. This interpretation is consistent with previous studies showing that temporal variation in Rs is difficult to explain using SWC alone (Liang et al., 2010; Yu et al., 2021). Soil respiration is influenced by environmental drivers beyond SWC, and key factors such as Ts can change concurrently with seasonal environmental conditions (Rodtassana et al., 2021). Accordingly, the response of Rs to SWC should be interpreted in relation to the combined effects of temperature and SWC rather than as an independent effect of SWC. The close seasonal correspondence between Rs and Ts in our data further supports temperature as a primary background control, which may obscure a simple direct relationship between SWC and Rs (Matsumoto et al., 2023).
4.2 Changes in the effect of soil water content under different temperature conditions
These findings indicate that the influence of SWC on Rs varies across temperature conditions rather than being expressed uniformly over the full Ts range. This temperature dependence suggests that the contribution of SWC to Rs variability becomes more evident under warm conditions, whereas its contribution is weaker or less consistent under cooler conditions.
Under sufficiently warm conditions, microbial and root activity increase (Birgander et al., 2013; González-García et al., 2023), and Rs may respond more sensitively to changes in SWC. At our forest site, rainfall events during warm periods likely generated short term fluctuations in SWC, and these fluctuations may have contributed substantially to temporal variation in Rs. One possible explanation is that dry soil conditions restrict solute transport and substrate diffusion, while severe moisture limitation can suppress microbial and root activity or induce dormancy (Huang et al., 2005; Wang et al., 2014). At the wetter end of the SWC range, Rs may also become constrained if oxygen diffusion declines and gas diffusion resistance increases (Yan et al., 2018).
In addition, the shift in Rs response around 15 °C cannot be fully explained by Ts or SWC alone and may also reflect seasonal biological changes, including increases in fine root biomass and activity and enhanced microbial activity during warmer phenophases (Schindlbacher et al., 2015; Heinzle et al., 2023).
We also observed that in some warm Ts bins, the Rs response weakened at higher SWC (Fig. 3), which is consistent with reports that increasing SWC does not always increase Rs linearly and that physical constraints at high SWC, such as limited oxygen diffusion, can constrain the response (Zhu et al., 2020). Taken together, these patterns suggest that the response of Rs to SWC under warm conditions is nonlinear, reflecting both the release of moisture limitation under dry conditions and the emergence of diffusion related constraints at higher SWC.
In contrast, under low Ts conditions, microbial and root activity generally decline (Chen et al., 2021; Schnecker et al., 2023), and the magnitude of the Rs response to changes in SWC may therefore remain limited even under similar changes in SWC. Consistent with this interpretation, the influence of SWC on Rs was weak and less consistent under Ts below 15 °C. In addition, during cold periods, lower rainfall frequency and weaker evapotranspiration effects can reduce the amplitude of SWC variability. This limited variability may also make the estimated effect of SWC less stable and more difficult to detect statistically in regression analyses.
4.3 Contribution of soil water content to soil respiration after accounting for the temperature effect
Under field conditions, Ts and SWC often change together with seasonal variability (Li et al., 2022), and Rs is generally higher during warmer periods (Zhang et al., 2023). Accordingly, the stronger SWC and Rs relationship observed during warm periods must be interpreted carefully to determine whether it reflects an independent SWC effect or arises partly from concurrent increases in Ts and shared seasonal patterns.
When the effect of temperature was constrained within comparable Ts ranges, the influence of SWC on Rs became clearer under warm conditions. This comparison further indicates that SWC can provide explanatory power beyond temperature alone, but primarily under sufficiently warm conditions.
The stronger contribution of SWC under Ts above 15 °C suggests that SWC explains additional variation in Rs under warm conditions beyond that captured by temperature alone, although seasonal covariance between Ts and SWC cannot be fully excluded. By contrast, under cooler conditions, the independent role of SWC appears limited. This interpretation is consistent with reduced microbial and root activity under low Ts, which may suppress the sensitivity of Rs to SWC (Huang et al., 2005). Reduced SWC variability during cold periods may also weaken the extent to which SWC related changes are expressed in Rs variability. Overall, these findings support the view that the contribution of SWC to Rs is temperature dependent and becomes more pronounced only under sufficiently warm conditions.
4.4 Effect of soil water content on soil respiration below and above the breakpoint
Previous studies that quantified temperature breakpoints in Rs have reported breakpoints near 18 °C in forests, and our estimates fall within a similar range (Almagro et al., 2025). In our study, the breakpoint was consistently detected near 17 °C in both years (Fig. 5). The relative contribution of SWC to Rs variability differed between cooler and warmer temperature conditions, with the influence of SWC becoming more evident under warmer conditions. A breakpoint in the Rs response to Ts was also detected near the temperature range where this pattern emerged. These results suggest a possible shift in the relative importance of the controls on Rs between cooler and warmer conditions (Johnston et al., 2021). This breakpoint may reflect the combined effects of enhanced plant and microbial activity under warmer conditions and greater variation in SWC during warm periods, which together increase the relative influence of SWC on Rs (Kaisermann et al., 2017; Wang et al., 2022; Li et al., 2025b). The location of such a breakpoint may vary with regional climate, vegetation characteristics, and soil environmental conditions, including soil moisture regimes (Lellei-Kovács et al., 2016; Almagro et al., 2025). However, because this study was conducted at a single site, the extent to which the identified breakpoint near 17 °C can be generalized remains limited. In addition, because Rs was not partitioned into autotrophic and heterotrophic components, the breakpoint cannot be attributed specifically to moisture related constraints on microbial activity rather than to phenological transitions in root respiration. An exploratory comparison between the foliage season and non foliage season showed that the contribution of SWC tended to be more evident during the foliage season and limited during the non foliage season. However, these results are not shown because root activity was not directly measured and Rs was not partitioned into autotrophic and heterotrophic components.
4.5 Implications and considerations
This study suggests that Rs variability is not governed consistently by a single factor and that the relative importance of key controls may vary depending on Ts conditions. In particular, the greater relative contribution of SWC under warm conditions indicates that understanding and predicting Rs variability during warm periods requires consideration of not only Ts but also the seasonal water regime, including variation in SWC and water supply. This implies that under conditions expected to involve longer warm periods or increased precipitation variability, Rs responses may not be adequately described by a temperature sensitivity metric such as Q10 alone. Accordingly, carbon cycle modeling and predictions based on field observations should explicitly represent the role of SWC terms under warm conditions or adopt a structure that allows the sensitivity of Rs to SWC to differ with Ts, such as interaction terms or models fitted within temperature bins. However, daily resolution may not fully capture short term moisture related Rs dynamics occurring near the identified breakpoint, and future studies using higher temporal resolution and event based approaches could better resolve rapid Rs responses to changes in SWC within this temperature range.
At the same time, because breakpoint location can shift with regional climate, vegetation, and soil environmental conditions, caution is required when interpreting our results beyond the hydroclimatic setting of this study. In particular, in temperate deciduous forests, monsoon climates tend to maintain relatively high soil water availability during the warm season because precipitation is concentrated in this period, whereas non monsoon climates tend to experience intensified seasonal drying with increasing temperature (Chae, 2011; Prigoliti et al., 2023). In this context, our findings should be interpreted within the hydroclimatic setting of monsoon influenced temperate forests where soil water availability remains relatively high during the warm season, and the same pattern may not occur in non-monsoon temperate forests where water availability is seasonally limited. In addition, because Ts and SWC can covary seasonally under field conditions, the breakpoint may reflect combined seasonal drivers rather than a purely physiological moisture threshold. Therefore, interpretation should emphasize the pattern of change across the breakpoint rather than the specific temperature value itself. Clarifying the processes underlying breakpoint formation will require further studies incorporating phenological indicators, microbial activity indices, and component partitioning.
This study suggests that the relative contribution of SWC to Rs variability changes across temperature conditions. Although the explanatory power of the SWC only regression was limited, we found a consistent pattern in which the relationship between SWC and Rs became stronger above a certain Ts. In addition, a breakpoint in the Rs response to Ts was identified near the temperature range where the relative contribution of SWC became more evident, with estimates near 17 °C in both years. Taken together, these findings suggest that the relative importance of controls on Rs may differ between cooler and warmer soil conditions, with the contribution of SWC becoming greater under warm soil conditions and remaining limited or less evident under cool soil conditions.
The segmented regression analyses were performed in R using standard packages. The custom scripts used for data processing and analysis are available from the corresponding author upon reasonable request. These scripts are not publicly archived because they were developed for the data processing and analysis workflow of an institutionally managed long term monitoring program.
The data supporting the findings of this study are available from the corresponding author upon reasonable request. The data are not publicly archived because they were collected as part of an ongoing long term monitoring project at a protected forest research site and are subject to institutional restrictions on public data sharing.
The supplement related to this article is available online at https://doi.org/10.5194/bg-23-3723-2026-supplement.
DS designed the study, carried out field measurements, performed data curation and formal analysis, and wrote the first draft of the manuscript. ML contributed to material preparation and field measurements. JaehL was responsible for instrument setup and installation. JaesL supervised the study and contributed to data interpretation and manuscript revision. All authors commented on previous versions of the manuscript and approved the final version.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We thank all those who helped with site maintenance, soil respiration measurements, and sample analysis.
This research has been supported by the National Research Foundation of Korea (grant no. 2020R1I1A2073000).
This paper was edited by Bertrand Guenet and reviewed by Timo Plaçais and one anonymous referee.
Almagro, M., Rey, A., Inclán, R. M., Barba, J., Vargas, R., Carrara, A., Grünzweig, J. M., Sternberg, M., Talmon, Y., McCulley, R. L., Marañón-Jiménez, S., Serrano-Ortiz, P., Martínez-López, J., Estruch, C., Guidolotti, G., Chang, C. T., Llovet, J., Lo Cascio, M., Perez-Quezada, J. F., Correia, A. C., Banza, J., Caldeira, M. C., Nogueira, C., Bugalho, M. N., Carbone, M. S., Martínez-Mena, M., Mereu, S., and Curiel Yuste, J.: Vegetation type and climate determine temperature thresholds of soil respiration across drylands, Soil Biol. Biochem., 211, 109984, https://doi.org/10.1016/j.soilbio.2025.109984, 2025.
Birgander, J., Reischke, S., Jones, D. L., and Rousk, J.: Temperature adaptation of bacterial growth and 14C-glucose mineralisation in a laboratory study, Soil Biol. Biochem., 65, 294–303, https://doi.org/10.1016/j.soilbio.2013.06.006, 2013.
Bond-Lamberty, B., Ballantyne, A., Berryman, E., Fluet-Chouinard, E., Jian, J., Morris, K. A., Rey, A., and Vargas, R.: Twenty years of progress, challenges, and opportunities in measuring and understanding soil respiration, J. Geophys. Res.-Biogeo., 129, e2023JG007637, https://doi.org/10.1029/2023JG007637, 2024.
Carey, J. C., Tang, J., Templer, P. H., Kroeger, K. D., Crowther, T. W., Burton, A. J., Dukes, J. S., Emmett, B., Frey, S. D., Heskel, M. A., Jiang, L., Machmuller, M. B., Mohan, J., Panetta, A. M., Reich, P. B., Reinsch, S., Wang, X., Allison, S. D., Bamminger, C., Bridgham, S., Collins, S. L., de Dato, G., Eddy, W. C., Enquist, B. J., Estiarte, M., Harte, J., Henderson, A., Johnson, B. R., Larsen, K. S., Luo, Y., Marhan, S., Melillo, J. M., Peñuelas, J., Pfeifer-Meister, L., Poll, C., Rastetter, E., Reinmann, A. B., Reynolds, L. L., Schmidt, I. K., Shaver, G. R., Strong, A. L., Suseela, V., and Tietema, A.: Temperature response of soil respiration largely unaltered with experimental warming, P. Natl. Acad. Sci. USA, 113, 13797–13802, https://doi.org/10.1073/pnas.1605365113, 2016.
Chae, N.: Annual variation of soil respiration and precipitation in a temperate forest (Quercus serrata and Carpinus laxiflora) under East Asian monsoon climate, J. Plant Biol., 54, 101–111, https://doi.org/10.1007/s12374-011-9148-9, 2011.
Chen, T., Lin, C., Song, T., Guo, R., Cai, Y., Chen, W., Xiong, D., Jiang, Q., and Chen, G.: Does root respiration of subtropical Chinese fir seedlings acclimate to seasonal temperature variation or experimental soil warming?, Agr. Forest. Meteorol., 308–309, 108612, https://doi.org/10.1016/j.agrformet.2021.108612, 2021.
Cook, F. J. and Orchard, V. A.: Relationships between soil respiration and soil moisture, Soil Biol. Biochem., 40, 1013–1018, https://doi.org/10.1016/j.soilbio.2007.12.012, 2008.
Cui, Y. B., Feng, J. G., Liao, L. G., Yu, R., Zhang, X., Liu, Y. H., Yang, L. Y., Zhao, J. F., and Tan, Z. H.: Controls of temporal variations on soil respiration in a tropical lowland rainforest in Hainan Island, China, Trop. Conserv. Sci., 13, 1940082920914902, https://doi.org/10.1177/1940082920914902, 2020.
Davidson, E. A., Belk, E., and Boone, R. D.: Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest, Glob. Change Biol., 4, 217–227, https://doi.org/10.1046/j.1365-2486.1998.00128.x, 1998.
González-García, M. P., Conesa, C. M., Lozano-Enguita, A., Baca-González, V., Simancas, B., Navarro-Neila, S., Sánchez-Bermúdez, M., Salas-González, I., Caro, E., Castrillo, G., and del Pozo, J. C.: Temperature changes in the root ecosystem affect plant functionality, Plant Commun., 4, 100514, https://doi.org/10.1016/j.xplc.2022.100514, 2023.
He, H., Zha, T., and Tan, J.: Soil-moisture-dependent temperature sensitivity of soil respiration in a poplar plantation in northern China, Forests, 15, 1466, https://doi.org/10.3390/f15081466, 2024.
Heinzle, J., Liu, X., Tian, Y., Kwatcho Kengdo, S., Heinze, B., Nirschi, A., Borken, W., Inselsbacher, E., Wanek, W., and Schindlbacher, A.: Increase in fine root biomass enhances root exudation by long term soil warming in a temperate forest, Front. For. Glob. Change, 6, 1152142, https://doi.org/10.3389/ffgc.2023.1152142, 2023.
Huang, X., Lakso, A. N., and Eissenstat, D. M.: Interactive effects of soil temperature and moisture on Concord grape root respiration, J. Exp. Bot., 56, 2651–2660, https://doi.org/10.1093/jxb/eri258, 2005.
Huang, Z., Liu, Y., Huang, P., Li, Z., and Zhang, X.: A new concept for modelling the moisture dependence of heterotrophic soil respiration, Soil Biol. Biochem., 185, 109147, https://doi.org/10.1016/j.soilbio.2023.109147, 2023.
Intergovernmental Panel on Climate Change (IPCC): Climate change 2021: the physical science basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge University Press, Cambridge, UK and New York, NY, USA, https://doi.org/10.1017/9781009157896, 2021.
Jian, J., Steele, M. K., Thomas, R. Q., Day, S. D., and Hodges, S. C.: Constraining estimates of global soil respiration by quantifying sources of variability, Glob. Change Biol., 24, 4143–4159, https://doi.org/10.1111/gcb.14301, 2018.
Johnston, A. S. A., Meade, A., Ardö, J., Arriga, N., Black, A., Blanken, P. D., Bonal, D., Brümmer, C., Cescatti, A., Dušek, J., Graf, A., Gioli, B., Goded, I., Gough, C. M., Ikawa, H., Jassal, R., Kobayashi, H., Magliulo, V., Manca, G., Montagnani, L., Moyano, F. E., Olesen, J. E., Sachs, T., Shao, C., Tagesson, T., Wohlfahrt, G., Wolf, S., Woodgate, W., Varlagin, A., and Venditti, C.: Temperature thresholds of ecosystem respiration at a global scale, Nat. Ecol. Evol., 5, 487–494, https://doi.org/10.1038/s41559-021-01398-z, 2021.
Kaisermann, A., de Vries, F. T., Griffiths, R. I., and Bardgett, R. D.: Legacy effects of drought on plant-soil feedbacks and plant-plant interactions, New Phytol., 215, 1413–1424, https://doi.org/10.1111/nph.14661, 2017.
Kengdo, S. K., Ahrens, B., Tian, Y., Heinzle, J., Wanek, W., Schindlbacher, A., and Borken, W.: Increase in carbon input by enhanced fine root turnover in a long-term warmed forest soil, Sci. Total Environ., 855, 158800, https://doi.org/10.1016/j.scitotenv.2022.158800, 2023.
Kim, D., Lee, M., and Seo, E.: Improvement of soil respiration parameterization in a dynamic global vegetation model and its impact on the simulation of terrestrial carbon fluxes, J. Climate, 32, 127–143, https://doi.org/10.1175/JCLI-D-18-0018.1, 2019.
Lai, L., Zhao, X., Jiang, L., Wang, Y., Luo, L., Zheng, Y., Chen, X., and Rimmington, G. M.: Soil respiration in different agricultural and natural ecosystems in an arid region, PLoS One, 7, e48011, https://doi.org/10.1371/journal.pone.0048011, 2012.
Lei, J., Guo, X., Zeng, Y., Zhou, J., Gao, Q., and Yang, Y.: Temporal changes in global soil respiration since 1987, Nat. Commun., 12, 403, https://doi.org/10.1038/s41467-020-20616-z, 2021.
Lellei-Kovács, E., Botta-Dukát, Z., de Dato, G., Estiarte, M., Guidolotti, G., Kopittke, G. R., Kovács-Láng, E., Kröel-Dulay, G., Larsen, K. S., Peñuelas, J., Smith, A. R., Sowerby, A., Tietema, A., and Schmidt, I. K.: Temperature dependence of soil respiration modulated by thresholds in soil water availability across European shrubland ecosystems, Ecosystems, 19, 1460–1477, https://doi.org/10.1007/s10021-016-0016-9, 2016.
Le Quéré, C., Andrew, R. M., Friedlingstein, P., Sitch, S., Pongratz, J., Manning, A. C., Korsbakken, J. I., Peters, G. P., Canadell, J. G., Jackson, R. B., Boden, T. A., Tans, P. P., Andrews, O. D., Arora, V. K., Bakker, D. C. E., Barbero, L., Becker, M., Betts, R. A., Bopp, L., Chevallier, F., Chini, L. P., Ciais, P., Cosca, C. E., Cross, J., Currie, K., Gasser, T., Harris, I., Hauck, J., Haverd, V., Houghton, R. A., Hunt, C. W., Hurtt, G., Ilyina, T., Jain, A. K., Kato, E., Kautz, M., Keeling, R. F., Klein Goldewijk, K., Körtzinger, A., Landschützer, P., Lefèvre, N., Lenton, A., Lienert, S., Lima, I., Lombardozzi, D., Metzl, N., Millero, F., Monteiro, P. M. S., Munro, D. R., Nabel, J. E. M. S., Nakaoka, S., Nojiri, Y., Padin, X. A., Peregon, A., Pfeil, B., Pierrot, D., Poulter, B., Rehder, G., Reimer, J., Rödenbeck, C., Schwinger, J., Séférian, R., Skjelvan, I., Stocker, B. D., Tian, H., Tilbrook, B., Tubiello, F. N., van der Laan-Luijkx, I. T., van der Werf, G. R., van Heuven, S., Viovy, N., Vuichard, N., Walker, A. P., Watson, A. J., Wiltshire, A. J., Zaehle, S., and Zhu, D.: Global Carbon Budget 2017, Earth Syst. Sci. Data, 10, 405–448, https://doi.org/10.5194/essd-10-405-2018, 2018.
Li, H., Xiao, Z., Wei, J., and Wang, G.: The seasonal and diurnal variation characteristics of soil moisture at different depths from observational sites over the Tibetan Plateau, Remote Sens.-Basel, 14, 5010, https://doi.org/10.3390/rs14195010, 2022.
Li, J., Wang, S., Zhu, J., Wang, D., and Zhao, T.: Accelerated shifts from heatwaves to heavy rainfall in a changing climate, npj Clim. Atmos. Sci., 8, 214, https://doi.org/10.1038/s41612-025-01113-w, 2025a.
Li, W., Zhang, Y., Zhang, X., He, Y., Hou, G., Shen, R., Xia, J., Zheng, J., Jiang, Q., and Zhu, J.: The drivers of respiration shift from soil nutrients to water with the increase in temperature, Funct. Ecol., 39, 2790–2802, https://doi.org/10.1111/1365-2435.70146, 2025b.
Liang, G., Stefanski, A., Eddy, W. C., Bermudez, R., Montgomery, R. A., Hobbie, S. E., Rich, R. L., and Reich, P. B.: Soil respiration response to decade-long warming modulated by soil moisture in a boreal forest, Nat. Geosci., 17, 905–911, https://doi.org/10.1038/s41561-024-01512-3, 2024.
Liang, N., Hirano, T., Zheng, Z.-M., Tang, J., and Fujinuma, Y.: Soil CO2 efflux of a larch forest in northern Japan, Biogeosciences, 7, 3447–3457, https://doi.org/10.5194/bg-7-3447-2010, 2010.
Liu, Y., Yang, Y., and Song, J.: Variations in global soil moisture during the past decades: climate or human causes?, Water Resour. Res., 59, e2023WR034915, https://doi.org/10.1029/2023WR034915, 2023.
Liu, Y., Wang, H., Schindlbacher, A., Liu, S., Yang, Y., Tian, H., Chen, L., Ming, A., Wang, J., Li, J., and Tian, Z.: Soil respiration related to the molecular composition of soil organic matter in subtropical and temperate forests under soil warming, Soil Biol. Biochem., 201, 109661, https://doi.org/10.1016/j.soilbio.2024.109661, 2025.
Lloyd, J. and Taylor, J. A.: On the temperature dependence of soil respiration, Funct. Ecol., 8, 315–323, https://doi.org/10.2307/2389824, 1994.
Mao, Y., Chen, K., Ji, W., and Yang, Y.: The response of soil respiration to temperature and humidity in the thermokarst depression zone of the headwater wetlands of Qinghai Lake, Biology, 13, 437, https://doi.org/10.3390/biology13060437, 2024.
Matsumoto, K., Terasawa, K., Taniguchi, S., Ohashi, M., Katayama, A., Kume, T., and Takashima, A.: Spatial and seasonal variations in soil respiration in a subtropical forest in Okinawa, Japan, Ecol. Res., 38, 479–490, https://doi.org/10.1111/1440-1703.12386, 2023.
Mayer, M., Prescott, C. E., Abaker, W. E. A., Augusto, L., Cécillon, L., Ferreira, G. W. D., James, J., Jandl, R., Katzensteiner, K., Laclau, J. P., Laganière, J., Nouvellon, Y., Paré, D., Stanturf, J. A., Vanguelova, E. I., and Vesterdal, L.: Tamm Review: Influence of forest management activities on soil organic carbon stocks: a knowledge synthesis, Forest Ecol. Manag., 466, 118127, https://doi.org/10.1016/j.foreco.2020.118127, 2020.
Muggeo, V. M.: Estimating regression models with unknown break-points, Stat. Med., 22, 3055–3071, https://doi.org/10.1002/sim.1545, 2003.
Podzikowski, L. Y., Billings, S. A., and Bever, J. D.: Plant functional diversity shapes soil respiration response to soil moisture availability, Ecosystems, 28, 15, https://doi.org/10.1007/s10021-024-00946-5, 2025.
Prigoliti, M., Chiofalo, M. T., Petruzzellis, F., Lo Gullo, M. A., and Trifilò, P.: Ecophysiological behavior of Fagus sylvatica L. growing at its southern distribution limit: insights for understanding the fate of the European beech under warmer and dryer growth conditions, Forests, 14, 2058, https://doi.org/10.3390/f14102058, 2023.
Rodtassana, C., Unawong, W., Yaemphum, S., Chanthorn, W., Chawchai, S., Nathalang, A., Brockelman, W. Y., and Tor-ngern, P.: Different responses of soil respiration to environmental factors across forest stages in a Southeast Asian forest, Ecol. Evol., 11, 15430–15443, https://doi.org/10.1002/ece3.8248, 2021.
Sáez-Sandino, T., García-Palacios, P., Maestre, F. T., Plaza, C., Guirado, E., Singh, B. K., and Delgado-Baquerizo, M.: The soil microbiome governs the response of microbial respiration to warming across the globe, Nat. Clim. Change, 13, 1382–1387, https://doi.org/10.1038/s41558-023-01868-1, 2023.
Schindlbacher, A., Schnecker, J., Takriti, M., Borken, W., and Wanek, W.: Microbial physiology and soil CO2 efflux after 9 years of soil warming in a temperate forest: no indications for thermal adaptations, Glob. Change Biol., 21, 4265–4277, https://doi.org/10.1111/gcb.12996, 2015.
Schnecker, J., Spiegel, F., Li, Y., Richter, A., Sandén, T., Spiegel, H., Zechmeister-Boltenstern, S., and Fuchslueger, L.: Microbial responses to soil cooling might explain increases in microbial biomass in winter, Biogeochemistry, 164, 521–535, https://doi.org/10.1007/s10533-023-01050-x, 2023.
Wang, B., Zha, T. S., Jia, X., Wu, B., Zhang, Y. Q., and Qin, S. G.: Soil moisture modifies the response of soil respiration to temperature in a desert shrub ecosystem, Biogeosciences, 11, 259–268, https://doi.org/10.5194/bg-11-259-2014, 2014.
Wang, C., Morrissey, E. M., Mau, R. L., Hayer, M., Piñeiro, J., Mack, M. C., Marks, J. C., Bell, S. L., Miller, S., Schwartz, E., Dijkstra, P., Koch, B. J., Stone, B. W., Purcell, A. M., Blazewicz, S. J., Hofmockel, K. S., Pett-Ridge, J., Hungate, B. A., and Allison, S. D.: The temperature sensitivity of soil: microbial biodiversity, growth, and carbon mineralization, ISME J., 15, 2738–2747, https://doi.org/10.1038/s41396-021-00959-1, 2021.
Wang, M., Dungait, J. A. J., Wei, X., Ge, T., Hou, R., Ouyang, Z., Zhang, F., and Tian, J.: Long-term warming increased microbial carbon use efficiency and turnover rate under conservation tillage system, Soil Biol. Biochem., 172, 108770, https://doi.org/10.1016/j.soilbio.2022.108770, 2022.
Win, K. and Sato, T.: Influence of stand characteristics and management activities on aboveground carbon storage in Japanese cedar and cypress plantations: sustainable management implications, Trees, Forests and People, 15, 100702, https://doi.org/10.1016/j.tfp.2024.100702, 2024.
Xu, L., Baldocchi, D. D., and Tang, J.: How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature, Global Biogeochem. Cy., 18, GB4002, https://doi.org/10.1029/2004GB002281, 2004.
Yan, J., Tong, M., Liu, J., Li, J., and Li, H.: Temperature and moisture sensitivities of soil respiration vary along elevation gradients: an analysis from long-term field observations, Sci. Total Environ., 912, 169150, https://doi.org/10.1016/j.scitotenv.2023.169150, 2024.
Yan, Y., Wang, X., Wohlfahrt, G., and Huang, N.: Seasonal and interannual variations of soil heterotrophic respiration and autotrophic respiration in subtropical forests of southeast China: independent process-based models, Geosci. Lett., 12, 27, https://doi.org/10.1186/s40562-025-00399-1, 2025.
Yan, Z., Bond-Lamberty, B., Todd-Brown, K. E., Bailey, V. L., Li, S., Liu, C., and Liu, C.: A moisture function of soil heterotrophic respiration that incorporates microscale processes, Nat. Commun., 9, 2562, https://doi.org/10.1038/s41467-018-04971-6, 2018.
Yu, J.-C., Chiang, P.-N., Lai, Y.-J., Tsai, M.-J., and Wang, Y.-N.: High rainfall inhibited soil respiration in an Asian monsoon forest in Taiwan, Forests, 12, 239, https://doi.org/10.3390/f12020239, 2021.
Zhang, Z., Li, Y., Williams, R. A., Chen, Y., Peng, R., Liu, X., Qi, Y., and Wang, Z.: Responses of soil respiration and its sensitivities to temperature and precipitation: a meta-analysis, Ecol. Inform., 75, 102057, https://doi.org/10.1016/j.ecoinf.2023.102057, 2023.
Zhu, M., De Boeck, H. J., Xu, H., Chen, Z., Lv, J., and Zhang, Z.: Seasonal variations in the response of soil respiration to rainfall events in a riparian poplar plantation, Sci. Total Environ., 747, 141222, https://doi.org/10.1016/j.scitotenv.2020.141222, 2020.