the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 25 Jun 2026
| 25 Jun 2026
Invertebrate communities on seasonal snow patches in the European lowlands are shaped by tree canopy-derived organic inputs
Dzmitry Lukashanets
Krzysztof Zawierucha
Jolita Petkuvienė
Donata Overlingė
Diana Vaičiūtė
Greta Kalvaitienė
Marija Kataržytė
The cryosphere encompasses a wide range of habitats that support cold-adapted communities. Seasonal snow patches (SSPs) in lowlands are underexplored cryohabitats, characterised by a short persistence period (late winter–early spring) and the presence of trees around, in contrast to large vegetation-free high-mountain and polar ecosystems. To provide the first assessment of organisms from SSPs, we focused on invertebrate diversity and densities from 40 sites in the Baltic States in relation to physicochemical parameters (i.e., suspended solids, chlorophyll a, nutrients), microalgae, and surrounding landscape features (i.e., tree canopy cover). SSPs appeared to be an important spot for bdelloid rotifers (Bdelloidea), tardigrades (Tardigrada), and nematodes (Nematoda), which together accounted for 60 %–100 % of all invertebrates, reaching densities >7000 ind m−2. Acari and Insecta were less abundant, whereas other invertebrates occurred only sporadically. The community was strongly determined by surroundings (trees), which supply snow ecosystems with organic and inorganic matter. Chlorophyll a, particulate phosphorus, total suspended solids and organic debris were strongest predictors of invertebrate distribution. The canopy cover also influenced invertebrate communities, highlighting the importance of the organic deposition from trees and also suggesting that trees may be a source of microscopic invertebrates to the snow. Results demonstrate the importance of SSPs as overlooked ephemeral habitats and can be used as a baseline for future changes in snow communities in temperate regions.
- Article
(3330 KB) - Full-text XML
-
Supplement
(336 KB) - BibTeX
- EndNote




Glaciers, snow, and sea ice, known as the cryosphere, shape the functioning of the Earth's systems by providing multiple services spanning from regulation of albedo to delivery of nutrients downstream and shaping biodiversity (e.g., Hodson et al., 2005; Domine, 2019; Bosson et al., 2023; Dong et al., 2023). But the cryosphere itself also supplies a range of habitats for cold-adapted biota – microbial communities and complex organisms (e.g., Anesio and Laybourn-Parry, 2012; Gobbi and Lencioni, 2020; Zawierucha et al., 2021). The cryospheric ecosystems, including glaciers, snow fields, are biologically active during summer when temperatures rise and liquid water is available for activity and reproduction of cold-living organisms (Hodson et al., 2008). For example, glaciers host a variety of bacteria, phototrophs, and invertebrates inhabiting mostly supraglacial environments, i.e., located on the glacier surface, such as water-filled bodies known as cryoconite holes (Zawierucha et al., 2015; Rozwalak et al., 2022; Pittino et al., 2023), or the spaces between ice crystals on the surface ice (Shain et al., 2016; Hotaling et al., 2019). Invertebrates on glacier surfaces represent higher levels of trophic chains, often are top consumers, with springtails (Collembola) dominating the surface ice, while tardigrades (Tardigrada) and rotifers (Rotifera: Bdelloidea) prevail in cryoconite holes (Zawierucha et al., 2018; Hotaling et al., 2020; Crosta et al., 2025; Dimattia et al., 2025). Nematodes (Nematoda), the most distributed group of microfauna in the world, are much less common on glaciers (Zawierucha et al., 2021; Devetter et al., 2025). Although glaciers are investigated in terms of biodiversity and ecology worldwide, other components of the cryosphere, like snow fields and snow patches, receive less attention.
In mountains, snow surface is known to host snow algae which bloom during the melting season, and change the colour of the surface, then trigger changes such as absorbing solar radiation, and shape the environment for bacteria, fungi and even invertebrates (Fiołka et al., 2020; Hoham and Remias, 2020; Procházková et al., 2024). Microalgae, the primary producers on the snow surface, are among the best recognised biotic components of snow (Leya, 2013; Segawa et al., 2018; Remias et al., 2023), while invertebrates, top-consumers, are known to a lesser extent. This knowledge gap limits our understanding of the bio-albedo feedback, and recognition of the biodiversity and biomass, which may trigger further changes following snowmelt. To date, only a few recent studies have reported data on snow-dwelling microscopic invertebrates (microinvertebrates, microfauna), covering mountains in Slovakia (Hanzelová et al., 2018), Canada (Yakimovich et al., 2020), and a single mountainous locality in northern Japan (Ono et al., 2021, 2022, 2025; Ono and Takeuchi, 2025). In contrast, studies on larger invertebrates on snow have much longer history and larger geography (e.g., Brauer, 1871; Chapman, 1954; Hågvar, 2010; Valle et al., 2024). This gap is alarming, given the impacts of global warming on the rapid loss of cryospheric ecosystems, which are closely linked to changes in snowfall patterns and amounts (Krasting et al., 2013). Many glacier invertebrates seem to be glacier-dependent, and cannot survive without glacial environments (Zawierucha et al., 2019). Similar assumptions were presented for snow microinvertebrates, however, the knowledge on these animals is insufficient and restricted to a few mountainous sites (e.g., Ono et al., 2021, 2022, 2025) without data for seasonal snow in lowlands. Even though invertebrates which inhabit snow or ice are freeze-tolerant, survive freeze-thaw cycles and have the capacity to enter dormant stages and survive even millennia (Shmakova et al., 2021), they are poorly adapted to increasing temperatures and are, therefore, particularly vulnerable to the effects of global warming (Zawierucha et al., 2019; Zawierucha and Shain, 2019).
Unlike the seasonal snow in mountains of Japan (Ono et al., 2021, 2022, 2025; Ono and Takeuchi, 2025), which persists from November to June, the last snow in lowlands of temperate Europe typically vanishes by late March and usually does not have algal blooms. These patches of late snow cover, or seasonal snow patches (SSPs), represent some of the most ephemeral and fragile components of the global cryosphere, and are particularly vulnerable to intensifying warming trends in Europe (Voigt et al., 2010; European Environment Agency, 2025).
SSPs are confined to late winter and early spring in temperate climates, predominantly occurring in the sheltered places under the canopy of forested areas. The remaining SSPs exhibit characteristics similar to those of glacial habitats: the presence of a thin water film resulting from temperatures above zero and increased solar radiation, along with albedo-reducing dark materials. However, in contrast to snow in polar and many alpine ecosystems, in SSPs in lowland Europe these organic particles might be deposited by falling from trees. Collectively, these factors suggest that SSPs may provide conditions beneficial for cold-adapted microinvertebrates. Considering that comprehensive information regarding dominant groups, their abundances, and distribution patterns in SSPs is entirely lacking, here we address this knowledge gap. The absence of such data for lowlands in temperate climates prevents understanding the role of SSPs for fauna, other organisms and ecosystem function (Kivinen et al., 2012).
The study provides an inventory of invertebrates, with a focus on poorly studied microfauna, inhabiting seasonal snow patches in the Baltic States, a representative lowland region of Europe with temperate climate. The aim of the study was to evaluate how, and to what extent, these ephemeral habitats support diverse metazoan communities and to assess how the surrounding landscape (e.g., type of forests, trees and their canopies) influence the seeding of invertebrates onto the snow surface. We hypothesize that deposition of food sources (organic matter) from the surrounding, particularly forest canopies, on snow surfaces supports the increase of microinvertebrate diversity and densities during early spring. Data obtained through a comprehensive approach, including biotic and abiotic variables on the surface of seasonal snow, will contribute to broader assessments of the potential loss of cryosphere habitats suitable for invertebrates in lowland regions. This study provides a background for understanding the SSPs as a habitat, spot for invertebrate reproduction or a source of organic matter to forest floor after snow melt.
2.1 Study area
Snow was collected in March 2023 and February–March 2024, in Lithuania, Latvia, and Estonia, at latitudes ranging from 55.22 to 59.00° N and at longitudes ranging from 24.59 to 27.16° E. The elevation varied from 5 to 170 m above sea level (Fig. 1). The sampling dates were chosen following the principle of collecting the last SSPs (las snow of the season) and therefore differed across the Baltic States (i.e., end of February in Lithuania, March in Latvia and southern Estonia, end of March in central and northern Estonia).
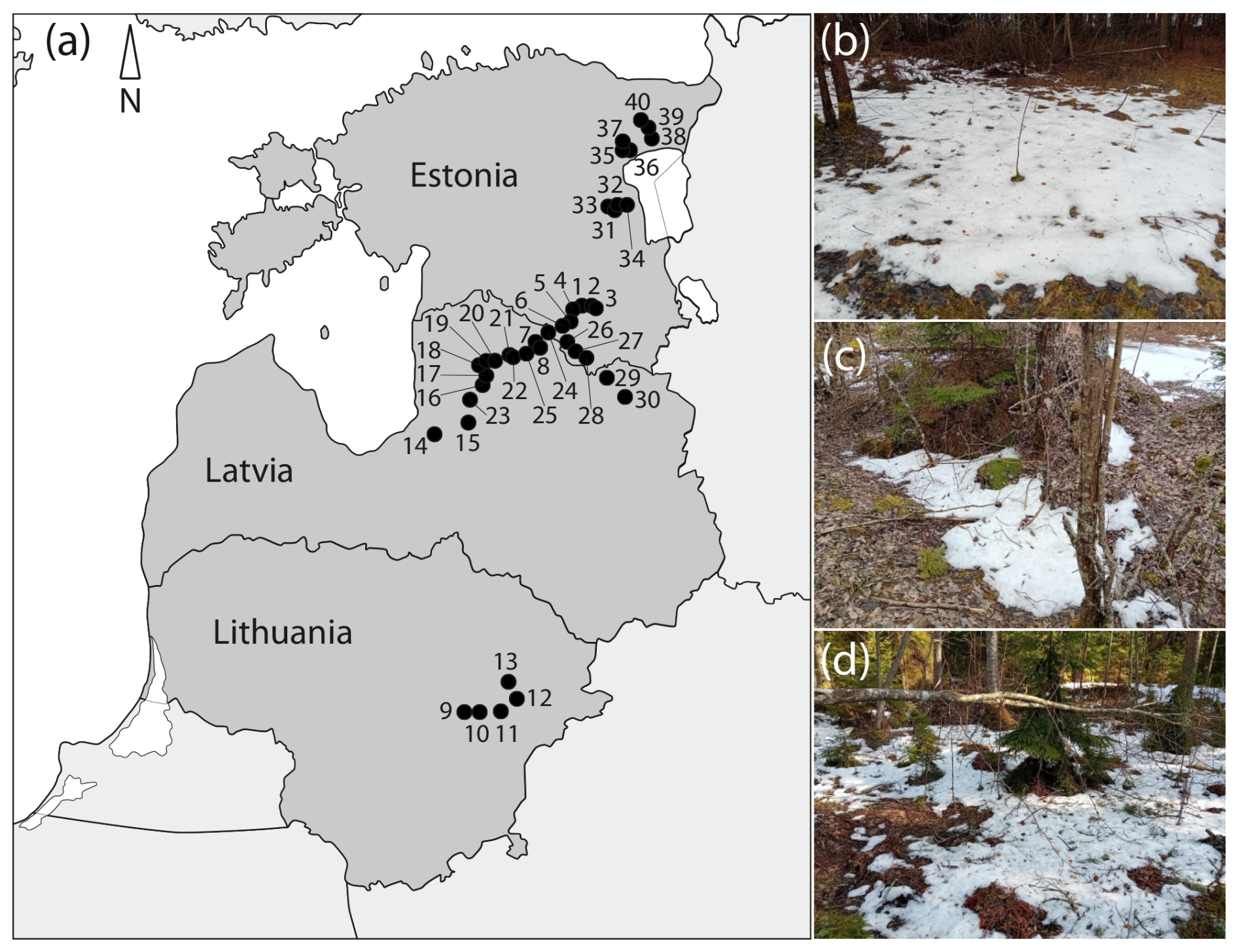
Figure 1Locations of sampling sites ## 1–40 in Lithuania, Latvia, and Estonia, 2023–2024 (a); SSPs at sampling sites # 11 (b), # 20 (c), and # 36 (d). For coordinates of sampling sites, see Table S1.
Snow-covered areas were selected in forests (mainly spruce, mixed spruce-birch, and pine-spruce-birch), in forest edges, and in shrubs. A total of 40 sites were sampled (Table S1 in the Supplement), each site represented one SSP. The priority was given to older and more undisturbed SSPs (i.e., showing no visible signs of human or animal trampling, mechanical disturbance, or recent surface disruption, and maintaining an intact snow surface) of different sizes (from less than 1 m2 to more than 200 m2). Dumps resulting from snow removal from forest roads and fresh, recently fallen snow were avoided.
2.2 Data collection
From each site, snow was collected using an iron or plastic shovel and placed in a 20 L plastic bucket with a locking lid. Tree branches and large leaves were removed before sampling. Other organic debris (conifer needles, and organic particles originated from surroundings, such as small pieces of tree bark, wood, etc. smaller than 1 cm) were considered as a part of the habitat and taken with the snow. The limited snow thickness did not allow for the sampling of distinct layers, as described in Ono et al. (2025) and Ono and Takeuchi (2025), therefore only upper layer (2–3 cm of depth) was collected. However, the thickness of each SSP exceeded 5 cm, thereby avoiding the snow-soil interface. The sampling was performed following the principle of taking the maximum possible area of the snow, until the whole SSP was sampled or, in cases of larger patches, until bucket became full. Due to the varying density and structure of the snow, the final volumes placed in buckets differed, ranging from 7.3 to 13.1 L between sampling sites. The sampled area varied from 0.34 to 2.8 m2. To analyse the composition and biomass of microalgae (potential food resource for rotifers and other microinvertebrates), a small portion of the collected snow from each SSP was placed in a 50 mL Falcon tube and fixed after melting with acetic Lugol's solution in a proportion of 0.5 mL solution per 100 mL sample.
At each site, we recorded surrounding landscape features that could potentially influence the densities of invertebrates on the snow surface. We measured the distance from the sampled area to the nearest large tree (diameter >10 cm) as well as the total number of trees of any size and shrubs in close proximity to the sampled area (within a 1 m radius of its edges).
All sampling sites were photographed and georeferenced. Additionally, one photo was taken from the middle of the sampled area at a height of 1 m above the ground surface vertically upwards and towards the sky to assess how the area was covered by canopies of the trees (Fig. 2). The total area of the snow collected for the sample was measured, as well as the total area of the SSP.

Figure 2Sampling sites # 16 (a), # 22 (b), and # 36 (c) representing different level of canopy cover: 5 %, 24 %, and 58 %, respectively. For coordinates of sampling sites, see Table S1.
2.3 Sample processing
To avoid heat shock, snow samples were melted slowly by placing buckets containing snow in a +5 °C room for 2–3 d. After complete melting, the meltwater, together with organic particles collected with snow, was measured by volume, gently mixed and subsampled for different analyses. Approximately 4–6 L were taken for chemical and nutrient analysis, while the remaining meltwater (approximately 3–7 L) was used for analyses of invertebrate densities.
2.3.1 Nutrients and chemical parameters
pH and conductivity were measured immediately after melting using the multiparameter probe WTW Multi 9630 IDS. Additionally, alkalinity was measured by the titration method (Federation and Aph Association, 2005) before filtration through Whatman GF/F (0.7 µm pore size) filters.
In all melted samples dissolved (, , , TDN) and total nitrogen (TN), dissolved inorganic and total dissolved phosphorus (DIP, TDP), total phosphorus (TP), organic carbon (Corg), total suspended matter solid (TSS), and chlorophyll a (Chl a) concentrations were measured. Aliquots for DIP, , , , TDN and TDP measurements were immediately filtered through Whatman GF/F (0.7 µm pore size), while unfiltered water samples were used for TP and TN measurements. For Corg, Chl a and TSS, snow water samples were filtered on the Whatman GF/F (0.7 µm pore size), and the filters were frozen for later analysis. For Corg and TSS measurements, filters were precombusted at 550 °C for 4 h.
Spectrophotometric methods were used for analysis of nutrients: (International Organisation for Standardisation, 1993), (International Organisation for Standardisation, 1984), DIP, TDP and TP (International Organisation for Standardisation, 2004), and Corg (International Organisation for Standardisation, 1995). , TDN and TN were measured spectrophotometrically by recording spectra and calculating the second derivative according to Ferree and Shannon (2001). Particulate nitrogen (PN) and phosphorus (PP) were calculated by subtracting the total form from the total dissolved form. Filters for total suspended solids were dried at 60 °C for 24 h and particulate weight per L was calculated.
Pigment concentrations were determined spectrophotometrically. Chl a concentration was estimated using the trichromatic method of Jeffrey and Humphrey (1975).
2.3.2 Abundance and biomass of microalgae
The composition of the microalgal communities was analyzed under an inverted microscope (LEICA DMI 3000, Leica Microsystems) at magnifications of 400×, according to the method described by Utermöhl (1958). According to European Committee for Standardization recommendations (2006), the abundance of microalgae (counts L−1) was calculated by multiplying the number of units counted with the coefficient C (dm3), calculated using Eq. (1):
where A is the cross-section area of the top cylinder of the combined sedimentation chamber (the usual inner diameter is 25.0 mm, giving A=491 mm2), N is the number of counted fields or transects, a is the area of single field or transect, and V is the volume (cm3) of sedimented aliquot.
The biomass of microalgae (mg L−1) was calculated by the allocation of algal species (genus) to size classes according to the scheme of Olenina et al. (2006) and updated appendix available at ICES website (https://www.ices.dk/data/vocabularies/Pages/default.aspx, last access: 5 November 2025). The biomass of microalgae (mg L−1) was calculated based on Eq. (2), as recommended by HELCOM (2021):
where VCU is the volume of the counting unit (µg).
2.3.3 Invertebrate extraction
Water after melting was filtered using a limnological or similar net with a 50 µm mesh size. The filtered residue with invertebrates was removed from the net with a spatula and tweezers and placed in 50 mL plastic Falcon tubes (2–5 per sampling site, depending on how many particles there were on it). Tubes with concentrated samples were stored in −20 °C until processing.
To analyse the taxonomic composition and abundances of snow invertebrates, contents of tubes were slowly refrozen at +5–+7 °C and placed in Petri dishes with a gridded bottom. Sorting, counting and isolation of invertebrate individuals were conducted using dissection microscopy (stereomicroscope Olympus SZ61 with bottom illumination and magnification of ×6.7–45).
2.4 Data analysis
Invertebrate densities obtained from the part of the sample were recalculated to the whole volume of the sample and then expressed in number of individuals per one square meter (ind m−2) and per one liter (ind L−1).
To explain the invertebrate presence and their distributional patterns on the snow we used all recorded and measured environmental variables (Table 1).
Table 1Environmental variables used for explaining the patterns of distribution of invertebrates in SSPs.
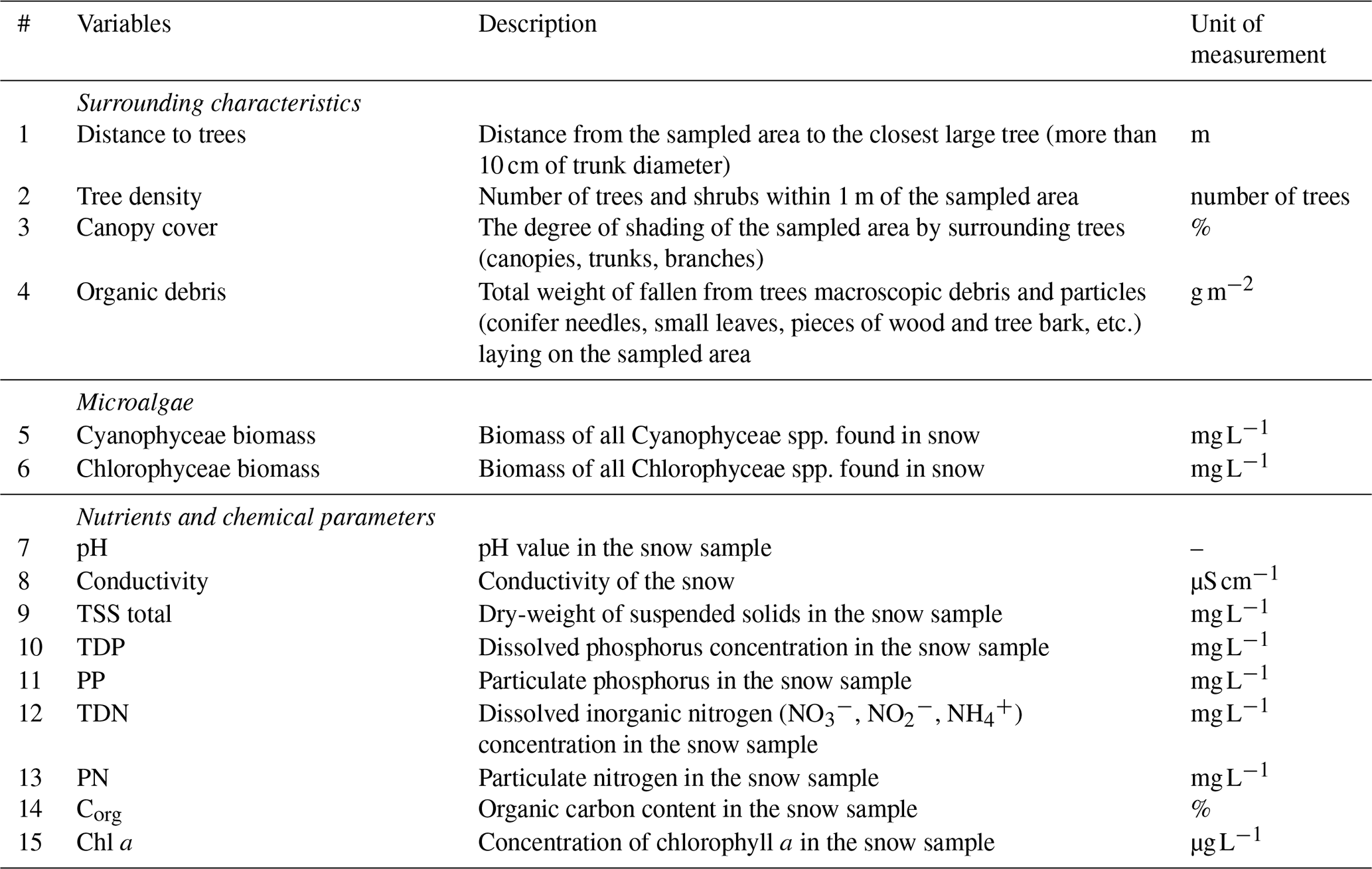
Since the development of invertebrates on the seasonal snow may be fostered by surrounding trees and particularly their canopies, we focused on characteristics describing the degree of forest cover in the immediate vicinity of each snow patch. We identified the following elements of the surrounding landscape as the most relevant: (i) distance to the nearest large tree/trees, (ii) number of trees and shrubs within a 1 m radius from the edges of the sampled area, and (iii) canopy cover (shadiness of sampled area). To quantify the latter, we calculated the proportion of tree canopies and trunks visible in upward-facing photographs taken from the centre of each sampled area, using object selection tools and colour histogram analysis in the Adobe Photoshop program. The canopy cover was expressed in percentages: 0 % – totally open area with no canopies above, 100 % – area completely covered by canopies.
Organic debris mass was considered as a factor, which is likely influenced by the surrounding landscape (the rate of material deposition from trees). Although it was impossible to determine the age of each snow patch, the presence of organic debris was used as an indicator of the longer existence of the patch (due to the longer accumulation of debris over time). To quantify organic debris mass, all particulate matter (conifer needles, leaves, wood fragments and bark, etc.) was carefully separated from each sample after analysis of invertebrates and dried at room temperature and then weighed to the nearest 0.01 g using laboratory scales.
When estimating the biomass of the microalgae, the most abundant algal classes were considered separately. Nutrient composition and chemical parameters were assessed in three replicates per sample, and mean values were subsequently calculated.
All plots and statistical analyses (Kruskal Wallis test, Spearman correlation, transformation-based redundancy ordination analysis) were performed in R using R Studio environment and following packages: “vegan” v2.5-7 (Oksanen et al., 2020), “ggplot2” v3.5.1 (Wickham, 2016), and “Hmisc” v4.7-1 (Harrel, 2022).
3.1 Characteristics of the sites
Characteristics of the SSPs and its surrounding environment varied significantly. The distance from the edge of sampled area to the nearest large tree ranged from 0.03 to 20 m, with more than half of the sites (n=26) located closer than 1 m. The maximum number of any trees within a 1 m radius was 19, while four SSPs were not surrounded by any trees or shrubs. Canopy cover, another parameter describing the degree of forest cover for each SSP, ranged from 0.4 % to 69.8 %. Since no significant correlations were found between three aforementioned environmental variables, they can be treated as independent variables in subsequent analyses. At the same time, the number and cover of trees around the snow influenced the surface accumulation of organic debris, whose mass ranged substantially (0.015–5.81 g m−2). A negative correlation between organic debris mass and distance to the nearest large tree (; p=0.0115) was observed as well as positive correlation between organic debris mass and canopy cover (Rs=0.45; p=0.0102).
All data on environmental variables are given in Table S2 in the Supplement.
3.2 Biomass of microalgae in SSPs
As expected, no algal blooms were observed. Two main groups of autotrophic microorganisms were identified in SSPs: cyanobacteria (Cyanophyceae) and green algae (Chlorophyceae), which were present in 97 % and 100 % of the analysed samples, respectively (Table S2). All cyanobacteria belonged to the genus Chroococcus, with abundances ranging from 50.8×103 to 2408.3×103 counts L−1 and biomass values ranging from 0.003 to 0.349 mg L−1. Green algae, represented by Chloromonas spp., exhibited higher abundances and biomass levels, ranging from 49.9×103 to 13 149.8×103 counts L−1 and from 0.012 to 3.347 mg L−1, respectively. Cryptophytes (Cryptomonas, Cryptophyceae) were recorded in only one sample, at low biomass (0.003 mg L−1).
3.3 Nutrients and chemical parameters in SSPs
The environmental variables in SSPs exhibited moderate variation across the studied samples (Table S2). The pH ranged from 5.40 to 6.62, indicating slightly acidic conditions. Conductivity values varied between 3.9 and 30.7 µS cm−1, reflecting generally low mineralization, and TSS showed a wide range from 36.5 to 1376.7 mg L−1. TDP was low, ranging from 0.005 to 0.050 mg L−1, whilst PP was higher, varying from 0.035 to 0.330 mg L−1. TDN ranged between 0.139 and 0.512 mg L−1. PN showed the widest variation, from 0.471 to 2.902 mg L−1. Corg ranged from 16.4 % to 55.9 %, indicating moderate to high organic content in the studied SSPs.
3.4 Invertebrates in SSPs: overview of taxonomy
Invertebrates were found in all snow patches, with five phyla recorded: rotifers (Rotifera), nematodes (Nematoda), tardigrades (Tardigrada), arthropods (Arthropoda), and annelids (Annelida). Rotifers were represented only by bdelloids (subclass Bdelloidea) and were recorded at every site (Table 2). At 18 sites they accounted for more than 50 % of all invertebrates, and at one site they were the only taxon recorded. Other dominant taxa in SSPs were tardigrades and mites (Acari), both found at 38 sites, and nematodes, recorded at 36 of the 40 sites.
Table 2Invertebrates in SSPs and their frequencies of occurrence (%).
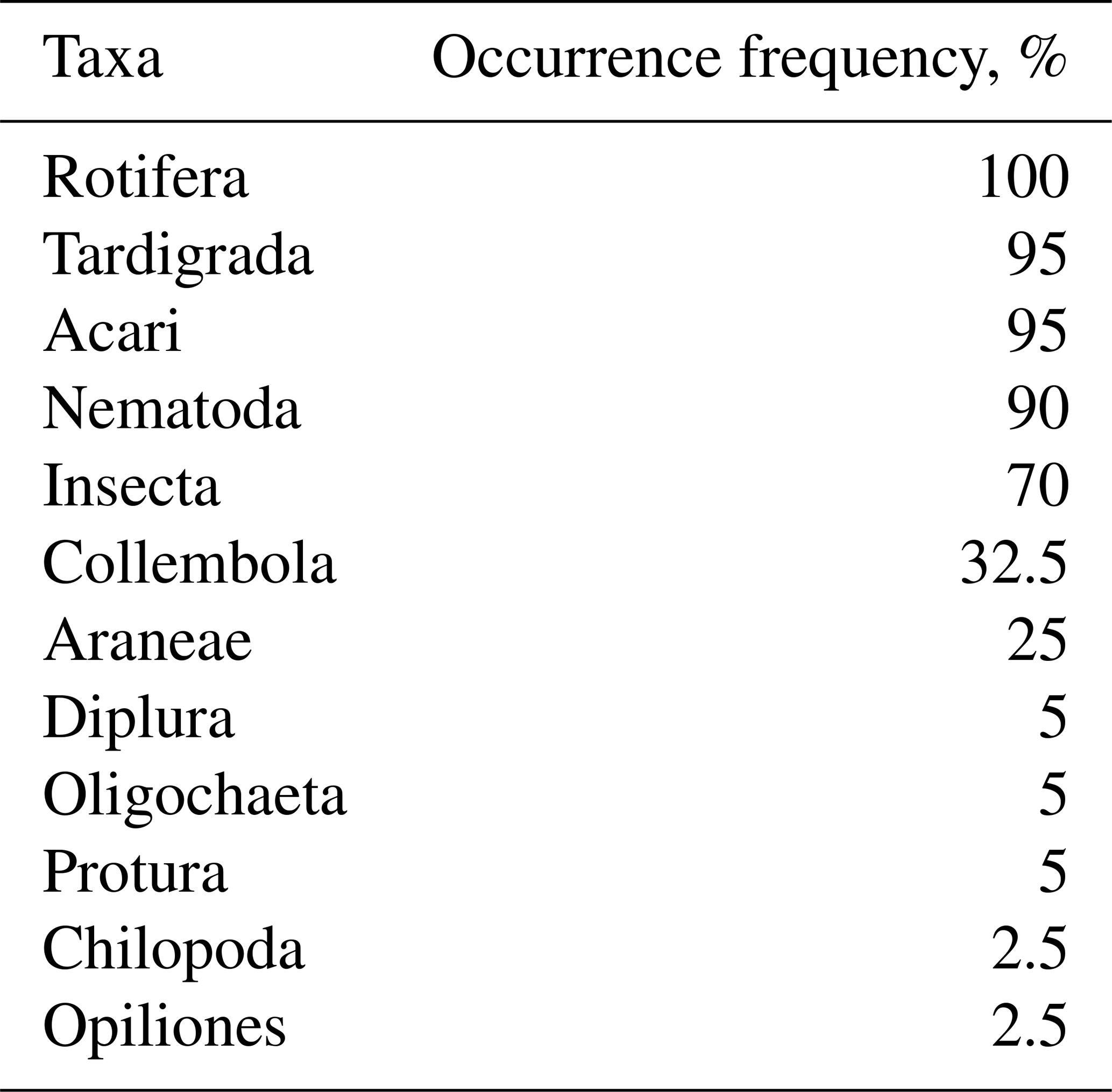
Among arthropods, in addition to mites, the most widespread groups were insects belonging to the orders Hymenoptera, Coleoptera, Hemiptera, and Diptera (chironomid larvae). Springtails (Collembola) were present at 32.5 % of the sites, and spiders (Araneae) at 25 %. Other arthropods occurred in only a few SSPs: diplurans (Diplura) and proturans (Protura) at two sites each, and centipedes (Chilopoda) and harvestmen (Opiliones) at one site each. Annelids were represented by oligochaetes (Oligochaeta), recorded in two SSPs.
All densities for invertebrates in SSPs are provided in Table S3 in the Supplement.
3.5 Main abundance patterns of invertebrates in SSPs
The total density of snow invertebrates was 653 ± 1240 ind m−2 (n=40), ranging from 17 to 7188 ind m−2, with bdelloid rotifers being the most abundant taxon (347 ± 722 ind m−2), followed by nematodes (131 ± 297 ind m−2) and tardigrades (129 ± 231 ind m−2) (Fig. 3). These three groups of microscopic invertebrates accounted for the majority of all metazoans, comprising 87 % ± 12.1 % of the total abundance. Although mites were widely distributed, their densities were comparatively low, reaching a maximum of 213 ind m−2, which was less than that of insects (264 ind m−2). Only in two low-density samples mites constitute a substantial proportion of the community – 43 % and 46 %, respectively (however, some specimens were destroyed, likely not active on snow). All other taxa were less abundant, with densities below 10 ind m−2, contributing up to 12 % of total individuals at most.
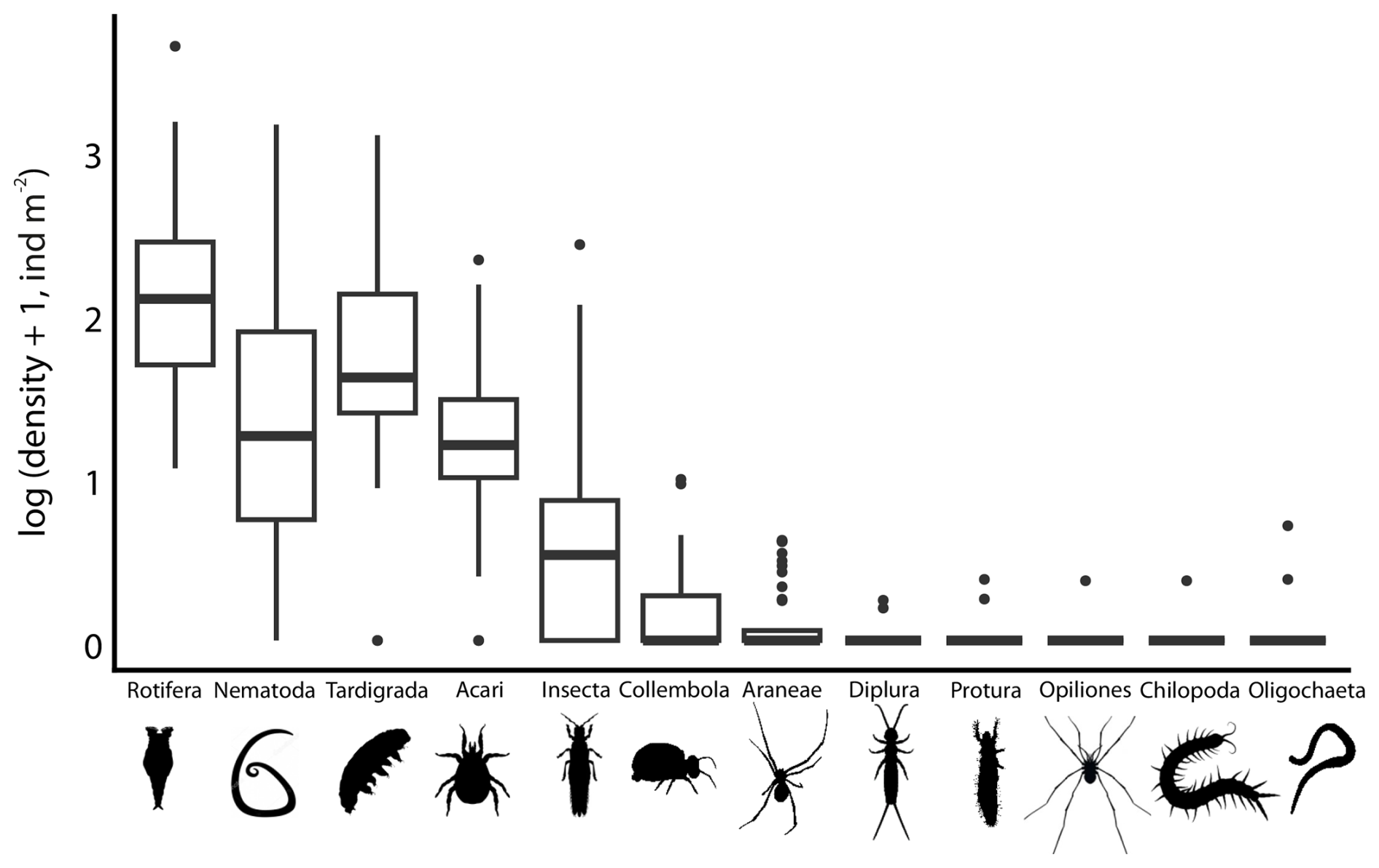
Figure 3Density (ind m−2 of snow) of all invertebrates in SSPs (n=40). Boxes show interquartile ranges; box horizontal midlines are medians; whiskers are maxima and minima; black dots represent outliers.
Possible relationships between invertebrates and microalgae were estimated for taxa presumed to depend on microalgae or cyanobacteria as a food source. The analysis revealed moderate positive correlations between the biomass of cyanobacteria and rotifers (Rs=0.48; p=0.0053), as well as between cyanobacteria and tardigrades (Rs=0.49; p=0.0037) (Fig. 4). Neither significant correlations were detected between nematodes and cyanobacteria, nor between mites and cyanobacteria. No correlations were observed between any of the taxa and green algae biomass.
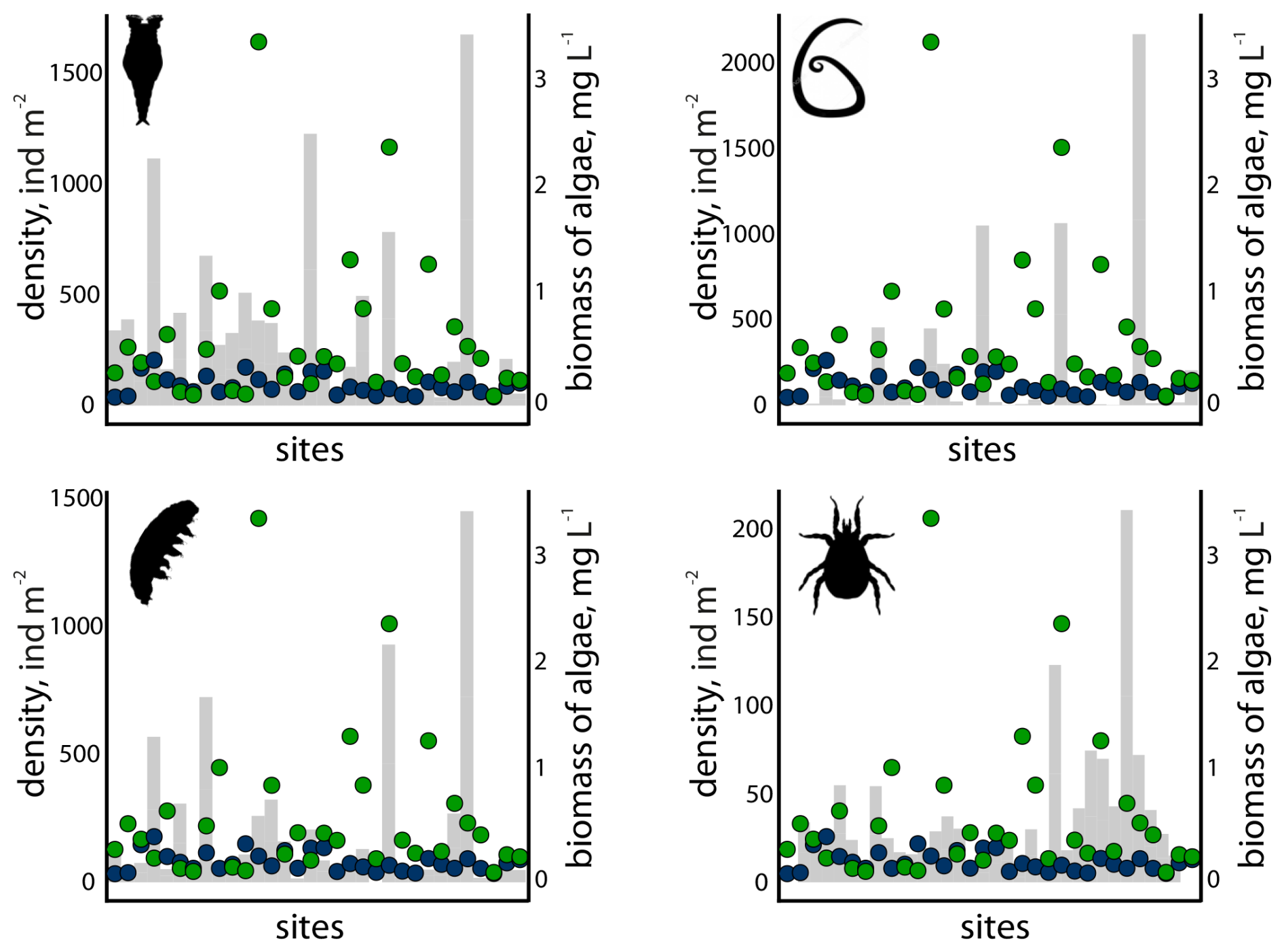
Figure 4Density (grey bars, ind m−2 of snow) of rotifers (top left), tardigrades (bottom left), nematodes (top right) and mites (bottom right) in SSPs (n=40) in relation to the biomass of green algae (green circles) and cyanobacteria (blue circles).
Considering the most abundant invertebrate groups (rotifers, nematodes, tardigrades, mites, insects), a slight increase in density was observed in SSPs located within coniferous and mixed forests compared to those located at deciduous forests and shrubs (Fig. 5). In the latter type of habitats, rotifer densities reached up to 350 ind m−2, while most of the other invertebrates generally occurred at densities below 100 ind m−2. Springtails were recorded only once on the snow in the deciduous forests. In contrast, SSPs located in mixed forests exhibited substantially higher densities of all groups, with rotifers exceeding 1000 ind m−2 in several sites (e.g., 4338 ind m−2 at site 8), along with nematodes (up to 1438 ind m−2) and tardigrades (up to 1238 ind m−2). Comparable values were also recorded in coniferous forests (e.g., site 5 with a density of rotifers 1496 ind m−2). Mite and insect densities in both mixed and coniferous forests were an order of magnitude higher than those observed in deciduous forests (e.g., 264 ind m−2 for insects at site 5), springtails were also more abundant. However, none of the differences in densities among SSPs from different habitat types were statistically significant.
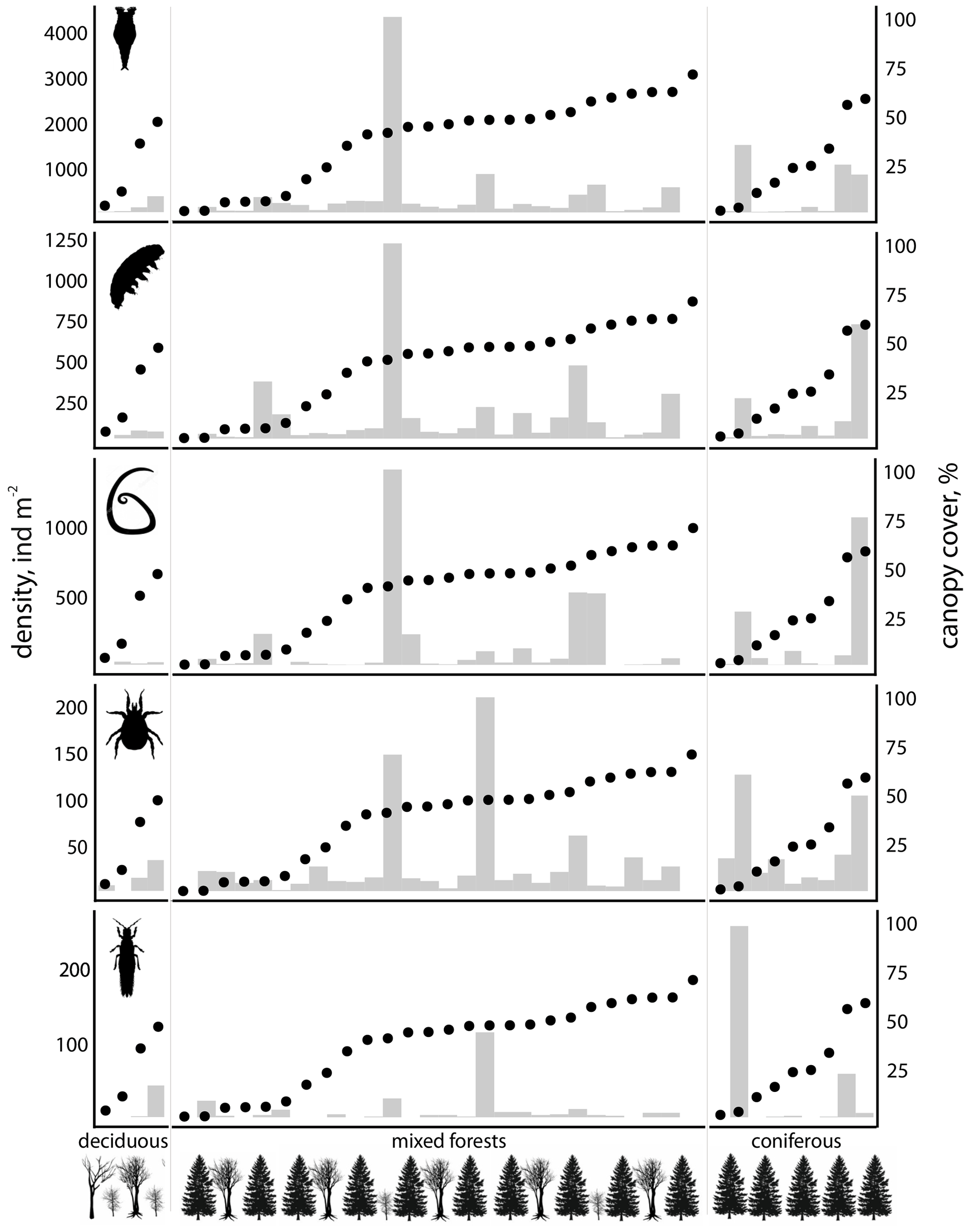
Figure 5Density (ind m−2 of snow) of the most distributed invertebrate taxa (from top to bottom – Rotifera, Tardigrada, Nematoda, Acari, Insecta) in SSPs (n=40) for different types of forest. Sites are shown in the order of increasing canopy cover (%, black dots).
Invertebrate densities were compared with canopy cover, which is considered one of the key predictors of animal distribution (Fig. 5). A moderate positive correlation was detected for rotifers (Rs=0.38; p=0.02), tardigrades (Rs=0.46; p=0.003), and insects (Rs=0.39; p=0.016). However, the strength of invertebrate–canopy relationships varied among forest types. The strongest correlations were observed for rotifers and insects in deciduous forests, despite the low number of sites (Rs=0.96; p<0.001 and Rs=0.94; p=0.047, respectively). In contrast, correlations in coniferous and mixed forests were weak or not statistically significant.
3.6 Invertebrates in SSPs respecting environmental variables
Transformation-based redundancy analysis (tbRDA, scaling type II) was performed for 32 sampling sites (## 9–40), for which all environmental variables were available. All these sites were sampled in second year of research (2024). The density of the invertebrates in SSPs is strongly determined by environmental variables, which contribute 63.4 % of the variation (tbRDA, scaling type II – Fig. 6). The first two axes explain 55.9 % of the entire variation and 88.3 % of the constrained variation. The results of the tbRDA analysis were statistically significant (permutation test: p=0.026). Canopy cover appears to explain only 5.5 % of the variation (p=0.001). The most important factors were Chl a concentration (20.6 %, p=0.001), TSS (16.5 % of the variation, p=0.001), organic debris (11.5 %, p=0.001), and PP (13.6 %, p=0.001). pH, density of the trees, and Corg were not significant. Other variables were statistically significant, however had a minor or almost no contribution (e.g., conductivity – 1.6 %, TDP – 0.5 %).
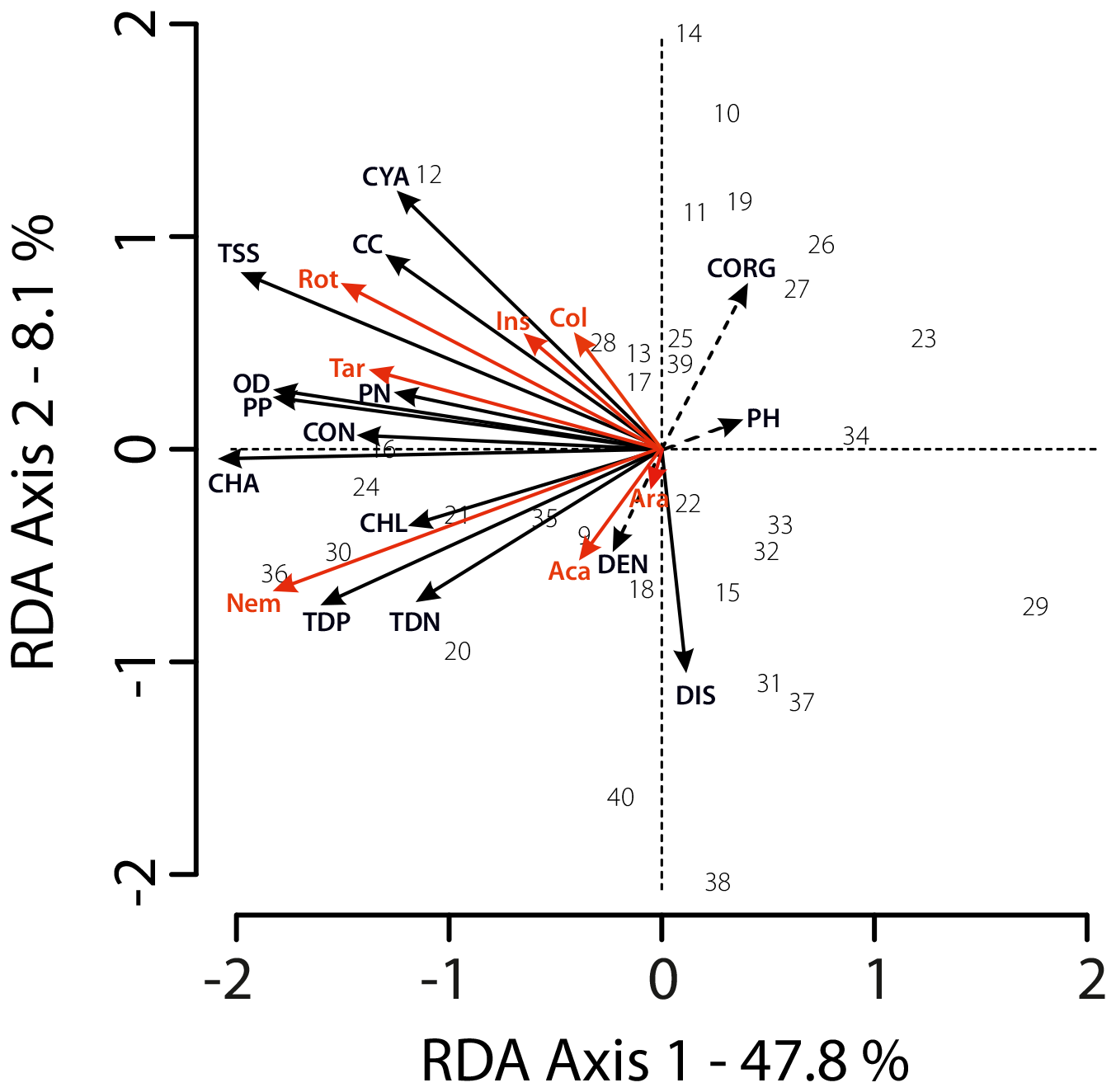
Figure 6Distance-based redundancy analysis (tbRDA) showing relationships between invertebrate taxa (response variables, red arrows) and environmental variables (explanatory variables, black arrows; see Table 1).
The direction of arrows indicates the direction of relationships: angles between arrows reflect correlations among variables, with smaller angles indicating positive relationships and opposite directions indicating negative relationships. Solid black arrows indicate environmental variables that significantly explain variation in community composition (Monte Carlo permutation test, p<0.05), dashed arrows represent non-significant variables (p>0.05).
Sites (n=32) are shown as grey numbers.
Response variables: Aca – Acari, Ara – Araneae, Col – Collembola, Ins – Insecta, Nem – Nematoda, Rot – Rotifera, Tar – Tardigrada. Explanatory variables: CC – canopy cover, CHA – chlorophyll a, CHL – Chlorophyceae biomass, CON – conductivity, CORG – organic carbon, CYA – Cyanophyceae biomass, DEN – tree density, DIS – distance to trees, OD – organic debris.
Correlations with environmental factors varied among taxa. Rotifers showed the strongest associations with canopy cover, TSS and cyanobacteria, similar to insects and springtails. Tardigrades were positively associated with PN, organic debris and PP, nematodes and mites – with dissolved nutrients (TDP and TDN).
The presence of microinvertebrates in snow reflects their broad global occurrence and near-ubiquitous distribution in almost all available limno-terrestrial habitats, especially among rotifers (Ricci, 1987; Fontaneto et al., 2006, 2007) and tardigrades (Ramazzotti and Maucci, 1983; Schill, 2018). Their ability to colonise new environments is enhanced by ability to survive, passive aeolian transport, hydrochory, and zoochory (Bertolani et al., 1990; Fontaneto, 2019; Robertson et al., 2020; Giannetti et al., 2025). However, recent data on the global distribution of some tardigrade species support the assumption of everything is everywhere but environment select (Morek et al., 2021; Zawierucha et al., 2023a). Some of the tardigrades or rotifers need a specific habitat for surviving and reproduction and can be regarded as cold-tolerant species or psychrophiles (Shain et al., 2024). Both taxa are commonly found in glacial habitats (Shain et el., 2016; Zawierucha et al., 2018, 2021, 2023b; Dimattia et al., 2025), while rotifers have also been recorded in the snow in the Antarctic (Murray, 1910). In the seasonal snow at the mountains of Japan, tardigrades and rotifers were found, some of them likely reproduce on snow and are cold-adapted species (Ono et al., 2021). This observation indicates that microfauna is linked with permanent cryosphere and its long evolutionary history, but also inhabits transient cold environments, which are similar in conditions but persist temporarily. In this context, SSPs in lowland Europe can be considered as cryohabitats at the margin, characterized by shorter persistence, yet still supporting microinvertebrate communities.
Apart from rotifers and tardigrades, SSPs also harbour large numbers of nematodes, the third major group of microinvertebrates. Nematodes frequently co-exist with the former two taxa in many types of habitats but are generally absent from permanent cryospheric environments. Their absence from cryoconite holes, with some exceptions, is likely due to a combination of nutrient limitation, rapid fluctuations in oxygen availability and other environmental factors (Zawierucha et al., 2021; Devetter et al., 2025). The presence of nematodes in SSPs, therefore, suggests that these habitats provide distinct environmental conditions that support taxonomically richer microfaunal communities.
Among the larger invertebrates recorded in the SSPs, collembolans and spiders were the most expected taxa, as they are typical inhabitants of winter and spring snow cover, e.g., in Fennoscandia (Hågvar, 2010). However, in the present study, the dominant arthropods were mites, followed by insects. The lack of information about mites from snow could be related to their small size and small number of experts on the diversity and ecology of this diverse arthropod's group. Mites are ubiquitous in terrestrial ecosystems; however, they are not common in cryospheric environments and their role on snow requires more studies in future. Other groups, such as Chilopoda, Diplura, Opiliones and Protura, were present in smaller numbers. This unusual taxonomic structure of arthropods in SSPs may reflect the specific characteristics of the studied habitats, particularly the much shorter duration of seasonal snow cover and the lower elevation compared with Fennoscandia. One of the most remarkable findings among the larger invertebrates was the presence of Diptera larvae, indicating that at least some insect species may be capable of reproduction under these environmental conditions. Chironomids (Diamesinae and others) represent one of the most widespread groups of insects inhabiting snow surfaces (Hågvar and Krzemińska, 2008; Hågvar, 2010, and others), including in lowlands (Soszyńska-Maj et al., 2016). However, in the SSPs, representatives of the orders Coleoptera, Hymenoptera, and Hemiptera were neither less frequent nor less abundant than non-biting midges. In contrast, Mecoptera, Plecoptera and Trichoptera, taxa commonly reported from the snow habitats (Bouchard et al., 2009; Hågvar, 2010; Soszyńska-Maj et al., 2016), were not found. Since many specimens were destroyed or in poor condition, our results on adult flying insects in SSPs should be regarded as preliminary, likely reflecting occasional occurrences rather than stable snow-associated assemblages. Similarly, other taxa of larger invertebrate taxa recorded in SSPs in low numbers (diplurans, proturans, spiders, harvestmen) may represent fortuitous findings. In addition, some insects and other arthropods may have been underrepresented due to their ability to actively avoid sampling by leaving the area in response to human approach. In contrast, the more abundant and widely represented small metazoans (rotifers, tardigrades, nematodes and mites) can be considered as true snow inhabitants in European lowlands. Considering their numbers and different instars, some of these animals are adapted to snow conditions and benefit from them.
The very short persistence of SSPs appears inconsistent with the high abundances that these SSP-dwelling invertebrates can reach. As shown in the results, total microinvertebrate densities exceeded 1000 ind m−2 in several of the snow patches examined. Comparing these numbers, expressed per litre values, with the densities of invertebrates from ice or snow surfaces gives controversial results. In the snow in the mountains of Japan (Ono et al., 2021), the density of both tardigrades and rotifers exceeded those in SSPs by an order of magnitude, but only within visibly coloured patches associated with algal blooms. In contrast, “clean” snow from the same region contained tardigrade densities comparable to those found in SSPs and even lower numbers of rotifers. Similarly, rotifers, tardigrades, and nematodes were more abundant in studied SSPs (reaching more than 400, 100, and 170 ind L−1, respectively) than on glaciers, where maximum reported values are approximately 40 ind L−1 for tardigrades and 3–4 ind L−1 for both rotifers and nematodes (Shain et al., 2021). Densities of bdelloid rotifers recorded on the ice surface of polar glaciers in Iceland (Shain et al., 2016) were also lower (1–100 ind L−1) than in SSPs. In Arctic cryoconite densities of tardigrades in cryoconite holes reach 82 ind cm−2 (820 000 ind m−2), however cryoconite is characterised by lower number of competitors (nematodes, mites) and higher biomass of potential food, i.e., algae and cyanobacteria (Jaroměřská et al., 2025). In the Antarctic, permanent snow described as “water-melon snow”, contained large densities of pigmentated rotifer individuals (Murray, 1910). However, this work did not include quantitative data or an explanation of the phenomenon.
In this regard, the microfauna in SSPs can be viewed as a cryophilic community in a transitional state: more developed, diverse, and abundant than those in many glacial systems, yet less abundant than communities in snow with algal blooms. This intermediate status is likely arising from the specific conditions of seasonal snow patches, which lack pronounced algal blooms but are nevertheless suitable for living. Microalgae belonging to Chloromonas and cyanobacteria belonging to Chroococcus were found at each site of early spring snow. First taxon was more abundant in SSPs; however, its numbers were much lower than in mountain snow across the world (e.g., Procházková et al., 2020). Unlike rotifers and tardigrades in the long-living seasonal snow of Japanese mountains (Ono et al., 2021, 2025), both taxa in SSPs showed little dependence on microalgal or cyanobacterial cell concentrations, exhibiting only weak positive correlations with cyanobacteria and no correlation with green algae (Fig. 4). Notably, high numbers of microinvertebrates in SSPs were recorded in the upper snow layer, whereas Ono et al. (2025) reported increasing densities of both rotifers and tardigrades in the subsurface layer of snowpacks in the mountains of Japan. The latter, however, represents a system fundamentally different from SSPs due to its larger thickness and longer persistence – until late spring, which at these latitudes (around 44 N) is characterised by high solar radiation (up to 800 W m−2) and ambient temperature exceeding 20 °C. Under such conditions, microinvertebrates and all motile microbes tend to migrate 10–20 cm below the surface during the day (Ono and Takeuchi, 2025). In contrast, all SSPs sampled in the Baltic states did not reach sufficient thickness to allow investigation of microfaunal densities across different layers. Moreover, they disappear much earlier in the season and hence not exposed to intense solar radiation. This suggests that microfaunal densities observed in the surface layer of SSPs provide a representative estimate of their distribution.
In an attempt to explain the observed densities of invertebrates under the conditions without algal blooms, we focused on the surrounding landscape. Thus, we observed a slight tendency for invertebrate abundances to increase in more shaded and sheltered locations, e.g., more forested areas (Fig. 5). The surrounding extensive and dense vegetation, ranging from shrubs to evergreen conifers with large canopies, appears to be another major environmental characteristic that distinguishes SSPs from the cryohabitats in polar and alpine snow and ice. Ono et al. (2025) highlighted the role of trees as a source of nutrients essential for primary producers in underlaying snow ecosystems. Our results extend this concept to snow ecosystems lacking algal blooms, indicating that invertebrate communities can develop using an alternative source of organic matter: allochthonous inputs derived from surrounding vegetation. As identified in ordination analysis (Fig. 6), the canopy cover, or the degree of shadiness of the area, influences on the invertebrate distribution, but tree density and proximity of tree trunks do not. These results suggest that the microfauna in SSPs is shaped predominantly by the vertical structure of the surrounding landscape rather than by the horizontal structure, meaning the primary role of precipitation-like inputs of organic and mineral material onto the snow surface. Moreover, the most affecting variables are likely linked to processes of deposition from the canopies. For example, Chl a showed only a weak relationship with green algae or cyanobacteria, neither of which contributed to the variation observed in snow-dwelling invertebrates. This suggests that the chlorophyll quantified after extraction from snow samples likely originates from microscopic plant fragments rather than active algal growth, supporting the importance of external inputs from the above-growing vegetation. Fine organic or mineral particles, which are also positively associated with invertebrate distribution (see PP, TSS, and conductivity variables in Fig. 6), originate from the close surrounding and – obviously to a lesser extent – due to atmospheric inputs. Another important variable, organic debris (needles, etc.) obviously emerges from trees growing above and accumulates on the snow surface. Seasonal snow in lowland forested landscapes may therefore function as boundary cryohabitats influenced by surrounding vegetation, where invertebrate communities are sustained primarily by externally derived organic inputs rather than by in situ primary production. Consequently, invertebrates on SSPs can feed on diverse food sources, including microalgae, but more likely on deposited organic particles, and heterotrophic bacteria and fungi, which decompose organic matter. Indeed, fungi and heterotrophic bacteria are part of the tardigrade diet on glaciers (Zawierucha et al., 2022).
Regarding the origin of microfauna itself on the snow surface, two main scenarios can be considered. First, individuals may arrive in SSPs as part of this depositional process, being passively transferred from surrounding vegetation together with plant particles and organic debris. Second, microfauna may settle in the snow patches via longer-distance dispersal, although their establishment would still depend on the organic inputs from the surrounding vegetation, first of all, from tree canopies above the snow. A combination of both mechanisms can occur as different taxa show slightly different correlations with environmental variables. Tardigrades, which are more strongly related to the particulate fraction in snow, and rotifers, which correlate with canopy cover, may primarily originate from surrounding trees. In contrast, nematodes, which are less related to these variables, may arrive via alternative dispersal mechanisms. Additionally, we cannot exclude the possibility of some invertebrates migrated from the soil beneath the snow, as is known for annelids (Dial et al., 2012), but less possible for microfauna, main animals on SSPs. To address these questions, more targeted future research is required, including the identification of species from both snow patches and surrounding habitats using combined morphological and molecular approaches.
Seasonal snow patches in European lowlands represent short-living and fragile cryoecosystems, nevertheless, they are capable of hosting diverse communities of invertebrates. Microfauna (e.g., bdelloid rotifers, tardigrades, and nematodes) were the most widespread and abundant metazoans on snow, followed by mites, and likely use the snow environment as a site for reproduction. Other invertebrates of larger body size (e.g., insects and springtails) were recorded only sporadically and in low numbers. The densities and diversity of invertebrates on SSPs are strongly driven by variables that are directly derived from the surroundings (trees and their canopies above the snow). Particularly, communities are shaped by Chl a likely originating from plant fragments, as well as by inorganic and organic particles, and fallen organic debris.
The results of the study support the view that SSPs represent a very specific type of cryoecosystem. Microfaunal communities in cryoconite holes on glaciers are primarily structured by the accumulation of dark-coloured particles that reduce surface albedo and by the presence of algae and bacteria, which serve as a primary food source for both rotifers and tardigrades. In contrast, multicellular life in seasonal snow in lowlands critically depends on the immediate surroundings rather than on wind-blown material from distant sources, and on externally derived organic matter rather than on autotrophic prokaryotes. The availability of a film of liquid water on the snow surface appears to be the only driver relevant across all types of cryosphere ecosystems, including SSPs. Considering the high abundance of invertebrates on SSPs and the substantial accumulation of organic matter on their surface over time, SSPs are important not only as habitats but also as a source seeding the ground immediately after snowmelt.
The source data on which the conclusions of this work are based are available within the article and Tables S1–S3 in the Supplement.
The online version contains supplementary material: Tables S1–S3. The supplement related to this article is available online at https://doi.org/10.5194/bg-23-4199-2026-supplement.
DL collected all samples, performed all statistical analyses, and prepared the text and the figures. Invertebrates were sorted and analysed by DL and KZ, and the research idea was developed by KZ and DL. JP conducted analyses of chemical parameters and nutrients in the snow samples, DV assessed pigment composition, DO processed all samples for microalgal analyses, and GK participated in sample preparation. MK provided overall supervision of the study. All authors contributed to writing the manuscript.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
This article is part of the special issue ”Cryospheric ecosystems: climate feedback loops, threatened ecosystems, and consequences of climate change”. It is not associated with a conference.
The authors are grateful to the Editor, Dr. Ewa Poniecka, Dr. Dorota L. Porazinska, and two anonymous reviewers for their valuable comments and suggestions, which substantially improved the manuscript.
This research has been supported by the Lithuanian Research Council (“Microfauna emerging in seasonal snow patch ecosystems – losers or winners? (SNOWLIFE)” (grant no. S-MIP-23-25)).
This paper was edited by Ewa Poniecka and reviewed by Dorota L. Porazinska and two anonymous referees.
Anesio, A. M. and Laybourn-Parry, J.: Glaciers and ice sheets as a biome, Trends Ecol. Evol., 27, 219–225, 2012.
Bertolani, R., Rebecchi, L., and Beccaccioli, G.: Dispersal of Ramazzottius and other tardigrades in relation to type of reproduction, Invertebr. Reprod. Dev., 18, 153–157, https://doi.org/10.1080/07924259.1990.9672137, 1990.
Bosson, J. B., Huss, M., Cauvy-Fraunié, S., Clément, J. C., Costes, G., Fischer, M., Poulenard, J., and Arthaud, F.: Future emergence of new ecosystems caused by glacial retreat, Nature, 620, 562–569, https://doi.org/10.1038/s41586-023-06302-2, 2023.
Bouchard Jr., R. W., Schuetz, B. E., Ferrington Jr., L. C., and Kells, S. A.: Cold hardiness in the adults of two winter stonefly species: Allocapnia granulata (Claassen, 1924) and A. pygmaea (Burmeister, 1839) (Plecoptera: Capniidae), Aquat. Insect., 31, 145–155, 2009.
Brauer, F.: Insecten-Leben im Winter, Vereine zur Verbreitung naturwissenschaftlicher Kenntnisse im Wien, 357–381, 1871.
Chapman, J. A.: Observations on snow insects in western Montana, Can. Entomol., 86, 357–363, 1954.
Crosta, A., Valle, B., Caccianiga, M., Gobbi, M., Ficetola, F. G., Pittino, F., Franzetti, A., Azzoni, R. S., Lencioni, V., Senese, A., and Corlatti, L.: Ecological interactions in glacier environments: a review of studies on a model Alpine glacier, Biol. Rev., 100, 227–244, 2025.
Devetter, M., Renčo, M., Háněl, L., Jimenez-Santos, M. A., Doležálková, M. K., and Janko, K.: Rare presence of nematodes in cryoconite holes: evidence from a norwegian glacier, Hydrobiologia, 853, 865–879, https://doi.org/10.1007/s10750-025-05971-6, 2025.
Dial, C. R., Dial, R. J., Saunders, R., Lang, S. A., Lee, B., Wimberger, P., Dinapoli, M. S., Egiazarov, A. S., Gipple, S. L., Maghirang, M. R., Swartley-McArdle, D. J., Yudkovitz S. R., and Shain, D. H.: Historical biogeography of the North American glacier ice worm, Mesenchytraeus solifugus (Annelida: Oligochaeta: Enchytraeidae), Mol. Phylogenet. Evol., 63, 577–584, 2012.
Dimattia, L. M., Saglam, N., Saunders, R., and Shain, D. H.: An ice inhabiting bdelloid rotifer from North America, Extremophiles, 29, 30, https://doi.org/10.1007/s00792-025-01390-6, 2025.
Domine, F.: Should we not further study the impact of microbial activity on snow and polar atmospheric chemistry?, Microorganisms, 7, 260, https://doi.org/10.3390/microorganisms7080260, 2019.
Dong, Z., Jiang, H., Baccolo, G., Di Mauro, B., and Zawierucha, K.: Biological and pollution aerosols on snow and ice: interplay between the atmosphere and the cryosphere, J. Earth Sci., 34, 1951–1956, https://doi.org/10.1007/s12583-023-2004-2, 2023.
European Committee for Standardization: Water quality – Guidance standard on the enumeration of phytoplankton using inverted microscopy (Utermöhl technique), EN 15204:2006, https://repository.oceanbestpractices.org/server9/api/core/bitstreams/827e1cd6-84f9-4771-a27d-815c95aedeed/content (last access: 20 October 2025), 2006.
European Environment Agency: Global and European temperatures, https://www.eea.europa.eu/en/analysis/indicators/global-and-european-temperatures, last access: 15 November 2025.
Federation, W. E. and Aph Association: Standard methods for the examination of water and wastewater, American Public Health Association (APHA), Washington, DC, USA, 21 pp., 2005.
Ferree, M. A. and Shannon, R. D.: Evaluation of a second derivative UV/visible spectroscopy technique for nitrate and total nitrogen analysis of wastewater samples, Water Res., 35, 327–332, 2001.
Fiołka, M. J., Takeuchi, N., Sofińska-Chmiel, W., Mieszawska, S., and Treska, I.: Morphological and physicochemical diversity of snow algae from Alaska, Sci. Rep.-UK, 10, 19167, https://doi.org/10.1038/s41598-020-76215-x, 2020.
Fontaneto, D.: Long-distance passive dispersal in microscopic aquatic animals, Mov. Ecol., 7, https://doi.org/10.1186/s40462-019-0155-7, 2019.
Fontaneto, D., Ficetola, G. F., Ambrosini, R., and Ricci, C.: Patterns of diversity in microscopic animals: are they comparable to those in protists or in larger animals?, Global Ecol. Biogeogr., 15, 153–162, https://doi.org/10.1111/j.1466-822X.2006.00193.x, 2006.
Fontaneto, D., Herniou, E. A., Barraclough, T. G., and Ricci. C.: On the global distribution of microscopic animals: New worldwide data on bdelloid rotifers, Zool. Stud., 46, 336–346, 2007.
Giannetti, D., Giovannini, I., Massa, E., Schifani, E., Rebecchi, L., Guidetti, R., and Grasso, D. A.: Dispersion and new shelters offered by ants: myrmecophoresy of tardigrades, Front. Zool., 22, 30, https://doi.org/10.1186/s12983-025-00581-3, 2025.
Gobbi, M. and Lencioni, V.: Glacial Biodiversity: Lessons from Ground-dwelling and Aquatic, Glaciers and the polar environment, 143, https://doi.org/10.5772/intechopen.92826, 2020.
Hågvar, S.: A review of Fennoscandian arthropods living on and in snow, Eur. J. Entomol., 107, 281–298, https://doi.org/10.14411/eje.2010.036, 2010.
Hågvar, S. and Krzemińska, E.: Contribution to the winter phenology of Trichoceridae (Diptera) in snow-covered southern Norway, Studia dipterologica, 14, 271–283, 2008.
Hanzelová, M., Vido, J., Škvarenina, J., Nalevanková, P., and Perháčová, Z.: Microorganisms in summer snow patches in selected high mountain ranges of Slovakia, Biologia, 73, 1177–1186, https://doi.org/10.2478/s11756-018-0136-0, 2018.
Harrell Jr., F. E.: Package “Hmisc” (Version 4.7-1), https://cran.r-project.org/web/packages/Hmisc/Hmisc.pdf (last access: 1 October 2025), 2022.
HELCOM: Guidelines for monitoring of phytoplankton species composition, abundance and biomass, HELCOM, Helsinki, Finland, 22 pp., https://doi.org/10.25607/OBP-1822, 2021.
Hodson, A., Anesio, A. M., Tranter, M., Fountain, A., Osborn, M., Priscu, J., Laybourn-Parry, J., and Sattler, B.: Glacial ecosystems, Ecol. Monogr., 78, 41–67, 2008.
Hodson, A. J., Mumford, P. N., Kohler, J., and Wynn, P. M.: The High Arctic glacial ecosystem: new insights from nutrient budgets, Biogeochemistry, 72, 233–256, 2005.
Hoham, R. W. and Remias, D.: Snow and glacial algae: a review, J. Phycol., 56, 264–282, https://doi.org/10.1111/jpy.12967, 2020.
Hotaling, S. Shain, D. H., Lang, S. A., Bagley, R. K., Tronstad, L. M., Weisrock, D. W., and Kelley, J. L.: Long-distance dispersal, ice sheet dynamics and mountaintop isolation underlie the genetic structure of glacier ice worms, P. Roy. Soc., 286, 20190983, https://doi.org/P10.1098/rspb.2019.0983, 2019.
Hotaling, S., Wimberger, P. H., Kelley, J. L., and Watts, H.: Macroinvertebrates on glaciers, Ecology, 101, 1–3, 2020.
International Organisation for Standardisation: Water quality. Determination of ammonium. Part 1: Manual spectrometric method, ISO 7150-1:1984, https://www.iso.org/standard/18782.html (last access: 12 November 2025), 1984.
International Organisation for Standardisation: Soil quality – Determination of organic and total carbon after dry combustion (“elemental analysis”), ISO 10694:1995, https://www.iso.org/standard/18782.html (last access: 10 November 2025), 1995.
International Organisation for Standardisation: Determination of nitrite. Molecular adsorption spectrometric method, ISOEN 26777:1993, https://standards.iteh.ai/catalog/standards/ cen/27c3cc18-7b44-4c2c-bd04-52ff233b7828/en-26777-1993? srsltid=AfmBOopvR1UlMOn5Lbp35GYEn9yHSb1sgM6zMts ECZ8wicu9y8eApW8T (last access: 31 October 2025), 1993.
International Organisation for Standardisation: Water quality – Determination of phosphorus – Ammonium molybdate spectrometric method, EN ISO 6878:2004, https://www.iso.org/standard/36917.html (last access: 15 October 2025), 2004.
Jaroměřská, T. N., Ambrosini, R., Richter, D., Pietryka, M., Niedzielski, P., Souza-Kasprzyk, J., Klimaszyk, P., Franzetti, A., Pittino, F., Vondrovicová, L., and Senese, A.: Insights Into Cryoconite Community Dynamics on the Alpine Glacier Throughout the Ablation Season, Ecol. Evol., 15, e71064, https://doi.org/10.1002/ece3.71064, 2025.
Jeffrey, S. W. and Humphrey, G. F.: New Spectrophotometric Equations for Determining Chlorophylls a, b, c1 and c2 in Higher Plants, Algae and Natural Phytoplankton, Biochem. Physiol. Pfl., 167, 191–194, https://doi.org/10.1016/S0015-3796(17)30778-3, 1975.
Kivinen, S., Kaarlejärvi, E., Jylhä, K., and Räisänen, J.: Spatiotemporal distribution of threatened high-latitude snowbed and snow patch habitats in warming climate, Environ. Res. Lett., 7, 034024, https://doi.org/10.1088/1748-9326/7/3/034024, 2012.
Krasting, J. P., Broccoli, A. J., Dixon, K. W., and Lanzante, J. R.: Future changes in Northern Hemisphere snowfall, J. Climate, 26, 7813–7828, 2013.
Leya, T.: Snow algae: adaptation strategies to survive on snow and ice, in: Polyextremophiles: Life under multiple forms of stress, edited by: Seckbach, J., Oren, A., and Stan-Lotter, H., Springer, Dordrecht, Netherlands, 401–423, https://doi.org/10.1007/978-94-007-6488-0_17, 2013.
Morek, W., Surmacz, B., López-López, A., and Michalczyk, Ł.: “Everything is not everywhere”: Time-calibrated phylogeography of the genus Milnesium (Tardigrada), Mol. Ecol., 30, 3590–3609, 2021.
Murray, J.: Antarctic Rotifera, British Antarctic Expedition 1907–9, 41–65, https://doi.org/10.5962/bhl.title.22427, 1910.
Oksanen, J., Blanchet, J. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., Minchin, P. R., O'Hara, R. B., Simpson, G. L., Solymos, P., Stevens, M. H. M., Szoecs, E., and Wagner, H.: Vegan: Community Ecology Package. R package version 2.5-7, https://CRAN.R-project.org/package=vegan (last access: 5 October 2025), 2020.
Olenina, I., Hajdu, S., Edler, L., Andersson, A., Wasmund, N., Busch, S., Gobel, J., Gromisz S., Huseby, S., Huttunen, M., Jaanus, A., Kokkonen, P., Jurgensone, I., and Niemkiewicz, E.: Biovolumes and size-classes of phytoplankton in the Baltic Sea, HELCOM Baltic Sea Environmental Proceedings, 106, 1–144, 2006.
Ono, M. and Takeuchi, N.: The diel vertical migration of microbes within snowpacks driven by solar radiation and nutrients, Arct. Antarct. Alp. Res., 57, 2460253, https://doi.org/10.1080/15230430.2025.2460253, 2025.
Ono, M., Takeuchi, N., and Zawierucha, K.: Snow algae blooms are beneficial for microinvertebrates assemblages (Tardigrada and Rotifera) on seasonal snow patches in Japan, Sci. Rep.-UK, 11, 5973, https://doi.org/10.1038/s41598-021-85462-5, 2021.
Ono, M., Takeuchi, N., and Zawierucha, K.: Description of a new species of Tardigrada Hypsibius nivalis sp. nov. and new phylogenetic line in Hypsibiidae from snow ecosystem in Japan, Sci. Rep.-UK, 12, 14995, https://doi.org/10.1038/s41598-022-19183-8, 2022.
Ono, M., Kobayashi, K., Seto, D., Konishi, F., Wada, K., Usuba, S., and Takeuchi, N.: Temporal and vertical changes in snow microbial communities during the melting season below canopy in Northern Japan, The Cryosphere, 19, 5983–5999, https://doi.org/10.5194/tc-19-5983-2025, 2025.
Pittino, F., Zawierucha, K., Poniecka, E., Buda, J., Rosatelli, A., Zordan, S., Azzoni, R. S., Diolaiuti, G., Ambrosini, R., and Franzetti, A.: Functional and taxonomic diversity of anaerobes in supraglacial microbial communities, Microbiology Spectrum, 11, e01004-22, https://doi.org/10.1128/spectrum.01004-22, 2023.
Procházková, L., Remias, D., Bilger, W., Křížková, H., Řezanka, T., and Nedbalová, L.: Cysts of the Snow Alga Chloromonas krienitzii (Chlorophyceae) Show Increased Tolerance to Ultraviolet Radiation and Elevated Visible Light, Front. Plant Sci., 11, 617250, https://doi.org/10.3389/fpls.2020.617250, 2020.
Procházková, L., Remias, D., Suzuki, H., Kociánová, M., and Nedbalová, L.: Chloromonas rubrosalmonea sp. nov. causes blooms of salmon-red snow due to high astaxanthin and low chlorophyll content, Bot. Lett., 171, 1–19, https://doi.org/10.1080/23818107.2023.2301608, 2024.
Ramazzotti, G. and Maucci, W.: Il Phylum Tardigrada, Mem. dell'Ist. Ital. di Idrobiol. Pallanza, 41, 1–1012, 1983.
Remias, D., Procházková, L., Nedbalová, L., Benning, L. G., and Lutz, S.: Novel insights in cryptic diversity of snow and glacier ice algae communities combining 18S rRNA gene and ITS2 amplicon sequencing, FEMS Microbiol. Ecol., 99, fiad134, https://doi.org/10.1093/femsec/fiad134, 2023.
Ricci, C.: Ecology of bdelloids: how to be successful, Hydrobiologia, 147, 117–127, https://doi.org/10.1007/BF00025734, 1987.
Robertson, M. W., Russo, N. J., McInnes, S. J., Goffinet, B., and Jiménez, J. E.: Potential dispersal of tardigrades by birds through endozoochory: evidence from Sub-Antarctic White-bellied Seedsnipe (Attagis malouinus), Polar. Biol., 43, 899–902, https://doi.org/10.1007/s00300-020-02680-9, 2020.
Rozwalak, P. Podkowa, P., Buda, J., Niedzielski, P., Kawecki, S., Ambrosini, R., Azzoni, R. S., Baccolo, G., Ceballos, J. L., Cook, J., and Di Mauro, B.: Cryoconite – From minerals and organic matter to bioengineered sediments on glacier's surfaces, Sci. Total Environ., 807, 150874, https://doi.org/10.1016/j.scitotenv.2021.150874, 2022.
Schill, R. O. (Ed.): Water bears: The biology of tardigrades, Springer Cham, Switzerland, 419 pp., ISBN 978-3-319-95701-2, 2018.
Segawa, T., Matsuzaki, R., Takeuchi, N., Akiyoshi, A., Navarro, F., Sugiyama, S., Yonezawa, T., and Mori, H.: Bipolar dispersal of red-snow algae, Nat. Commun., 9, 3094, https://doi.org/10.1038/s41467-018-05521-w, 2018.
Shain, D. H. Halldórsdóttir, K., Pálsson, F., Aðalgeirsdóttir, G., Gunnarsson, A., Jónsson, Þ., Lang, S. A., Pálsson, H. S., Steinþórssson, S., and Arnason, E.: Colonization of maritime glacier ice by bdelloid Rotifera, Mol. Phylogenet. Evol., 98, 280–287, 2016.
Shain, D. H., Novis, P. M., Cridge, A. G., Zawierucha, K., Geneva, A. J., and Dearden, P. K.: Five animal phyla in glacier ice reveal unprecedented biodiversity in New Zealand's southern Alps, Sci. Rep.-UK, 11, 3898, https://doi.org/10.1038/s41598-021-83256-3, 2021.
Shain, D. H., Rogozhina, I., Fontaneto, D., Nesje, A., Saglam, N., Bartlett, J., Zawierucha, K., Kielland, Ø. N., Dunshea, G., Arnason, E., and Rosvold, J.: Ice-inhabiting species of Bdelloidea Rotifera reveal a pre-Quaternary ancestry in the Arctic cryosphere, Biol. Letters, 20, 20230546, https://doi.org/10.1098/rsbl.2023.0546, 2024.
Shmakova, L. Malavin, S., Iakovenko, N., Vishnivetskaya, T., Shain, D., Plewka, M., and Rivkina, E.: A living bdelloid rotifer from 24 000-year-old Arctic permafrost, Curr. Biol., 31, R712–R713, 2021.
Soszyńska-Maj, A., Paasivirta, L., and Giłka, W.: Why on the snow? Winter emergence strategies of snow-active Chironomidae (Diptera) in Poland, Insect Sci., 23, 754–770, 2016.
Utermöhl, H.: Zur Vervollkommnung der quantitativen Phytoplankton-Methodik, Int. Assoc. Theor. Appl. Limnol., 9, 1–38, 1958.
Valle, B., Porco, D., Skarżyński, D., Frati, F., Caccianiga, M., Rodriguez-Prieto, A., Zeni, M., and Gobbi, M.: Alpine blooming of “snow fleas”: the importance of snow for Alpine springtails (Hexapoda: Collembola) ecology and biodiversity, Rend. Lincei-Sci. Fis., 35, 163–180, 2024.
Voigt, T., Füssel, H. M., Gärtner-Roer, I., Huggel, C., Marty, C., and Zemp, M.: Impacts of climate change on snow, ice, and permafrost in Europe: Observed trends, future projections, and socio-economic relevance, ETC/ACC Technical Paper, 13, 1–117, 2010.
Wickham, H.: Ggplot2: Elegant Graphics for Data Analysis. R package version 3.5.1, https://cran.r-project.org/web/packages/ggplot2/index.html (last access: 20 October 2025), 2016.
Yakimovich, K. M., Engstrom, C. B., and Quarmby, L. M.: Alpine snow algae microbiome diversity in the Coast Range of British Columbia, Front. Microbiol., 11, 1721, https://doi.org/10.3389/fmicb.2020.01721, 2020.
Zawierucha, K. and Shain, D. H.: Disappearing Kilimanjaro snow – Are we the last generation to explore equatorial glacier biodiversity?, Ecol. Evol., 9, 8911–8918, 2019.
Zawierucha, K., Kolicka, M., Takeuchi, N., and Kaczmarek, Ł.: What animals can live in cryoconite holes? A faunal review, J. Zool., 295, 159–169, 2015.
Zawierucha, K. Buda, J., Pietryka, M., Richter, D., Łokas, E., Lehmann-Konera, S., Makowska, N., and Bogdziewicz, M.: Snapshot of micro-animals and associated biotic and abiotic environmental variables on the edge of the south-west Greenland ice sheet, Limnology, 19, 141–150, 2018.
Zawierucha, K., Buda, J., Azzoni, R. S., Niśkiewicz, M., Franzetti, A., and Ambrosini, R.: Water bears dominated cryoconite hole ecosystems: densities, habitat preferences and physiological adaptations of Tardigrada on an alpine glacier, Aquat. Ecol., 53, 543–556, https://doi.org/10.1007/s10452-019-09707-2, 2019.
Zawierucha, K., Porazinska, D. L., Ficetola, G. F., Ambrosini, R., Baccolo, G., Buda, J., Ceballos, J. L., Devetter, M., Dial, R., Franzetti, A., and Fuglewicz, U.: A hole in the nematosphere: tardigrades and rotifers dominate the cryoconite hole environment, whereas nematodes are missing, J. Zool., 313, 18–36, 2021.
Zawierucha, K. Trzebny, A., Buda, J., Bagshaw, E., Franzetti, A., Dabert, M., and Ambrosini, R.: Trophic and symbiotic links between obligate-glacier water bears (Tardigrada) and cryoconite microorganisms, PLoS One, 17, e0262039, https://doi.org/10.1371/journal.pone.0262039, 2022.
Zawierucha, K., Kašparová, E. Š., McInnes, S., Buda, J., Ambrosini, R., Devetter, M., Ficetola, G. F., Franzetti, A., Takeuchi, N., Horna, P., and Jaroměřská, T. N.: Cryophilic Tardigrada have disjunct and bipolar distribution and establish long-term stable, low-density demes, Polar Biol., 46, 1011–1027, 2023a.
Zawierucha, K., Stec, D., Dearden, P. K., and Shain, D. H.: Two new tardigrade genera from New Zealand's Southern Alp glaciers display morphological stasis and parallel evolution, Mol. Phylogenet. Evol., 178, 107634, https://doi.org/10.1016/j.ympev.2022.107634, 2023b.