the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 08 Jul 2026
| 08 Jul 2026
Ideas and perspectives: Nitrite turnover controls nitrogen fate across redox gradients
Mathieu Sebilo
Rosanna Margalef-Marti
Reactive nitrogen fate in natural systems remains difficult to predict because pathway partitioning occurs at the stage of nitrite turnover, where rapid and tightly coupled production and consumption processes obscure the underlying fluxes. Concentration-based assessments emphasize the dominant pools – nitrate and ammonium – while pathway divergence is determined at the stage of nitrite turnover, independently of pool size. Nitrite is the principal dissolved inorganic intermediate linking the oxidative and reductive branches of the nitrogen cycle and the obligatory precursor to all downstream dissolved and gaseous products. Because nitrite rarely accumulates, it has often been treated as a transient intermediate of limited interpretive value. This apparent invisibility reflects rapid, tightly coupled turnover and does not indicate functional insignificance. Its low and frequently undetectable concentration is the kinetic signature of this central position rather than evidence against it: rapid coupled turnover sustains high gross flux at near-zero standing concentration. Nitrogen retention, recycling and losses to the atmosphere are determined during nitrite turnover, where competing pathways partition fluxes according to kinetic and environmental constraints.
Observed concentrations integrate formation and consumption into a net signal that masks opposing fluxes when internal cycling is rapid. Coupled δ15N–δ18O measurements of nitrite constrain simultaneous production and consumption and differentiate biological from abiotic pathways. Partial oxygen isotope exchange with water increases the diagnostic primacy of δ15N in resolving hidden turnover. However, its low concentration in natural environments can pose some challenges for analysis, requiring more sensitive approaches.
Centering nitrogen-cycle interpretation on nitrite dynamics and isotopic expression across redox gradients from oxic soils to oxygen minimum zones, provides a mechanistic basis for predicting nitrogen budgets, N2O emissions, and ecosystem sensitivity to increasing redox variability under climate change and land-use intensification.
- Article
(1886 KB) - Full-text XML
- BibTeX
- EndNote




The nitrogen cycle regulates ecosystem productivity, water quality and climate. Anthropogenic perturbations have profoundly altered nitrogen fluxes across terrestrial and aquatic systems, leading to nitrate (NO) contamination, eutrophication, and enhanced emissions of nitrous oxide (N2O), a potent greenhouse gas and ozone-depleting substance (Vitousek et al., 1997; Galloway et al., 2004; Gruber and Galloway, 2008). Uncertainty persists regarding the mechanisms that determine whether reactive nitrogen is retained within ecosystems or transferred to the atmosphere.
Conceptual representations of the nitrogen cycle have traditionally emphasized stable pools – ammonium (NH) and nitrate (NO) – providing a coherent basis for large-scale budget analyses (Canfield et al., 2010). However, the location within the reaction sequence at which pathway divergence is determined remains unresolved.
In reaction networks governed by rapid kinetics and tight coupling, the apparent invisibility of intermediates reflects rapid turnover rather than limited functional relevance. Nitrite (NO) exemplifies this conceptual blind spot. Described as an ephemeral intermediate that does not accumulate under steady-state conditions (Heil et al., 2016; Wrage et al., 2001), it is often omitted from conceptual frameworks or treated implicitly. This assumption is not consistent with the structure of the nitrogen cycle reaction network.
The central role of nitrite follows from its position in the reaction network. It occupies the +III oxidation state, between ammonium and nitrate, and every transformation linking oxidative and reductive branches passes through it (Burgin and Hamilton, 2007; Lam and Kuypers, 2011; Kraft et al., 2011). Other intermediates, such as nitric oxide (NO) and N2O, also participate in both branches, but their lower stability in aqueous systems leaves nitrite as the only dissolved intermediate readily accessible to direct observation. No other inorganic nitrogen compound combines this dual role of convergence and divergence. The fate of nitrogen at this junction is governed by the balance between nitrite-producing and nitrite-consuming processes under specific redox, kinetic and environmental constraints (Firestone and Davidson, 1989; Denk et al., 2017; Deb et al., 2024).
Concentration measurements of nitrate, ammonium and gaseous products integrate multiple processes, masking mechanistic controls on pathway partitioning: similar nitrate concentrations can arise from fundamentally different combinations of nitrification, denitrification and DNRA (e.g. in riparian zones where both processes operate simultaneously), leading to divergent outcomes in nitrogen retention and N2O emissions.
Stable isotope measurements provide direct constraints on these hidden dynamics. Nitrite integrates oxidative and reductive fluxes; its isotopic composition records concurrent production and consumption even when net concentrations remain unchanged, though disentangling these overlapping signals requires the dual δ15N–δ18O approach descrived in detail in the Isotopic constraints section below. Available methods advances permit direct determination of natural-abundance δ15N and δ18O in nitrite (McIlvin and Altabet, 2005; Sebilo et al., 2019; Deb and Lewicka-Szczebak, 2025; Hu et al., 2026). Oxygen isotopes of nitrite are partially reset due to equilibrium with oxygen isotopes of ambient water, modifying the primary biological signal and thereby increasing the diagnostic importance of δ15N.
Resolving nitrogen fate requires shifting analytical focus from accumulated pools to the intermediate at which pathway divergence is decided. The reaction network structure, kinetics of nitrite turnover, and stable isotope constraints together provide a mechanistic basis for predicting nitrogen budgets and N2O emissions. This basis relies on considering nitrite as a central component of the nitrogen cycle rather than a transient residual.
The central role of nitrite emerges directly from the reaction network architecture (Fig. 1). Nitrite does not constitute a regulatory control point in a biochemical sense, but rather the reaction network node at which pathway partitioning is resolved. Under oxic conditions, nitrite is produced during ammonium oxidation by ammonia-oxidizing bacteria and archaea and subsequently oxidized to nitrate, or diverted into alternative pathways depending on oxygen availability and enzyme kinetics (Casciotti, 2016). In comammox organisms, both oxidation steps occur within a single metabolic framework (Daims et al., 2015; van Kessel et al., 2015), further constraining accumulation while maintaining high gross turnover.
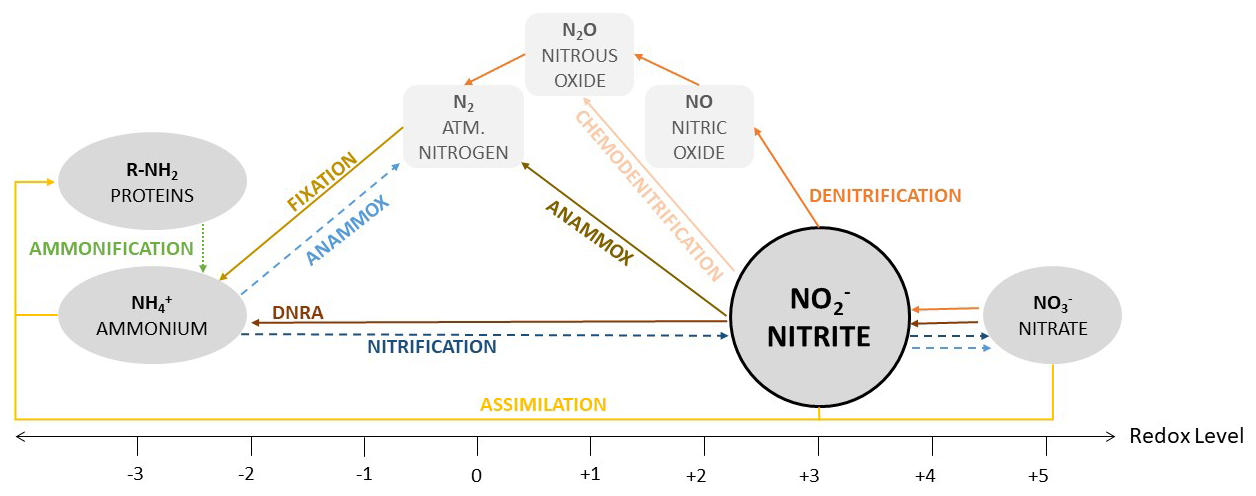
Figure 1Nitrite-based nitrogen cycling reactions across redox gradients. Oxidative pathways are presented in blue arrows, reductive ones in orange/brown and those involving organic matter in yellow/green. To simplify the scheme, hydroxylamine has not been included in the figure. Hydroxylamine is also involved in oxidative and reductive pathways. It can be produced during ammonia oxidation and nitrite reduction. It is primarily oxidized by nitrifying microorganisms to nitrite, with minor side production of nitric oxide and nitrous oxide, and it can be reduced to ammonium. However, it is highly reactive, it is usually rapidly oxidized before accumulating in the environment, limiting its influence at the ecosystem scale.
Under oxygen-limited or anoxic conditions, nitrite is produced during nitrate reduction and partitioned among denitrification, DNRA, anammox, and abiotic reduction. Each pathway channels nitrite toward distinct fates, from nitrogen retention to irreversible gaseous loss (Zumft, 1997; Lam and Kuypers, 2011). Abiotic reactions extend this network in both redox directions: reduction of nitrite by ferrous iron generates NO and N2O independently of enzymatic control, competing with biological pathways (Jones et al., 2015; Grabb et al., 2017; Robinson et al., 2021), and oxidation of nitrite to nitrate by ligand-bound Mn(III) proceeds abiotically under both oxic and anoxic conditions (Karolewski et al., 2021; Luther et al., 2021).
The defining characteristic of this convergence-divergence architecture is redistribution, not accumulation. Nitrite operates as a flux junction, a node at which inputs from multiple upstream pathways are portioned among competing downstream transformations, so that its concentration reflects the balance between upstream formation and downstream consumption, while its turnover rate determines the direction and magnitude of nitrogen transfer. In spatially heterogeneous environments, nitrite produced in one microdomain may be consumed in an adjacent zone within short diffusion distances (Firestone and Davidson, 1989). Accumulation arises primarily when this coupling is disrupted by kinetic limitation, redox fluctuation, or imbalance in electron donor and acceptor supply.
If the structural position of nitrite determines where pathway divergence occurs, the kinetic balance between gross production and gross consumption determines how. The direction and magnitude of nitrogen redistribution at the nitrite node depend on the kinetic balance between gross production and gross consumption. When these proceed at comparable rates, net concentration change approaches zero while flux through the intermediate remains substantial; pool size therefore provides limited information about pathway activity (Margalef-Marti et al., 2026). Such tightly coupled nitrite turnover has recently been observed in nitrate-rich groundwater systems using combined isotope and microbial approaches (Deb et al., 2025). A distinct regime arises when consumption is limited only by the rate of supply: nitrite is consumed as rapidly as it is produced, so that the standing concentration itself – not merely its rate of change – approaches zero even as gross flux remains intense. This is the limiting case in which an active and decisive node becomes analytically invisible.
Accumulation and persistence along time of nitrite is determined by the degree of kinetic coupling between sequential reactions. Tight coupling in oxic systems constrains accumulation and shortens residence time; disruption by fluctuating oxygen supply, transport limitation, or electron donor imbalance prolongs residence time and increases the probability of accumulation (see Sensitivity across environmental gradients section). Shifts in environmental conditions reorganize flux distribution at the nitrite node even when nitrate or ammonium pools exhibit minimal change.
Gaseous nitrogen (i.e., NO, N2O and N2) production represents a relevant downstream outcome of nitrite turnover. In microbial denitrification, the relative gross rates of successive reductions from nitrite to NO, N2O, and ultimately N2, determine the N2O : N2 ratio. Sustained electron donor supply (such as organic matter) and active nitrous oxide reductase favour completion of the sequence and N2 dominance; partial decoupling enhances expression of intermediate products, particularly N2O (Firestone and Davidson, 1989; Zumft, 1997; Lewicka-Szczebak et al., 2020). During oxygen-limited nitrification, nitrite generated from ammonium oxidation may be partially reduced within ammonia oxidizers, producing N2O under fluctuating oxygen conditions (Wrage et al., 2001). Abiotic reduction of nitrite by ferrous iron and reduced mineral phases generates NO and N2O independently of enzymatic control (Jones et al., 2015; Grabb et al., 2017).
Although N2O is not a primary product of canonical anammox metabolism – hydrazine oxidation yields N2 directly – anammox competes with denitrifiers for the available nitrite pool, thereby influencing the partitioning of nitrite between N2-producing and N2O-producing pathways in mixed-metabolism environments (Kartal et al., 2011). Therefore, the N2O : N2 ratio, and the relative production of N2 versus N2O more broadly, is controlled by turnover intensity and kinetic coupling at the nitrite stage rather than by pool size.
Stable isotope measurements of nitrite provide direct constraints on nitrogen transformations at the stage where flux redistribution occurs: the nitrite node. Isotopic composition responds to gross production and gross consumption rather than to net pool change alone. Constant concentration does not imply constant δ15N: progressive isotopic shifts may occur under steady-state pool conditions, revealing turnover intensity undetectable in concentration data (Fig. 2).
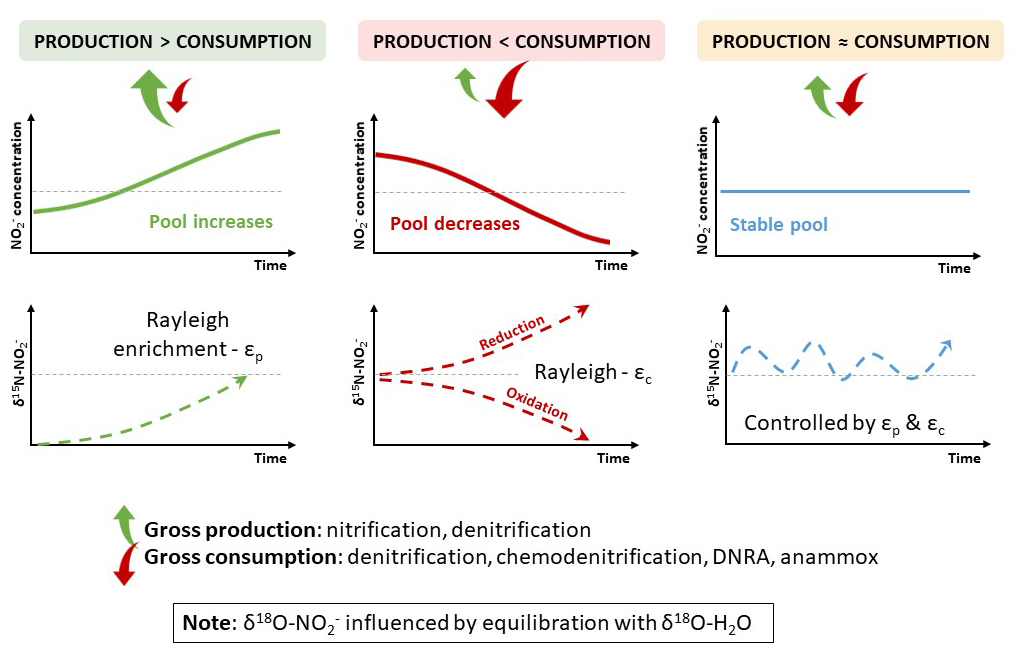
Figure 2Isotopic expression of gross nitrite turnover under contrasting production–consumption regimes. Top panels show nitrite concentration ([NO]) as a function of time, reflecting the net balance between gross production (P) and gross consumption (C): accumulation when P>C, depletion when P<C, and quasi-steady state when P≈C despite high internal fluxes. The P≈C regime spans a continuum of standing concentrations: where production and consumption balance independently, nitrite stabilises at a moderate level; where consumption is limited only by the rate of supply, nitrite is consumed as fast as it forms and its concentration approaches zero despite intense gross flux. Across this continuum, δ15N(NO) continues to evolve in response to gross turnover, so that the isotopic signal persists even where the pool itself becomes analytically invisible. Bottom panels illustrate conceptual trajectories of δ15N(NO). In contrast to concentration, isotopic composition responds to gross turnover rather than net change. δ15N(NO) evolves according to the isotopic signatures of contributing sources and the combined effects of isotope fractionation during production (εP) and consumption (εC). The direction of isotopic change is therefore process-dependent and not universal (e.g. inverse isotope effects during nitrite production by nitrification). Under conditions of balanced production and consumption (P≈C), δ15N(NO) may drift despite constant concentration, reflecting ongoing gross turnover. Oxygen isotope signals (δ18O) may be partially overprinted by exchange with water, increasing the diagnostic primacy of δ15N.
Each pathway imposes a characteristic kinetic isotope fractionation. During ammonia oxidation, inverse nitrogen isotope effects may enrich newly formed nitrite relative to its ammonium source (Casciotti et al., 2003; Santoro and Casciotti, 2011). During nitrite oxidation to nitrate, preferential removal of lighter isotopes enriches the residual pool in both 15N and 18O (Casciotti, 2009; Buchwald et al., 2012). Reductive pathways generate distinct fractionation patterns associated with nitrite reductase activity (Brunner and Bernasconi, 2005; Casciotti et al., 2010). Abiotic reduction by ferrous iron produces additional nitrogen isotope effects governed by surface-mediated electron transfer (Jones et al., 2015; Grabb et al., 2017). Abiotic oxidation of nitrite to nitrate by ligand-bound Mn(III) imparts a large inverse nitrogen isotope effect (20 ‰) closely resembling that of biological nitrite oxidation, with the additional oxygen atom derived from water (Karolewski et al., 2021). This convergence of biotic and abiotic fractionation factors reinforces the value of interpreting isotopic signatures within environmental and redox context. Overlapping fractionation factors across biological and abiotic pathways preclude simple end-member mixing and require integration with environmental and redox context. The direction of δ15N-NO evolution is not universal and depends on the relative contributions of concurrent production and consumption pathways, their associated isotope effects (εP, εC), and the isotopic composition of the source substrate.
Oxygen isotopes introduce an additional dimension. Exchange between nitrite oxygen atoms and ambient water may proceed on timescales comparable to biological turnover (Buchwald and Casciotti, 2010; Casciotti et al., 2010; Granger and Wankel, 2016), attenuating the independence of δ18O as a pathway tracer. Nitrogen isotopes do not undergo analogous exchange, preserving sensitivity to pathway-specific fractionation. Under conditions of significant oxygen isotope exchange, δ15N provides the more robust constraint on gross turnover.
Isotopic signatures propagate to gaseous products. The bulk δ15N of N2O reflects fractionation associated with nitrite reductases and kinetic coupling among successive reduction steps. Intramolecular 15N site preference (SP) provides additional mechanistic resolution: because SP is largely independent of the isotopic composition of precursor nitrite, it records enzyme-specific reaction pathways involved in N2O formation (Brunner and Bernasconi, 2005; Toyoda et al., 2017). Variations in SP discriminate among nitrifier-denitrification, canonical denitrification, and partial reduction sequences downstream of nitrite. Isotopomer signatures must account for subsequent N2O reduction and mixing, which may overprint primary fractionation signals (Toyoda et al., 2017; Lewicka-Szczebak et al., 2020).
Despite their strong mechanistic potential, nitrite isotope measurements are only feasible when sufficient nitrite accumulates for analysis and remain analytically challenging. Nitrite is chemically unstable and may undergo rapid oxidation or reduction during sampling and storage, potentially altering its isotopic composition before analysis. Reliable preservation therefore requires rapid processing and carefully controlled storage conditions, often under alkaline conditions that minimize transformation rates. In many terrestrial and aquatic systems, low nitrite concentrations restrict isotope analyses to zones where nitrite transiently accumulates. This emphasizes the need for continued development of high-sensitivity analytical approaches (Lewicka-Szczebak et al., 2021; Deb and Lewicka-Szczebak, 2025). For instance, a recent study developed a method for freshwater samples by coupling anion-exchange resin preconcentration with the azide reduction method for nitrite isotopic characterization at concentrations of 0.02 µmol L−1 or higher (Jiang et al., 2026). Tracer additions and molecular or transcriptomic measurements characterise the potential for nitrite turnover and necessarily perturb the system they interrogate. Natural-abundance isotope analysis of an in situ sample is alone non-perturbative: it records the actual expression of turnover under real conditions. The complementary methods therefore address a different question and cannot substitute for the in situ signal. The principal analytical challenge is ultimately not detection of the nitrite pool but deconvolution of a composite in situ signature integrating concurrent production and consumption with their respective fractionation factors.
Environmental systems differ not in whether nitrite turnover operates, but in how sensitively flux redistribution at the nitrite node responds to perturbation. Sensitivity is highest where oxidative and reductive metabolisms overlap at oxic-anoxic transition zones, and attenuates where reaction coupling is constrained to a single redox regime. Small shifts in oxygen supply, electron donor availability, or hydrological residence time can reorganize pathway partitioning at the nitrite node without producing detectable changes in bulk nitrogen pools. The nitrogen cycle is therefore most vulnerable to mechanistic misinterpretation precisely where nitrite turnover is most intense.
In fully oxic environments, nitrite is produced during ammonia oxidation and rapidly oxidized to nitrate, reflecting tight coupling between nitrification steps (Casciotti, 2016). Redox transition zones – oxic–anoxic interfaces in soils, sediments, riparian zones and stratified water columns – represent the most dynamic settings (Burgin et al., 2011). Simultaneous production from ammonia oxidation and nitrate reduction, combined with kinetically constrained consumption, promotes nitrite accumulation and amplifies sensitivity to environmental change (Buchwald and Casciotti, 2010; Bristow et al., 2016). These zones are recognized as hotspots of N2O production, consistent with the central role of nitrite in regulating the N2O : N2 ratio (Butterbach-Bahl et al., 2013; Babbin et al., 2020). In persistently anoxic environments – deep sediments, saturated soils and oxygen minimum zones – nitrite is predominantly generated via nitrate reduction and consumed through denitrification, DNRA or anammox (Zumft, 1997; Lam and Kuypers, 2011; Ward et al., 2009; Dalsgaard et al., 2012; Kalvelage et al., 2013; Denk et al., 2017; Deb et al., 2024). Hydrological and transport processes further modulate this coupling across all settings: long residence times favor complete turnover, whereas rapid transport can decouple production from consumption, allowing accumulation or downstream export (Sebilo et al., 2006).
Nitrogen cycling is commonly interpreted through the distribution of dominant inorganic pools. This perspective captures accumulation and export, it does not identify where transformation trajectories are decided. The reaction network places that decision at the stage of nitrite turnover. Environmental conditions regulate this distribution by modulating the rates of nitrite production and consumption.
This reframing resolves several persistent ambiguities. Similar nitrate or ammonium concentrations can arise from fundamentally different internal configurations of production and consumption. N2O emissions cannot be predicted from pool size alone because their magnitude depends on the completeness of reduction downstream of nitrite. Redox transition zones emerge as hotspots not because they contain larger pools, but because they intensify flux redistribution at this intermediate. A nitrite-centred framework shifts emphasis from storage to flux (Fig. 3), from accumulation to coupling, and from static pools to dynamic turnover, linking microbial metabolism, abiotic reactivity, redox heterogeneity, and atmospheric exchange within a single mechanistic perspective. Because both production and reduction of N2O depend on nitrite availability and turnover, resolving dynamics at this node provides a direct mechanistic link between microbial processes and climate-relevant gas fluxes.
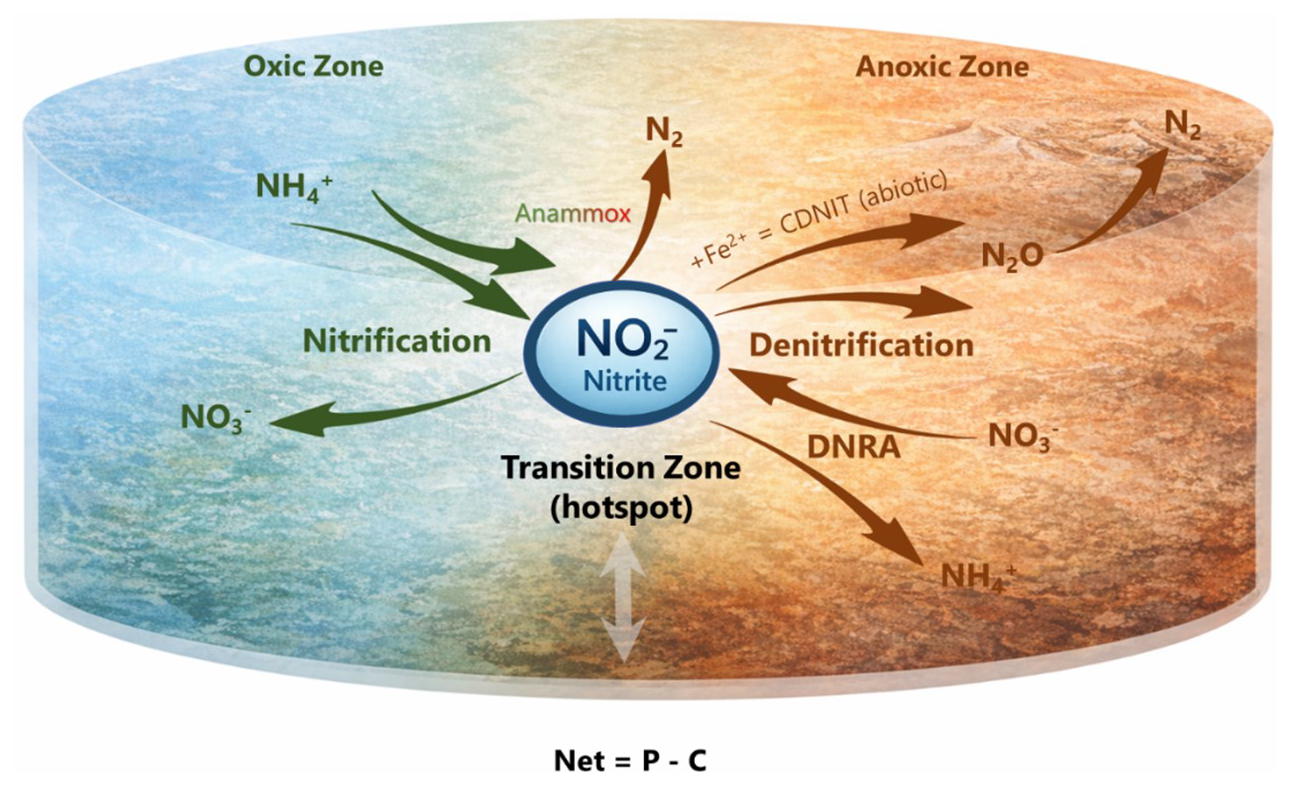
Figure 3Conceptual synthesis positioning nitrite turnover as the control point linking redox gradients, gross flux redistribution, isotopic expression, and nitrogen fate. Nitrite integrates oxidative and reductive processes across environmental gradients. Gross production and gross consumption determine residence time and flux partitioning among retention, recycling, and gaseous loss. The δ15N of NO records turnover intensity, whereas the δ18O may be modified by exchange with water. Isotopic signals propagate to N2O and inform pathway attribution. Nitrogen fate is therefore governed at the nitrite stage rather than by the size of accumulated nitrate or ammonium pools.
Changes in concentration reflectthe net balance between production and consumption. δ15N records the imprint of concurrent production and consumption even when pool size remains constant. Oxygen isotope exchange constrains interpretation but reinforces the diagnostic primacy of nitrogen isotopes. At the same time, the extent of oxygen exchange with water may provide information on nitrite residence times and the relative rates of biological turnover versus abiotic exchange (Buchwald and Casciotti, 2013; Lewicka-Szczebak et al., 2021). Incorporating nitrite isotopes converts an otherwise unobservable internal turnover into a measurable quantity.
Translating this framework into practice requires prioritising direct nitrite isotope measurements alongside conventional concentration measurements, particularly in redox-dynamic systems where net fluxes are most ambiguous. Dual δ15N–δ18O approaches, combined with isotopomer analysis of N2O, offer the most diagnostic power. Although nitrite isotope measurements offer strong mechanistic insight, their application is constrained by low concentrations and analytical challenges. Recent advances in high-sensitivity methods are expanding their feasibility in natural systems. Nonetheless, these limitations continue to restrict their widespread use in environmental studies.
At the modelling scale, representing nitrite explicitly as a state variable, rather than collapsing nitrification and denitrification into net transformations, would improve mechanistic fidelity in biogeochemical models applied to nitrogen budgets and greenhouse gas inventories. The measurement and modelling strategies that follow from this reorientation are those organised around the node where nitrogen fate is actually decided.
As environmental systems experience increasing redox variability under climate change and land-use intensification, sensitivity at the nitrite stage is likely to amplify (Callbeck et al., 2026). Understanding nitrogen cycling at Earth-system scale requires resolving where and how nitrite flux is redistributed. Without explicit consideration of turnover at this branching node, interpretations of nitrogen budgets, greenhouse gas emissions, and isotopic signals remain incomplete.
No new data or code were generated for this study. The perspective is based exclusively on bibliographic sources, all of which are cited in the reference list.
MS and RM jointly conceived the perspective and wrote the manuscript.
The contact author has declared that neither of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We gratefully acknowledge the constructive feedback of the two anonymous reviewers and the editor, which significantly improved this manuscript.
This paper was edited by David Janssen and reviewed by two anonymous referees.
Babbin, A. R., Buchwald, C., Morel, F. M. M., Wankel, S. D., and Ward, B. B.: Nitrite oxidation exceeds reduction and fixed nitrogen loss in anoxic Pacific waters, Mar. Chem., 224, 103814, https://doi.org/10.1016/j.marchem.2020.103814, 2020.
Bristow, L. A., Dalsgaard, T., Tiano, L., Mills, D. B., Bertagnolli, A. D., Wright, J. J., Hallam, S. J., Ulloa, O., Canfield, D. E., and Revsbech, N. P.: Ammonium and nitrite oxidation at nanomolar oxygen concentrations in oxygen minimum zone waters, P. Natl Acad. Sci. USA, 113, 10601–10606, 2016.
Brunner, B. and Bernasconi, S. M.: A revised isotope fractionation model for dissimilatory sulfate reduction in sulfate reducing bacteria, Geochim. Cosmochim. Ac., 69, 4759–4771, 2005.
Buchwald, C. and Casciotti, K. L.: Oxygen isotopic fractionation and exchange during bacterial nitrite oxidation, Limnol. Oceanogr., 55, 1064–1074, 2010.
Buchwald, C. and Casciotti, K. L.: Isotopic ratios of nitrite as tracers of the sources and age of oceanic nitrite, Nat. Geosci., 6, 308–313, 2013.
Buchwald, C., Santoro, A. E., McIlvin, M. R., and Casciotti, K. L.: Oxygen isotopic composition of nitrate and nitrite produced by nitrifying cocultures and natural marine assemblages, Limnol. Oceanogr., 57, 1361–1375, 2012.
Burgin, A. J. and Hamilton, S. K.: Have we overemphasized the role of denitrification in aquatic ecosystems? A review of nitrate removal pathways, Front. Ecol. Environ., 5, 89–96, 2007.
Burgin, A. J., Yang, W. H., Hamilton, S. K., and Silver, W. L.: Beyond carbon and nitrogen: how the microbial energy economy couples elemental cycles in diverse ecosystems, Front. Ecol. Environ., 9, 44–52, 2011.
Butterbach-Bahl, K., Baggs, E. M., Dannenmann, M., Kiese, R., and Zechmeister-Boltenstern, S.: Nitrous oxide emissions from soils: how well do we understand the processes and their controls?, Philos. T. R. Soc. B, 368, 20130122, https://doi.org/10.1098/rstb.2013.0122, 2013.
Callbeck, C. M., Mazzoli, A. P., Paulus, T. J., Frey, C., Bürgmann, H., Schuber, C. J., and Lehmann, M. F.: Seasonality of lake microbial denitrification and its sensitivity to climate warming, Nat. Microbiol., https://doi.org/10.1038/s41564-026-02349-9, 2026.
Canfield, D. E., Glazer, A. N., and Falkowski, P. G.: The evolution and future of Earth's nitrogen cycle, Science, 330, 192–196, 2010.
Casciotti, K. L.: Inverse kinetic isotope fractionation during bacterial nitrite oxidation, Geochim. Cosmochim. Ac., 73, 2061–2076, 2009.
Casciotti, K. L.: Nitrogen and oxygen isotopic studies of the marine nitrogen cycle, Annu. Rev. Mar. Sci., 8, 379–407, 2016.
Casciotti, K. L., McIlvin, M. R., and Buchwald, C.: Oxygen isotopic exchange and fractionation during bacterial ammonia oxidation, Limnol. Oceanogr., 55, 753–762, 2010.
Casciotti, K. L., Sigman, D. M., Hastings, M. G., Böhlke, J. K., and Hilkert, A.: Measurement of the oxygen isotopic composition of nitrate in seawater and freshwater using the denitrifier method, Anal. Chem., 75, 4905–4912, 2003.
Daims, H., Lebedeva, E. V., Pjevac, P., Han, P., Herbold, C., Albertsen, M., Jehmlich, N., Palatinszky, M., Vierheilig, J., Bulaev, A., Kirkegaard, R. H., von Bergen, M., Rattei, T., Bendinger, B., Nielsen, P. H., and Wagner, M.: Complete nitrification by Nitrospira bacteria, Nature, 528, 504–509, 2015.
Dalsgaard, T., Thamdrup, B., Farías, L., and Revsbech, N. P.: Nitrogen cycling in a deep-water oxygen-deficient zone of the Arabian Sea, Limnol. Oceanogr., 57, 1331–1346, 2012.
Deb, S. and Lewicka-Szczebak, D.: Simplified bacterial denitrification method using Stenotrophomonas nitritireducens for nitrite dual isotope analysis in low-concentration environmental samples, Front. Environ. Sci. 13, 1536882, https://doi.org/10.3389/fenvs.2025.1536882, 2025.
Deb, S., Espenberg, M., Well, R., Bucha, M., Jakubiak, M., Mander, Ü., Jędrysek, M.-O., and Lewicka-Szczebak, D.: N transformations in nitrate-rich groundwaters: combined isotope and microbial approach, Biogeosciences, 22, 5535–5556, https://doi.org/10.5194/bg-22-5535-2025, 2025.
Deb, S., Lewicka-Szczebak, D., and Rohe, L.: Microbial nitrogen transformations tracked by natural abundance isotope studies and microbiological methods: a review, Sci. Total Environ., 926, 172073, https://doi.org/10.1016/j.scitotenv.2024.172073, 2024.
Denk, T.R.A., Mohn, J., Decock, C., Lewicka-Szczebak, D., Harris, E., Butterbach-Bahl, K., Kiese, R., and Wolf, R.: The nitrogen cycle: A review of isotope effects and isotope modeling approaches, Soil Biol. Biochem., 105, 121–137, 2017.
Firestone, M. K. and Davidson, E. A.: Microbiological basis of NO and N2O production and consumption in soil, in: Exchange of Trace Gases between Terrestrial Ecosystems and the Atmosphere, edited by: Andreae, M. O. and Schimel, D. S., Wiley, New York, 7–21, https://hero.epa.gov/reference/92805/ (last access: 6 July 2026), 1989.
Galloway, J. N., Dentener, F. J., Capone, D. G., Boyer, E. W., Howarth, R. W., Seitzinger, S. P., Asner, G. P., Cleveland, C. C., Green, P. A., Holland, E. A., Karl, D. M., Michaels, A. F., Porter, J. H., Townsend, A. R., and Vörösmarty, C. J.: Nitrogen cycles: past, present, and future, Biogeochemistry, 70, 153–226, 2004.
Grabb, K. C., Buchwald, C., Hansel, C. M., and Wankel, S. D.: A dual nitrite isotopic investigation of chemodenitrification by mineral-associated Fe(II) and its production of nitrous oxide, Geochim. Cosmochim. Ac., 196, 388–402, 2017.
Granger, J. and Wankel, S. D.: Isotopic overprinting of nitrification on denitrification as a ubiquitous and unifying feature of environmental nitrogen cycling, P. Natl Acad. Sci. USA, 113, E6391–E6400, 2016.
Gruber, N. and Galloway, J. N.: An Earth-system perspective of the global nitrogen cycle, Nature, 451, 293–296, 2008.
Heil, J., Vereecken, H., and Brüggemann, N.: A review of chemical reactions of nitrification intermediates and their role in nitrogen cycling and nitrogen trace gas formation in soil, Eur. J. Soil Sci., 67, 23–39, 2016.
Hu, J., Yao, S.J., Li, Y., Ren, H., Yang, K., and Liu, W.: Accurate isotopic analysis of nitrate and nitrite in freshwater: integration of ion exchange and bacterial denitrification methods, J. Anal. Atom. Spectrom., 41, 1774–1783, 2026.
Jiang, M., Koba, K., Ono, M., and Hayashi, K.: Improved Isotopic Analysis of Low-Concentration Freshwater Nitrite by Anion-Exchange Resin Enrichment and Azide Reduction, Anal. Chem. 98, 4, 2956–2967, 2026.
Jones, L. C., Peters, B., Pacheco, J. S. L., Casciotti, K. L., and Fendorf, S.: Stable isotopes and iron oxide mineral products as markers of chemodenitrification, Environ. Sci. Technol., 49, 3444–3452, 2015.
Kalvelage, T., Jensen, M. M., Contreras, S., Revsbech, N. P., Lam, P., Günter, M., LaRoche, J., Lavik, G., and Kuypers, M. M. M.: Nitrogen cycling driven by organic matter export in the South Pacific oxygen minimum zone, Nat. Geosci., 6, 228–234, 2013.
Karolewski, J. S., Sutherland, K. M., Hansel, C. M., and Wankel, S. D.: An isotopic study of abiotic nitrite oxidation by ligand-bound manganese (III), Geochim. Cosmochim. Ac., 293, 365–378, 2021.
Kartal, B., Kuenen, J. G., and van Loosdrecht, M. C. M.: Molecular mechanism of anaerobic ammonium oxidation, Nature, 479, 127–130, 2011.
Kraft, B., Strous, M., and Tegetmeyer, H. E.: Microbial nitrate respiration – genes, enzymes and environmental distribution, J. Biotechnol., 155, 104–117, 2011.
Lam, P. and Kuypers, M. M. M.: Microbial nitrogen cycling processes in oxygen minimum zones, Annu. Rev. Mar. Sci., 3, 317–345, 2011.
Lewicka-Szczebak, D., Lewicki, M. P., and Well, R.: N2O isotope approaches for source partitioning of N2O production and estimation of N2O reduction – validation with the 15N gas-flux method in laboratory and field studies, Biogeosciences, 17, 5513–5537, https://doi.org/10.5194/bg-17-5513-2020, 2020.
Lewicka-Szczebak, D., Jansen-Willems, A., Müller, C., Dyckmans, J., and Well, R.: Nitrite isotope characteristics and associated soil N transformations, Sci. Rep., 11, 5008, https://doi.org/10.1038/s41598-021-83786-w, 2021.
Luther, G. W., Karolewski, J. S., Sutherland, K. M., Hansel, C. M., and Wankel, S. D.: The abiotic nitrite oxidation by ligand-bound manganese (III): the chemical mechanism, Aquat. Geochem., 27, 207–220, 2021.
Margalef-Marti, R., Bourbonnais, A., Knöller, K., Mayer, B., Altabet, M., and Sebilo, M.: Using N and O isotope fractionation for evaluating denitrification in aquatic systems, TrAC Trends Anal. Chem., 194, 118527, https://doi.org/10.1016/j.trac.2025.118527, 2026.
McIlvin, M. R. and Altabet, M. A.: Chemical conversion of nitrate and nitrite to nitrous oxide for nitrogen and oxygen isotopic analysis in freshwater and seawater, Anal. Chem., 77, 5589–5595, 2005.
Robinson, T. C., Latta, D. E., Notini, L., Schilling, K. E., and Scherer, M. M.: Abiotic reduction of nitrite by Fe(II): a comparison of rates and N2O production, Environ. Sci. Process. Impacts, 23, 1531–1541, 2021.
Santoro, A. E. and Casciotti, K. L.: Enrichment and characterization of ammonia-oxidizing archaea from the open ocean, ISME J., 5, 1796–1808, 2011.
Sebilo, M., Billen, G., Grably, M., and Mariotti, A.: Isotopic composition of nitrate-nitrogen as a marker of riparian and benthic denitrification at the scale of the whole Seine River system, Biogeochemistry, 75, 35–51, 2006.
Sebilo, M., Aloisi, G., Mayer, B., Perrin, E., Vaury, V., Mothet, A. and Laverman, A.M.: Controls on the isotopic composition of nitrite (δ15N and δ18O) during denitrification in freshwater sediments, Sci. Rep., 9, https://doi.org/10.1038/s41598-019-54014-3, 2019.
Toyoda, S., Yoshida, N., and Koba, K.: Isotopocule analysis of biologically produced nitrous oxide in various environments, Mass Spectrom. Rev., 36, 135–160, 2017.
van Kessel, M. A. H. J., Speth, D. R., Albertsen, M., Nielsen, P. H., Op den Camp, H. J. M., Kartal, B., Jetten, M. S. M., and Lücker, S.: Complete nitrification by a single microorganism, Nature, 528, 555–559, 2015.
Vitousek, P. M., Aber, J. D., Howarth, R. W., Likens, G. E., Matson, P. A., Schindler, D. W., Schlesinger, W. H., and Tilman, D. G.: Human alteration of the global nitrogen cycle: sources and consequences, Ecol. Appl., 7, 737–750, 1997.
Ward, B. B., Devol, A. H., Rich, J. J., Chang, B. X., Bulow, S. E., Naik, H., Pratihary, A., and Jayakumar, A.: Denitrification as the dominant nitrogen loss process in the Arabian Sea, Nature, 461, 78–81, 2009.
Wrage, N., Velthof, G. L., van Beusichem, M. L., and Oenema, O.: Role of nitrifier denitrification in the production of nitrous oxide, Soil Biol. Biochem., 33, 1723–1732, 2001.
Zumft, W. G.: Cell biology and molecular basis of denitrification, Microbiol. Mol. Biol. Rev., 61, 533–616, 1997.
- Abstract
- Introduction
- Nitrite as the structural branching node
- Kinetic control of nitrite turnover and gaseous nitrogen speciation
- Isotopic constraints on gross nitrite turnover
- Sensitivity across environmental gradients
- Nitrite as the control point of nitrogen fate
- Code and data availability
- Author contributions
- Competing interests
- Disclaimer
- Acknowledgements
- Review statement
- References
- Abstract
- Introduction
- Nitrite as the structural branching node
- Kinetic control of nitrite turnover and gaseous nitrogen speciation
- Isotopic constraints on gross nitrite turnover
- Sensitivity across environmental gradients
- Nitrite as the control point of nitrogen fate
- Code and data availability
- Author contributions
- Competing interests
- Disclaimer
- Acknowledgements
- Review statement
- References