the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 20 May 2026
| 20 May 2026
Sensitivity of a mechanistic photosynthesis model to tropical Andean species and environments
Sebastián González-Caro
Mirindi Eric Dusenge
Zorayda Restrepo
Andrew J. F. Cox
Ian P. Hartley
Patrick Meir
Adriana Sánchez
Daniel Ruiz-Carrascal
Lina M. Mercado
Andean tropical montane forests are highly biodiverse ecosystems with a carbon storage capacity comparable to lowland forests. However, their response to climate change remains uncertain, as species photosynthesis depends on their thermal acclimation capacity. This study evaluates the variability of photosynthetic traits across montane and lowland tree species using a leaf level photosynthesis model and data from a transplant experiment across three elevations varying in their mean annual temperatures (14, 22, and 26 °C) in the tropical Andes. Six montane species and two lowland species were analyzed to assess photosynthetic responses to environmental conditions. We find that intraspecific variability in photosynthetic parameters, such as the apparent maximum carboxylation capacity and the apparent maximum electron transport rate, is key to accurately model photosynthesis in these ecosystems. Apparent maximum carboxylation capacity was identified as the primary determinant of diurnal variations in photosynthesis, especially under varying thermal environments. Additionally, stomatal conductance was highly variable and responded to vapor pressure deficit, suggesting that stomatal regulation is crucial for adaptation to environmental changes. Sensitivity analysis revealed that at higher altitudes (14 °C), photosynthetically active radiation and temperature were the main limiting factors for photosynthesis, while at lower altitudes (22 °C), vapor pressure deficit was the dominant factor. Finally, the study demonstrates that the common use, within global vegetation models, of average parameters from lowland species to simulate montane forest is inadequate as such parameterizations tend to underestimate montane forest photosynthesis by up to 65 %. It is also recommended that vegetation models incorporate both intra- and interspecific variability to improve predictions of the carbon cycle in tropical Andean forests and their response to climate change.
- Article
(3164 KB) - Full-text XML
-
Supplement
(976 KB) - BibTeX
- EndNote




Tropical montane forests are among the planet's most biodiverse ecosystems (Myers et al., 2000) and play a critical role in global carbon storage. Large-scale syntheses have shown that these ecosystems store substantial amounts of aboveground carbon, sometimes comparable to that of tropical lowland forests when differences in temperature and productivity are considered (Cuni-Sanchez et al., 2021; Duque et al., 2021). In the Andes, for example, Duque et al. (2021, Table 1) reported a monotonic decline in aboveground carbon stocks – from about 89 Mg C ha−1 at 500–1200 m to 59 Mg C ha−1 at 2800–3600 m a.s.l. – yet these values remain high relative to the cooler, less productive conditions under which they occur. Such evidence suggests that, although biomass decreases with elevation, tropical montane forests maintain unexpectedly large carbon stores, likely supported by physiological adjustments that sustain carbon assimilation under cold and cloudy environments (Fyllas et al., 2017; Girardin et al., 2010). While biomass per unit land area integrates processes such as recruitment, mortality, and allocation, these processes are ultimately constrained by carbon assimilation at the leaf level (Bahar et al., 2017; Malhi et al., 2017; van de Weg et al., 2014). Photosynthesis therefore defines the physiological potential for carbon gain, and its capacity to acclimate to temperature may explain how tropical montane forests maintain relatively high carbon stocks despite cooler climates. Experimental evidence further indicates that photosynthetic capacities of tropical montane and lowland trees are similar when both are grown under comparable mid-elevation conditions (Dusenge et al., 2021; Cox et al., 2023a; Dusenge et al., 2025). However, when evaluated in their native environments – cooler high-elevation or warmer low-elevation sites – photosynthetic capacities diverge. This pattern supports the idea that tropical montane species can acclimate, at least partially, to local thermal conditions, providing a mechanistic explanation for the stability of montane forest productivity under a warming climate.
Such thermal plasticity of photosynthetic capacity could enable montane species to sustain tree growth and aboveground biomass accumulation at rates in highland forests comparable to those in lowland forests. Nevertheless, it remains uncertain whether this physiological acclimation is sufficient to maintain growth and photosynthetic functioning of montane species under the warmest experimental conditions they experience. Observational studies in tropical Andean and Afromontane forests suggest that species composition has shifted in response to recent warming, likely driven by selective mortality of montane tree species to increased mortality at lower elevations (Duque et al., 2015; Fadrique et al., 2018). Specifically, lowland tree species have been observed migrating upslope, while montane species experience range contractions due to high mortality at the warmest margins of their geographical distribution – an effect known as thermophilisation (Duque et al., 2015; Fadrique et al., 2018; Cuni-Sanchez et al., 2024). This mortality is thought to result from thermal limitations to photosynthesis and associated carbon imbalance under extreme warming. At temperatures near or above the optimum for photosynthesis, enzymatic activity declines and photorespiration increases, reducing net carbon gain (Mercado et al., 2018). When this reduction in carbon assimilation persists, it can lead to growth suppression and eventually higher mortality for species living at the warmest margins of their distributions, where physiological acclimation may no longer compensate for thermal stress. To better assess species' ability to maintain photosynthetic function under rapidly changing conditions, characterised by accelerated warming, reduced cloud immersion, and altered moisture regimes in tropical montane regions (Fadrique et al., 2018; Duque et al., 2021), a combination of experimental and modeling is required, particularly in the context of increasing environmental extremes.
Most global vegetation, land surface, and Earth system models most vegetation models currently represent both montane and lowland tropical forests using a single plant functional type, which neglects the physiological diversity and thermal specialization of species across elevations (Fisher et al., 2018; Harper et al., 2018; Poulter et al., 2011). But it is now known that species from these forests differ in key photosynthetic traits and that these traits are plastic in response to warming (Cox et al., 2024; Dusenge et al., 2021; Kumarathunge et al., 2019). Furthermore, these models scale up leaf-level physiological processes, such as photosynthesis, respiration, stomatal conductance and transpiration, from leaf to canopy level which is then used to perform simulations at site, regional and global levels. Therefore, model parameterizations and evaluations at the leaf level are crucial for advancing model accuracy. Photosynthesis is one of the key processes at the heart of these models. However, photosynthetic traits employed in land surface and Earth system model predictions are commonly averaged across species (Fisher et al., 2018; Oliver et al., 2022; Poulter et al., 2011), and the (interspecific and intraspecific) variance is rarely considered in such large scale process-based models. This simplification can be problematic if systematic differences in key physiological parameters exist among species groups, for example the differential responses to warming observed for tropical montane versus lowland trees (Cox et al., 2023a; Mujawamariya et al., 2023). While it is neither feasible nor necessary to parameterize every species in these highly diverse forests, identifying consistent physiological contrasts among major functional groups can improve model realism without excessive complexity. Given the substantial contribution of tropical montane forests to global carbon cycling (Cuni-Sanchez et al., 2021; Duque et al., 2021), there is a need for leaf-level models parameterized and evaluated with montane tropical data to inform large-scale models and produce more reliable estimates of regional carbon budgets and their responses to future climate.
Tropical Andean forests are known to harbour high biodiversity (Myers et al., 2010) with communities composed of species that differ widely in their functional traits and environmental tolerances (Fyllas et al., 2017; González-Caro et al., 2023). Consequently, substantial interspecific variation in photosynthetic traits can be expected within any given location. Such variation implies that the use of single average parameter values in models (Knauer et al., 2023; Oliver et al., 2022) may obscure the actual diversity of physiological responses to environmental conditions. Andean species are exposed to a variety of thermal conditions because of the large range in environmental conditions across elevational bands, including within a single location through large diurnal variation (Martínez et al., 2011; van de Weg et al., 2014). This is also reflected in the large thermal ranges of some abundant species which can be found within a natural mean annual temperature span of up to 15 to 20° (Montaño-Centellas et al., 2024). Additionally, fluctuating cloud and wind regimes create constantly changing environmental conditions, making it impractical for plants to maintain photosynthesis at optimal levels at all times. All these conditions could promote both high intraspecific variation and acclimation of photosynthetic traits in Andean tree species (Bahar et al., 2017; Cox et al., 2023a, 2024). Empirical observations and photosynthesis models can be used to assess the effect of intraspecific variance of photosynthetic traits on photosynthesis of Andean forests.
Mechanistic models, such as the widely used C3 photosynthesis model developed by Farquhar, von Caemmerer, and Berry (hereafter referred to as the FvCB model), effectively describe photosynthesis by integrating the biochemical drivers of this process along with the response of stomatal conductance to leaf-to-air vapor pressure deficit (Farquhar et al., 1980). The FvCB model is useful to analyse the impact of environmental drivers on photosynthesis by varying parameters that are closely associated with enzymatic activity, primarily related to Rubisco during the CO2 assimilation process. Previous modelling efforts have applied this framework to quantify the limitations of photosynthesis imposed by temperature, vapor pressure deficit (VPD), and light availability in tropical forests (Mercado et al., 2018; Smith et al., 2019). However, these studies have been largely based on lowland species, and the extent to which montane trees differ in their photosynthetic sensitivity to these environmental drivers remains poorly understood. Addressing this gap requires parameterisations derived from direct field measurements across contrasting elevations. Given the high environmental heterogeneity of tropical montane forests and the potentially plastic response capabilities of their native species (Cox et al., 2023a), the parameterisation of the FvCB model must therefore be approached cautiously to accurately capture both species response variability and environmental heterogeneity (Blonder et al., 2020).
In this study, we combined the FvCB model with field-based data from a transplant experiment in the tropical Andes to address two main questions: (i) what is the effect of intraspecific variation in plant physiological traits on photosynthetic performance and on the differences between montane and lowland species? We hypothesised that intraspecific variation in physiological traits contributes significantly to differences in leaf-level photosynthetic capacity among tropical montane tree species. Specifically, we predicted that species with higher photosynthetic capacity (e.g., higher and Jmax) would maintain greater carbon assimilation rates under warmer conditions. (ii) How do photosynthesis and related traits respond to environmental conditions at different sites, and how do these responses influence forest-level assimilation depending on the plant functional type (montane vs. lowland species)? We further hypothesised that environmental drivers such as air temperature, vapour pressure deficit (VPD), and light availability (PAR) exert distinct and interacting effects on photosynthesis across thermal regimes. Accordingly, we predicted that temperature would be the dominant limiting factor at high-elevation (cool) sites, whereas VPD would impose stronger constraints at low-elevation (warm) sites. Finally, we expected that including both plant functional types in simulations would increase overall assimilation rates and reduce their temporal variability within a site.
We used the FvCB processes-based C3 photosynthesis model along with leaf gas-exchange data from trees planted in the field to calibrate and evaluate photosynthesis models using different parameterisationsfor montane and lowland tree species. Subsequently, we investigate the sensitivity of simulated photosynthesis to key model parameters and various environmental conditions (Fig. 1).
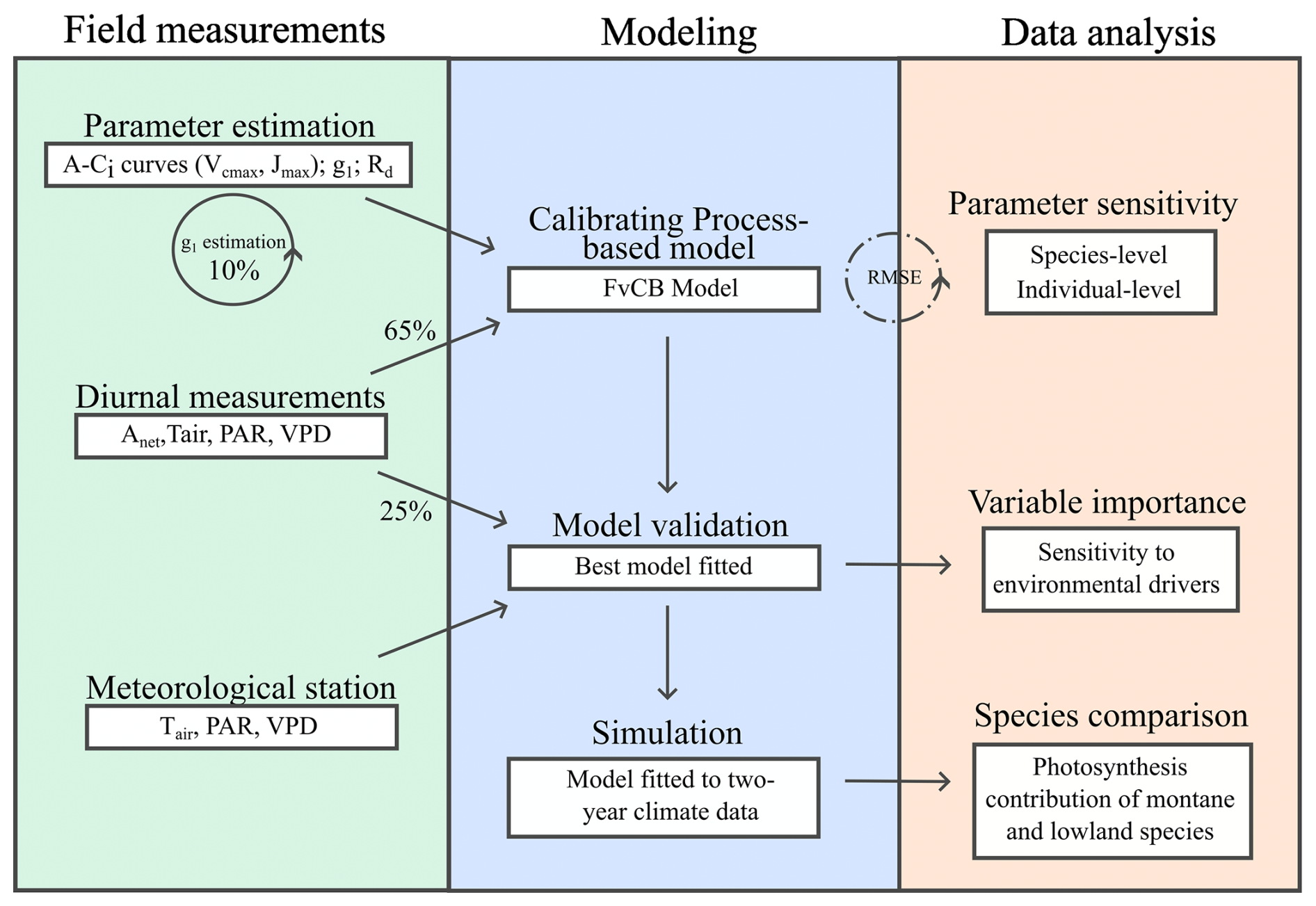
Figure 1Summary of methodology applied in this study comprising field measurements, modelling (calibration and evaluation) and data analysis related to sensitivity analyses.
2.1 Common garden transplant
The experiment is based on the transplant of eight dominant tropical Andean species across two sites at different elevations (1500 and 2500 m), where mean annual temperatures (14 and 22 °C) covaries with elevation but was not manipulated independently. The tree species are divided into two groups (Table 1): six montane tree species that naturally inhabit montane forests above 2000 m (where mean air temperatures range from 10–22 °C in their native distributions) were transplanted to the lower-elevation site with warmer temperature (22 °C). these conditions represent at the warm extreme of their thermal ranges and simulate the scenarios where range retractions of montane species have been reported in Andean forests (Duque et al., 2015; Fadrique et al., 2018). Conversely two lowland tree species that inhabit foothill forests below 1000 m (air temperatures ranging from 22–35 °C) were transplanted to high elevations/cooler temperatures (14 °C) to simulate upslope migration from the lowlands (Duque et al., 2015; Fadrique et al., 2018). Twenty-four saplings per species per site (total of trees planted per site n=360) were planted between December 2018 and January 2019 under common soils extracted from a nearby location to the 14 °C experimental site (i.e., the soils transported to each experimental site, 400 kg of soil used to plant each tree in soil pits of 0.32 m3) and were continuously watered to remove drought confounding effects.
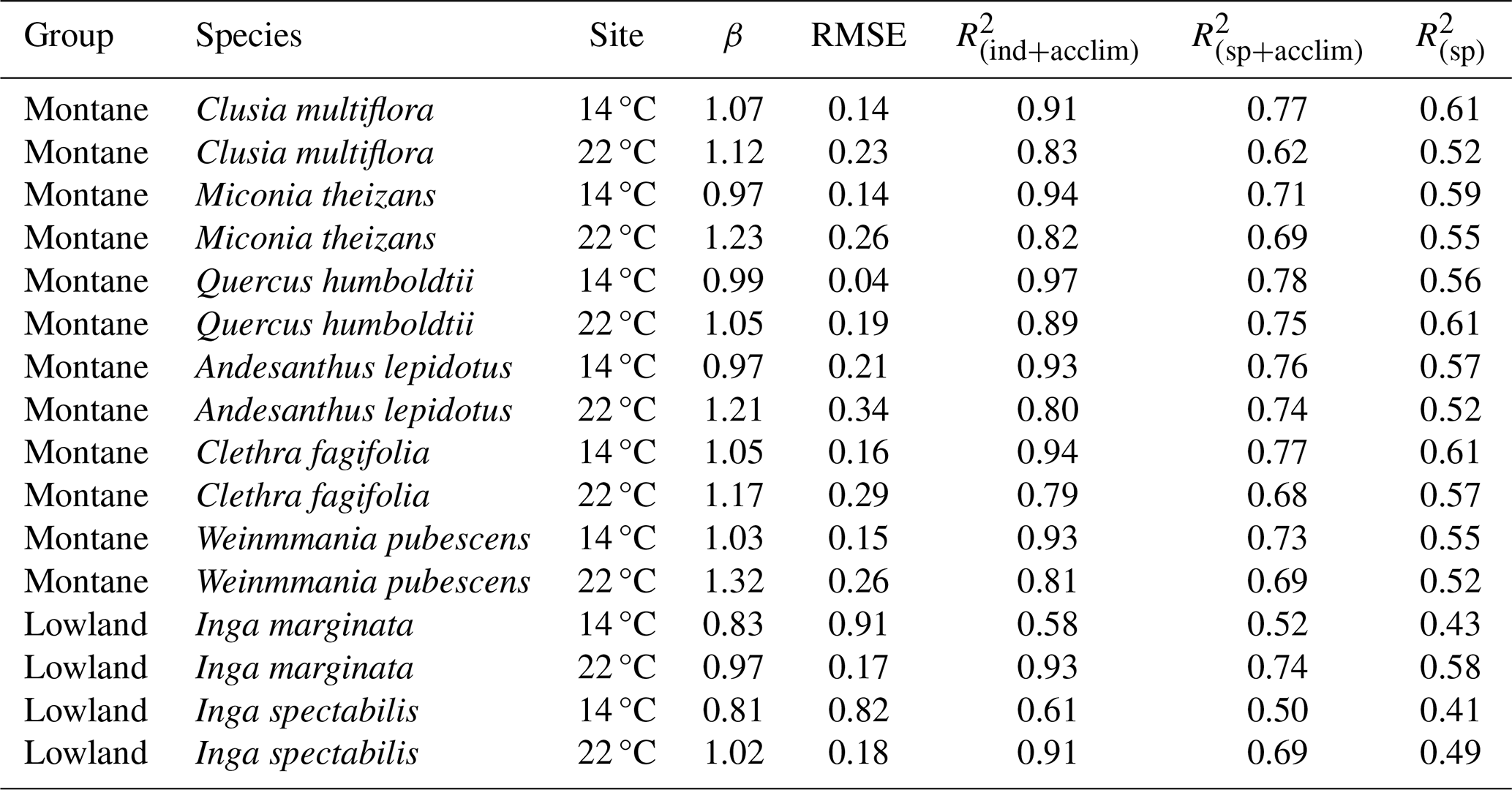
Table 1Photosynthesis model performance for all study species applied at two experimental sites (14 and 22 °C) using three indicators: regression slope between observed and predicted values (β), residual mean squared error (RMSE) of final model, and coefficient of determination (R2) at individual-level parametrization (iterative modelling approach) plus acclimation (ind + acclim), species level parametrization plus acclimation (sp + acclim) and species level parametrization without acclimation (sp). Species were separated in two groups (montane and lowland) accordingly to their thermal affiliation and elevational distribution.
2.2 Photosynthesis model
The FvCB model is a key tool for interpreting leaf gas exchange observations and for modelling photosynthesis. Its central function is to simulate photosynthesis reactions to changes in [CO2] concentrations inside the leaf, and it can adjust for enzymatic reaction temperature-dependence with appropriate parameter settings (Medlyn et al., 2002). The model represents net [CO2] assimilation rate (An expressed in mmol m−2 s−1) as the difference between the minimum of two limiting photosynthetic rates: the Rubisco-limited rate (Ac expressed in mmol m−2 s−1), which depends on the apparent maximum rate of Rubisco carboxylation (), and light limited rate (Aj, expressed in mmol m−2 s−1) which is related to the apparent maximum electron transport rate (Jmax), minus the leaf respiration in the dark (Rd expressed in mmol m−2 s−1):
Stomatal conductance is incorporated using the Medlyn et al. (2011) model as applied in Lin et al. (2015):
The model describes the diffusion of CO2 through mesophyll layers and the stomata response to leaf to air vapor pressure difference (D expressed in kPa). Ca (in ppm) is CO2 concentration at the leaf surface and g1 (kPa0.5) is a proxy for the plant water use strategy with small g1 values indicating a conservative water use and large g1 indicating the opposite (Lin et al., 2015).
2.3 Field leaf-level data and model application
We used gas exchange measurements from juvenile tropical Andean trees planted as part of a common garden transplant experiment in the Colombian Andes to derive parameters for the FvCB model. The experiment is based on the transplant of eight dominant tropical Andean species across two sites at different elevations (1500 and 2500 m), where mean annual temperatures (14 and 22 °C) covaries with elevation but was not manipulated independently. The species are divided into two groups: six montane tree species that naturally inhabit montane forests above 2000 m (where mean air temperatures range from 10–22 °C in their native distributions) were transplanted to the lower-elevation site with warmer temperature (22 °C). these conditions represent at the warm extreme of their thermal ranges and simulate the scenarios where range retractions of montane species have been reported in Andean forests (Duque et al., 2015; Fadrique et al., 2018). Conversely two lowland tree species that inhabit foothill forests below 1000 m (air temperatures ranging from 22–35 °C) were transplanted to high elevations/cooler temperatures (14 °C) to simulate upslope migration from the lowlands (Duque et al., 2015; Fadrique et al., 2018). Twenty-four saplings per species per site (total of trees planted per site n=360) were planted between December 2018 and January 2019 under common soils extracted from a nearby location to the 14 °C experimental site (i.e., the soils transported to each experimental site, 400 kg of soil used to plant each tree in soil pits of 0.32 m3) and were continuously watered to remove drought confounding effects.
We used leaf-level photosynthetic CO2 responses (A–Ci) collected using a Li-6800 portable photosynthesis system during two field campaigns that took place during June and July 2019 (Cox et al., 2023a) and during January and February 2022. The A–Ci data were measured under light-saturated conditions set at 1800 PPFD micromol photons per metre per second, 60 % relative humidity and by setting the block temperature of the Li-6800 to 29 °C. Parameterisation of Rd was done using measurements of foliar respiration in the dark collected on the same leaves that A–Ci were measured, maintaining the same relative humidity and temperature as A–Ci data (Cox et al., 2023a). A–Ci and Rd data were gathered from four individuals per species per site and were fed into the Photosyn function of the plantecophys R package (Duursma, 2015) to derive the photosynthetic parameters (apparent and Jmax) during each field campaign. Temperature response curves of apparent and Jmax were fitted with a modified Arrhenius equation (Medlyn et al. 2002) to estimate activation energy (Ea) and optimum temperature (Topt), while fixing deactivation energy (Hd) at 200 kJ mol−1 (Dusenge et al., 2025). These kinetics were derived from A–Ci curves measured at five to six temperatures per site, ensuring that parameters reflected species-specific acclimation responses rather than default assumptions. We evaluated impact of accounting for acclimation of the temperature response of apparent and Jmax on model performance using the thermal acclimation equations from (Kumarathunge et al., 2019). Non-acclimated temperature responses parameters for apparent and Jmax were taken from Afromontane forests as described in Cox et al. (2023a).
We recognize that our parameter estimations are based on Ci rather than Cc, due to limitations in estimating mesophyll conductance. As a result, our estimates should be interpreted as apparent and Jmax. Measuring mesophyll conductance is particularly challenging under tropical field conditions and introduces uncertainty due to interspecific variation. Moreover, no standard model currently exists to represent the temperature sensitivity of mesophyll conductance. Therefore, as in many studies (Dusenge et al., 2025), we used Ci-based parameterization for and Jmax, as well as for their acclimation models, which are useful for assessing temperature effects on photosynthesis in poorly studied tree species.
Diurnal cycles of net CO2 assimilation (An) under ambient conditions were also collected using a Li-6800 portable photosynthesis system by taking spot measurements at different times of the day during two field campaigns. Measurements were performed on clear-sky days representative of typical climatic conditions at each site and within the same seasonal window to ensure comparability between locations. We measured three individuals per species, taking hourly measurements from 09:00 to 16:00 LT over three consecutive days. Before each measurement, the Li-6800 leaf chamber was opened to record ambient air temperature, relative humidity, and incoming radiation incident on each leaf. These environmental conditions were then imposed in the Li-6800 chamber, and leaves were enclosed under those same conditions. Data were recorded once gas exchange stabilized, which usually occurred within a few seconds. Diurnal cycles of An were measured on four individuals per species per site. A subset of the data (10 %) was used to estimate the stomatal slope parameter (g1) for each species and field campaign based on the Medlyn et al. (2011) model, as implemented in the fitBB function (model option = “BBOpti”) of the plantecophys R package (Lin et al., 2015; Duursma, 2015). The remaining data were divided for model calibration (65 %) and evaluation (25 %) (Fig. 1).
Additionally, the following meteorological data from a Campbell weather station located at each experimental site were used in this study: air temperature (Tair, °C), relative humidity (that allow us estimate vapor pressure deficit (VPD, kPa)) and photosynthetic active radiation (PAR, µmol m−1 s−1) recorded from January 2020 to June 2022 which included periods of physiological data collection.
2.4 Model parameterization and evaluation
A two-step approach was used to parameterize the model. First, species level average parameters (apparent , apparent Jmax, g1, Rd) were used to run the model for each species at each site. To account for the observed intraspecific variability in our data sets (Cox et al., 2023a) but also reported elsewhere in the Andes (Bahar et al., 2017), the second approach uses the observed range of each parameter per species (i.e., the minimum, and maximum value in our dataset per each species) to select parameter combinations that improve model fitting. Here an iterative parameter selection procedure based on Monte Carlo Markov Chain (MCMC) methods was used: first, apparent was randomly selected within the parameter's range for the target species, then the Photosyn function was run to evaluate whether model performance improved relative to the previous step. If model performance did not improve, the algorithm repeated this procedure ten times. The regression slope, the coefficient of determination (R2), and the root mean squared error (RMSE) were used indicators of model performance. The expected slope of the linear model is one, the expected R2 is also one, and zero for expected RMSE. If model performance improved, a new apparent value close to the previous value got selected, the Photosyn function was rerun, and model performance was reassessed. This procedure was repeated 10 000 times to find the apparent parameter that best fitted the observations. After completing the search for apparent , the same procedure was applied to apparent Jmax, g1, and Rd. Parameter values from each iteration were used to calculate the probability distribution function of each parameter to represent the intraspecific variability that allows model fitting.
All simulations were performed with prescribed environmental drivers (PAR, Tleaf, VPD) as recorded by the Li-6800 during the An diurnal cycles and evaluated against the observed An. Model performance was assessed using a linear regression between the observed An from diurnal cycles and simulated using the Photosyn function by fixing the intercept of the linear model at zero. To evaluate model biases under different environmental conditions residuals of observed and simulated photosynthesis were plotted against Ci, PAR, Tleaf, VPD.
2.5 Sensitivity analysis
We evaluate the impact of intraspecific variability in photosynthetic traits on leaf level photosynthesis of tropical montane tree species comparing simulations from the two approaches used for model parameterization. The iterative approach allows to estimate a robust intraspecific distribution of physiological parameters for further interspecific comparisons.
To assess the relative contribution of physiological traits to diurnal variation of leaf level photosynthesis of tropical montane tree species at two thermal regimes (mean diurnal Tair, VPD and PAR during field campaigns for 14 and 22 °C sites), we used a Sobol index of variable importance which is particularly useful in complex models involving numerous inputs and nonlinear interactions. Sobol index is used to quantify the contribution and sensitivity of each input parameter (i.e. apparent , apparent Jmax, g1, Rd) or the impact of their change to the output variance of a modelled variable (i.e. An). The computation of Sobol index involves the creation of multiple model evaluations using systematically varied input values. Typically, a Monte Carlo method or other random sampling techniques are used to generate these evaluations. The indices are calculated based on the variance of model outputs due to the variance of each input. The equation for the first-order Sobol index is:
where, Y is the model output, Xi is the input parameter, and E[Y|Xi] is the expected output under Xi parameter.
Here Sobol indices are used to assess the effect of varying apparent , apparent Jmax, g1 and Rd, within their measured distributions in our data set, on simulated An under different environmental conditions throughout the diurnal cycle. For this sensitivity analysis, the photosynthesis model was applied using average 30 min weather station data recorded during the period that the measurements of diurnal cycles of Anet took place. Subsequently, a matrix containing 10 000 different values for each parameter, consistent with the distributions obtained from our iterative search approach (as previously described in Model paremeterisation and evaluation section) was constructed. The sobol_indices function from the sensobol R package (Puy et al., 2022) was used to calculate the Sobol indices.
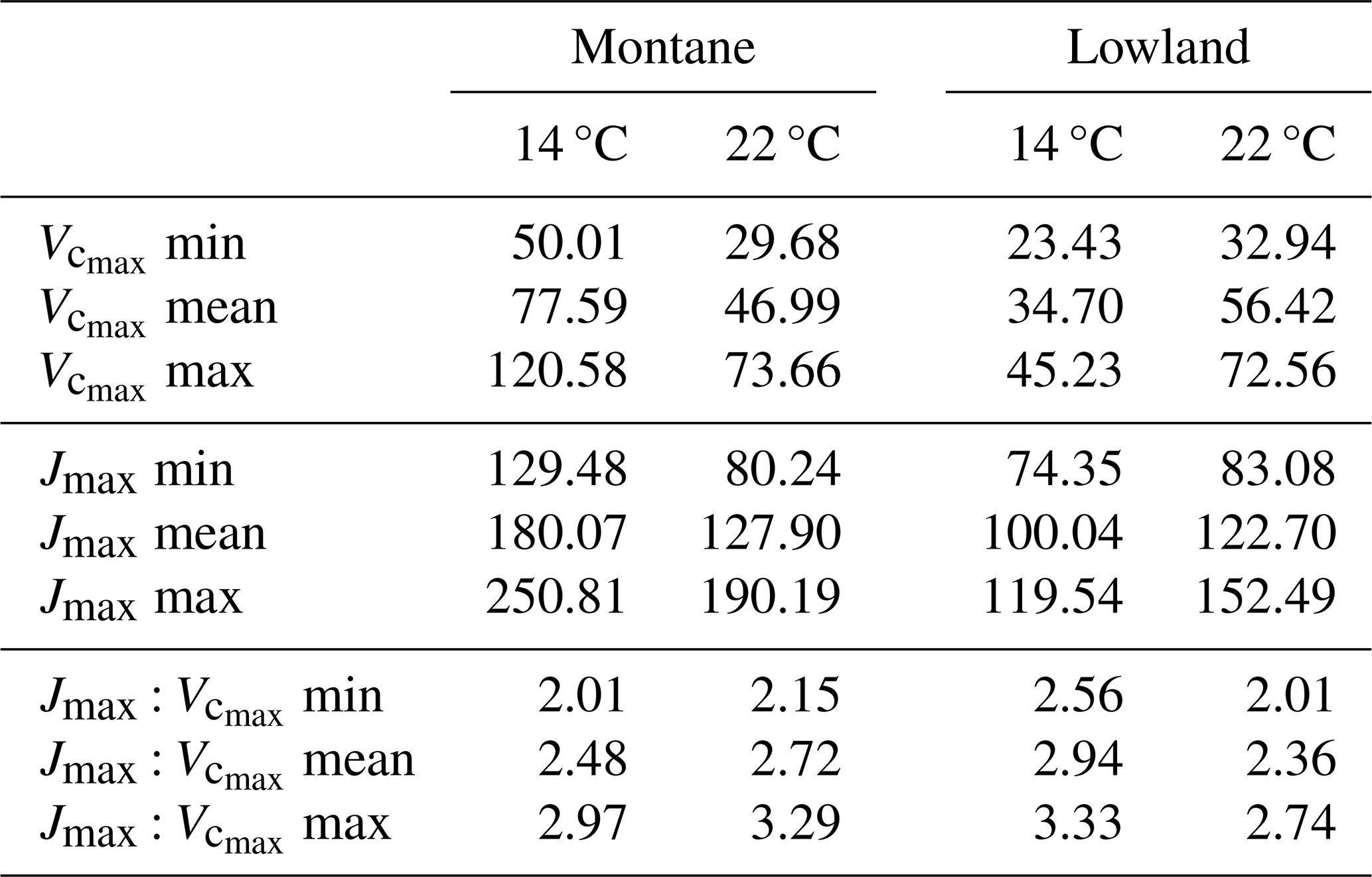
To assess the relative contribution of key environmental drivers of photosynthesis (Tair, VPD and PAR) in tropical montane tree species we also make use of the Sobol index. The computation of the Sobol index requires fixing some values while iterating over the target variable or parameter, in this case, the environmental variables. Therefore, we held the physiological parameters at known values representing the observed variation of these parameters in the studied species as follows. We used observed minimum, average, and maximum values of g1 (2.5, 5 and 7.5 kPa0.5) and the Jmax : ratio (1.75, 2.5 and 3) that represent both, montane and lowland species, and the full range of observed variations in (25 to 75 µmol m−2 s−1). Subsequently, we used Tair, VPD and PAR from the weather station at each site to define the range of each variable and constructed a matrix with 10 000 values per variable and then ran the Sobol index analysis.
2.6 Model application
To evaluate whether the common assumption in large-scale models – using mean photosynthetic parameters derived from lowland species to simulate montane forest photosynthesis – is a valid approach, we compared simulations including both montane and lowland tree species with those including only lowland species under identical environmental conditions. Photosynthesis was simulated using best-fit parameter values for montane and lowland species at each site, together with one year of hourly weather station data from the 14 and 22 °C sites. The relative abundances of montane and lowland species used in the simulations were based on field observations reported in previous studies of tropical Andean forests (Duque et al., 2015; González-Caro et al., 2020). Montane species represent approximately 75 % of the total species above 2200 m a.s.l. (14 °C site) and about 30 % between 1200–2000 m a.s.l. (22 °C site). Accordingly, we simulated forests with these respective proportions of montane and lowland species to reflect realistic community composition. The simulations aimed to assess how the magnitude and temporal stability of average photosynthesis change depending on whether one or both plant functional groups are considered. We simulated leaf-level photosynthesis for 1 year at each site under both scenarios (only lowland vs. lowland + montane species). Monthly totals were obtained by summing daily values, and scenarios were compared using paired t tests. Finally, we calculated the percentage change in photosynthesis between scenarios to quantify the influence of including montane species in model parameterisation.
3.1 Overall FvCB model performance
We modelled photosynthesis with the FvCB model for eight tree species, comprising six montane species and two lowland species from the tropical Andes. Inclusion of thermal acclimation of the temperature response of photosynthetic capacity improved model performance by ∼20 % (Table 1). Using the best approaches to select model parameters based in our analyses (iterative searching for physiological parameters) and thermal acclimation of the temperature response of photosynthetic capacity, model fitting showed high model performance (averaged R2=0.90; Fig. 2a), suggesting that the model with best fit parameters accurately simulate photosynthesis for these species when accounting for intraspecific variation (Fig. S1 in the Supplement, Table 1). Overall, simulated An represents the observed diurnal variation with high accuracy (Table 1), although there model tends to underpredict the observations at very high PAR or Tair at the 14 °C site and over predict at high VPD at the 22 °C site (Fig. S2). Model performance was best for all species at sites close to their native environments: specifically, species level model application for montane species was closest to observations at the 14 °C site and similarly for lowland species at the 22 °C (Table 1). The model application for montane species also showed very good performance under warming (i.e., 22 °C site), with the main model observational biases happening around noon (12:00–14:00 LT, Fig. S2) with a slight trend to overestimate the observations (β>1 in Table 1). In contrast, model application for lowland species under the cold environment (i.e., 14 °C site,) showed intermediate values of model performance (Inga marginata, R2=0.58 and Inga spectabilis, R2=0.61) and also exhibited a significant reduction in simulated An respect to simulations at the 22 °C site (Table 1; β=0.81 and β=0.83, respectively).
3.2 Intraspecific variability effect on model performance
Model performance improved across all species and sites when using the iterative parameter searching approach (R2=0.90, RMSE=0.20, β=1.09) compared to simulations that relied on average parameter values (R2=0.78, RMSE=0.52, β=0.91). This suggest that intraspecific variation of physiological traits plays a crucial role in enhancing model accuracy and reducing prediction errors (Tables 1 and 2). Specifically, apparent values obtained from the iterative searching method were, on averaged, 1.4 times higher than species-level mean values (Fig. 2b), emphasizing the importance of capturing trait variability rather than relying solely on species-level means. Similarly, the stomatal conductance parameter g1 also showed a variation in the selected value via iterative searching compared to the average observed value and the former tended to be highest, but their shift was no systematic (Fig. 2d). In contrast, the effect of apparent Jmax (Fig. 2c) and Rd in model calibration was negligible (plots for Rd not shown), indicating that not all physiological parameters contribute equally to model performance improvements.
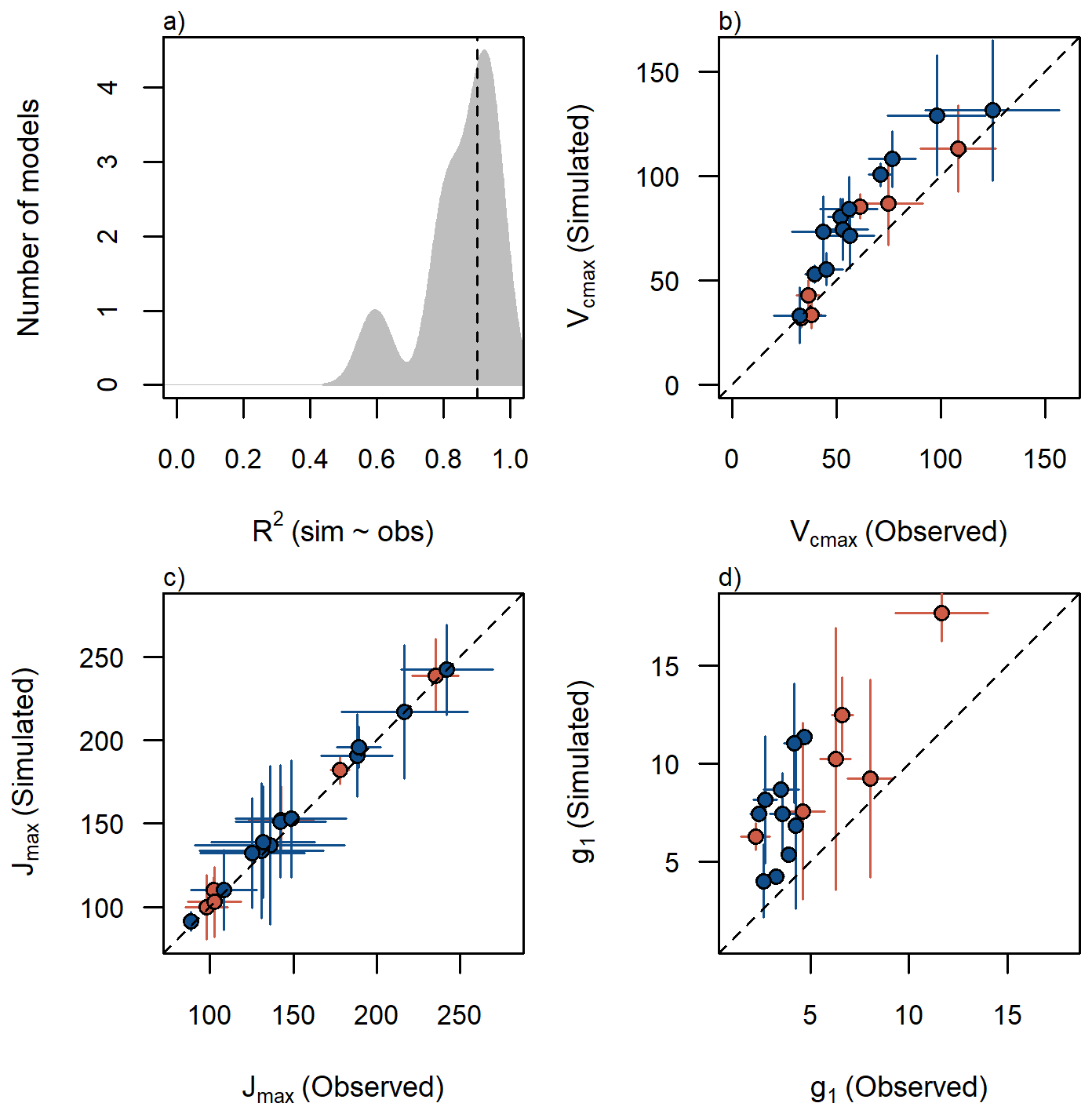
Figure 2Model performance using the iterative parameter searching approach. (a) R2 distribution of final best model parameterization per species per site for all species and sites. The dashed line represents the median. Relationship between observed average parameter value and predicted value by the iterative searching approach for (b) (µmol m−2 s−1), (c) Jmax (µmol m−2 s−1) and (d) g1 (kPa0.5) per species per site. Blue circles represent montane species, and red circles are lowland species. Error bars are added to each point representing standard error of observed values in y axis and range of simulated values in x axis. Dashed lines indicate the 1:1 relationship.
Table 2Photosynthetic parameters ( and Jmax in µmol m−2 s−1, and their ratio Jmax : ) expressed as maximum (max), minimum (min), and mean (mean) values for montane and lowland species at each site (14 C and 22 °C) used for sensitivity analyses. The maximum and minimum values represent the absolute range of fitted parameters across all individual observations for each species group.
3.3 Contribution of physiological traits to diurnal variation of simulated photosynthesis
Apparent was found to have the largest relative contribution to diurnal variation in simulated leaf level photosynthesis in all cases, accounting for ∼56 % (averaged throughout the day for all groups and sites, Fig. 3). The relative contribution of apparent is lower at the native environment for montane (∼52 %) and lowland (∼41 %) species than at 22 °C (∼61 %) or 14 ° (∼65 %) environments, respectively (Fig. 3). Specifically, at 14 °C, there were larger contributions from apparent Jmax to Anet in montane (17.2 %) than in lowland (9.5 %) species. And at 22 °C, lowland species had larger contribution from apparent Jmax (16.1 %) than montane species (5.7 %). Also, at 22 °C, g1 had the second largest contribution to An for both groups of species (Fig. 3c and d) (13.6 % montane and 16.8 % for lowland) with importance increasing in the afternoon in response to high leaf to air vapor pressure deficit. Respective g1 contributions to Anet at 14 °C were 6.5 % for montane and 6.2 % for lowland species (Fig. 3a and b). The contribution of Rd and interactions among parameters were low (∼4 %) and similar across species groups and sites throughout the day (Fig. 3).
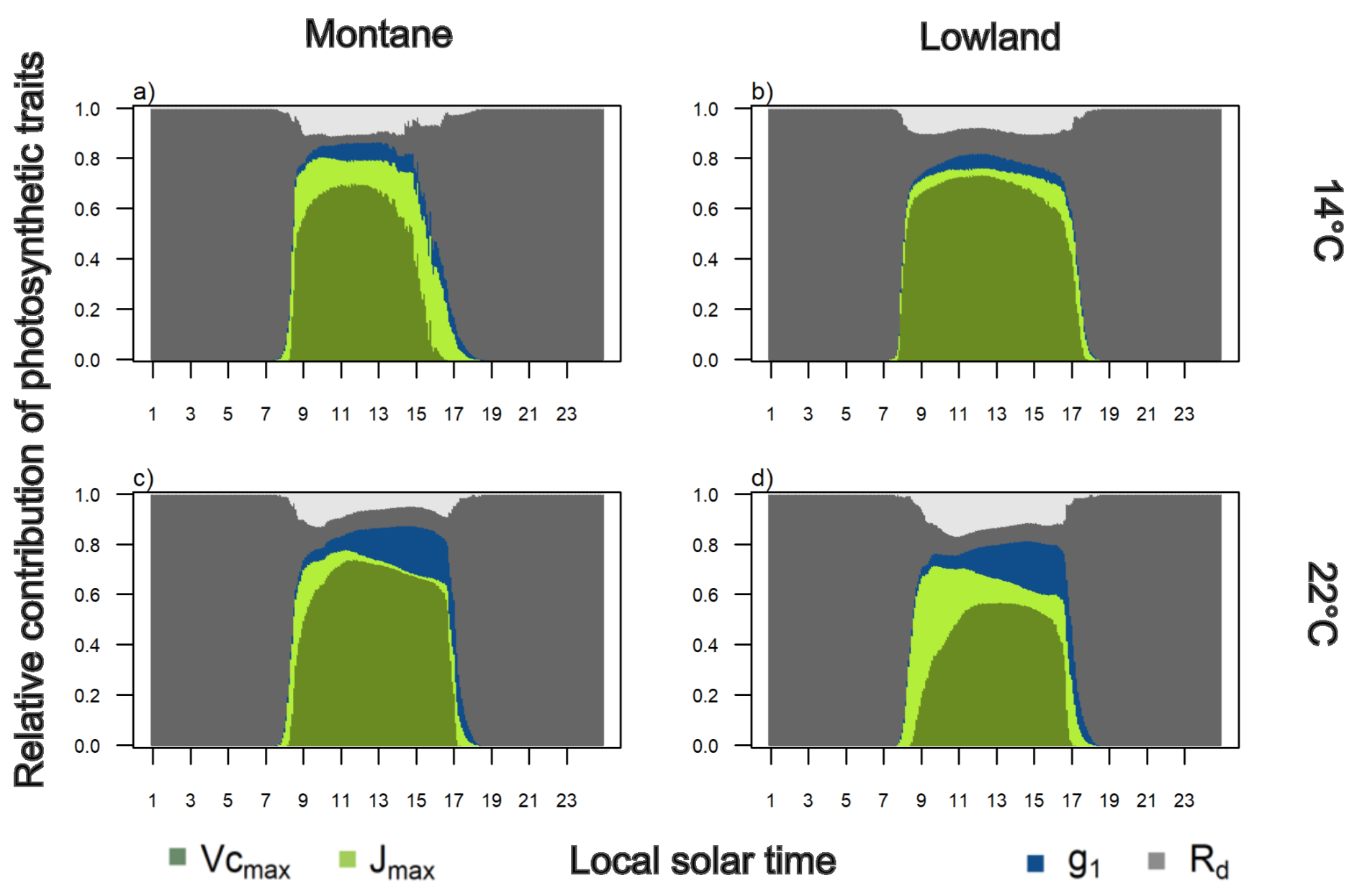
Figure 3Diel variation of the simulated relative contribution of physiological parameters to simulated leaf-level net carbon uptake for montane (a, c) and lowland species (b, d), at two growth temperatures (14 and 22 °C). Light grey represents interaction (Int) among study parameters.
3.4 Impact of environmental conditions on leaf photosynthesis of tropical montane species
We found differences in the influence of environmental variables (Tair, VPD and PAR) on simulated An between both sites, 14 °C (Fig. 4) and 22 °C (Fig. 5). Overall, at 14 °C PAR and Tair are the main drivers of An with contributions of 48 % and 39 % respectively (values averaged across comparisons; Fig. 4). At 22°, VPD and Tair are the main drivers of An with a contribution of 37 % and 28 % (averaged across comparisons; Fig. 5). In sum, the relative contributions of PAR and Tair to An are larger at 14 °C than at 22 °C and vice versa for VPD.
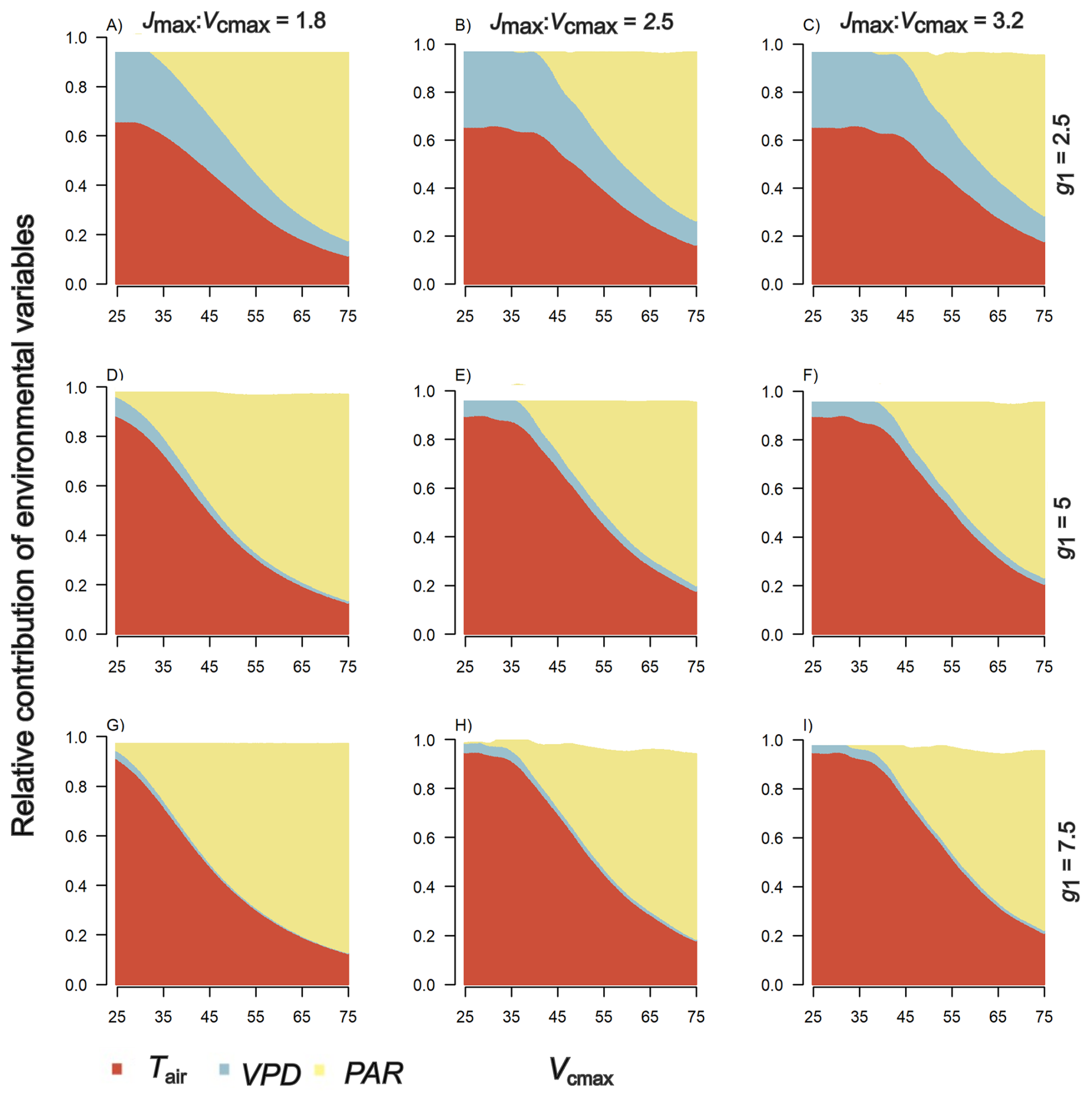
Figure 4Simulated relative contribution of environmental conditions – Tair (14–25 °C), VPD (0.6–2.4 kPa) and PAR (200–1600 µmol m−2 s−1) to leaf level net photosynthetic uptake at the 14 °C site. The x axis is the (µmol m−2 s−1) range of most common values of this parameter in field data. Panel columns represent different values of the ratio between Jmax (µmol m−2 s−1) and (µmol m−2 s−1) observed in field data (Table 2). Panel rows represent different g1 (kPa0.5) values observed in montane and lowland species.
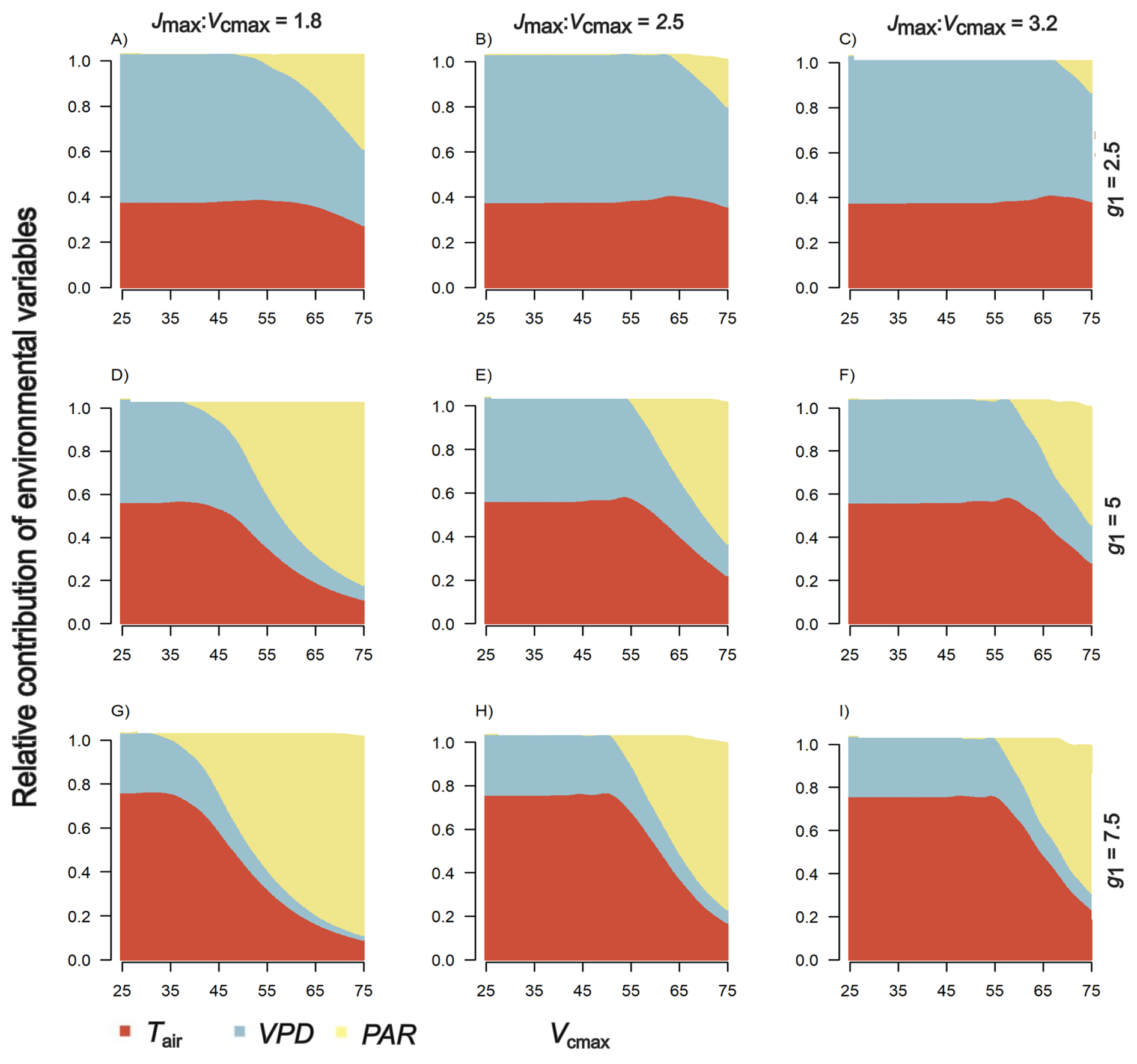
Figure 5Simulated relative contribution of environmental conditions – Tair (19–35 °C), VPD (1–3.8 kPa) and PAR (400–1600 µmol m−2 s−1) – to the 22 °C site to leaf level net photosynthetic uptake. The x-axis is the (µmol m−2 s−1) range of most common values of this parameter in field data. Panel columns represent different values of the ratio between Jmax (µmol m−2 s−1) and (µmol m−2 s−1) observed in field data (Table 2). Panel rows represent different g1 (kPa0.5) values observed in montane and lowland species.
Specifically, at the 14 °C site, for the range of studied values of Jmax: for both montane and lowland species, photosynthesis is primarily controlled by temperature and PAR (Fig. 4), and their relative contributions are mediated by g1 (i.e., stomatal conductance). When g1 is close to 2.5, meaning a conservative plant water use strategy, the relative importance of VPD increases with decreasing apparent (Fig. 4a–c), whereas under g1 values above 2.5 (more wasteful water use strategy), air temperature increases its relative importance (Fig. 4d–f). Under high g1 values, PAR has a large contribution to photosynthesis at low Jmax: and the relative importance of Tair increases with increasing Jmax: (Fig. 4g–i). On the other hand, at the 22 °C site, photosynthesis is primarily controlled by VPD under low g1 values (2.5), independently of Jmax: (Fig. 5a–c). When, g1 increases, the influence of VPD decreases while the contribution of Tair increases (Fig. 5d–f). Additionally, under low Jmax: values, the relative contribution of PAR increases (Fig. 5d and g).
3.5 Relative contribution of montane and lowland species to average photosynthesis
We found large differences between simulations that included both montane and lowland species and those based solely on lowland species. At the 14 °C site, simulations incorporating both groups produced larger annual photosynthesis than including solely lowland species (; P<0.001; Fig. 6a). On average, 65 % higher photosynthesis during at least six months of the year compared to simulations using only lowland species parameters. This period coincided with lower solar radiation levels. Conversely, during months with higher solar radiation and reduced cloud cover, simulations including both groups yielded ∼25 % lower photosynthesis than those using only lowland species (Fig. 6a). In contrast, at the 22 °C site, simulations using only lowland species consistently resulted in 29 % higher photosynthesis, on average, across the entire year compared to simulations including both species groups (Fig. 6b).
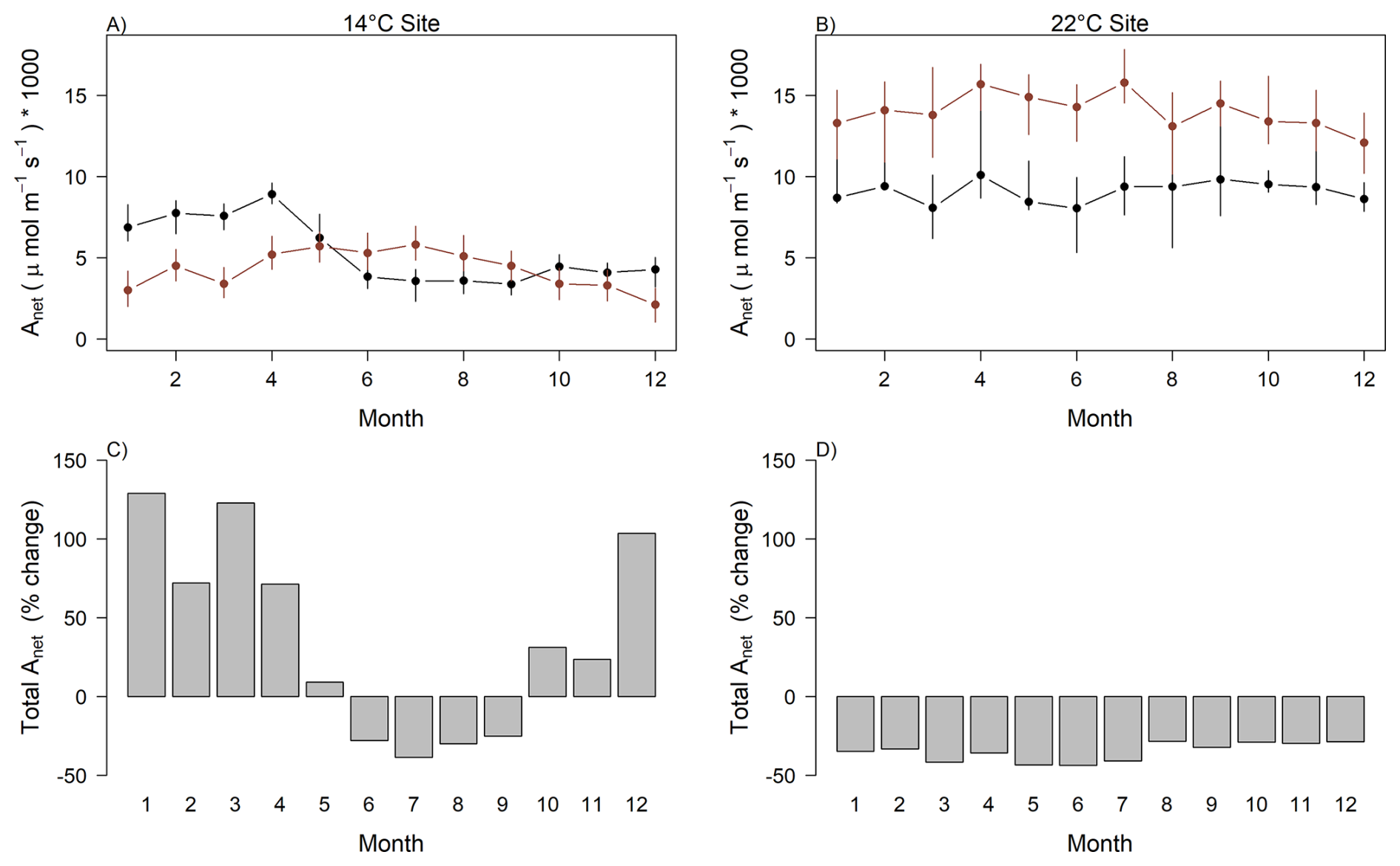
Figure 6Simulated leaf level photosynthesis – Anet (µmol m−2 s−1) – including montane and lowland species weighing their contribution accordingly to their abundance in a forest of 1000 individuals (black) and only lowland species assuming all individuals correspond to this group (red) under environmental conditions at the (a) 14 °C site and at the (b) 22 °C site. Percentage change in the photosynthesis including both species groups (montane and lowland) relative to only include lowland species for each site, respectively (c, d).
4.1 Intraspecific variability of photosynthetic traits
Our results display large variability in photosynthetic traits of montane tree species. This ranges from species with higher photosynthesis capacity like Miconia theizans (apparent mmol m−2 s−1and apparent Jmax=210 mmol m−2 s−1) to Quercus humboldtii (apparent mmol m−2 s−1 and apparent Jmax=115 mmol m−2 s−1; Fig. S1). The high interspecific variance in photosynthesis traits could stem from their ecological origin, environmental heterogeneity and phylogenetic position (Yan et al., 2023). This variance can influence the community-level response of tropical Andean forests to climate. If photosynthesis traits, such as apparent or Jmax, of a single species are negatively impacted by high temperature, low humidity, or low light availability, a different species may compensate and maintain ecosystem functioning. For example, under environmental conditions similar to the 14 °C site, which is the site closest to the optimal temperature conditions to which many tropical montane species are adapted to, Miconia theizans and Clusia multiflora exhibit high photosynthetic capacity (apparent ) but, because of their low water use efficiency (g1), are strongly affected by the warmer and drier conditions experienced at the 22 °C site (Fig. S1), which is at the warm extreme of the thermal range of these species. In contrast, Quercus humboldtii, maintains a similar photosynthetic capacity (apparent ) at both sites, suggesting that the Andean oak is more tolerant to warming and can help tropical Andean forests be resilient to climate change. Therefore, the combination of species with different photosynthesis traits may positively influence productivity, as high interspecific variance can help maintain stable levels across a wide range of environmental conditions, including variations in air temperature, VPD or solar radiation. Here, we suggest that regional predictions of photosynthesis and carbon dynamics of tropical Andean forests should include interspecific variation in photosynthetic traits and how particular combinations of these traits maintain high photosynthesis rates under changing environmental conditions (Bahar et al., 2017, Fig. 6e).
Surprisingly, intraspecific and interspecific variance of the physiological traits in our data set are of similar magnitudes (σ2 intra = 28 % to σ2 inter = 43 % of total variance). Our parametrization approach, which iteratively searches for the combination of physiological traits (apparent , apparent Jmax and g1), improves prediction of photosynthetic rates. The large difference between the best fitted physiological trait values relative to average observed values suggest that every leaf/tree adjusts their physiology to the experienced environmental conditions to maintain photosynthesis. Therefore, our analysis highlights the importance of field-based measurements and the inclusion of intraspecific variance in physiological traits to be used in vegetation modelling (Maréchaux et al., 2025). Given this variability, an important question arises: how does trait variation influence overall canopy functioning metrics? If trait variability is not explicitly considered, do we risk misrepresenting canopy-level photosynthetic performance? The ability of species to rapidly acclimate to changing conditions suggests a potential compensatory mechanism, but it remains unclear whether this plasticity is sufficient to maintain overall canopy function at similar levels or if significant differences emerge under shifting environmental conditions. Addressing this uncertainty is crucial for refining dynamic vegetation models, as compensatory responses could either buffer or amplify ecosystem-level responses to climate change. The high variability of photosynthetic traits and the ability of species to acclimate to changing conditions may be adaptations to the high environmental variability that occurs in montane forests (Fyllas et al., 2017). Furthermore, intraspecific variance can enhance interspecific variance in terms of maintaining photosynthesis under increased warming (Bassow and Bazzaz, 1997; Zaka et al., 2016). Therefore, the explicit inclusion of intra- and interspecific variance in dynamic vegetation models can improve our ability to understand the response of tropical montane forests to future warming.
We found that the stomatal conductance trait (g1) showed the highest intraspecific variance detected by our approach, suggesting that stomatal control is critical to maintaining photosynthesis under changing environmental conditions. The intraspecific variance of functional traits has been highlighted as an overlooked diversity property that can influence the community response to environmental conditions (Lin et al., 2015). For example, the intraspecific variance of leaf morphological traits such as leaf mass per area has been globally evaluated, showing that intra- and interspecific variance can be similar in magnitude (Anderegg et al., 2018; Siefert et al., 2015). This result aligns with our findings on physiological traits in montane tropical forest; however, measuring intraspecific variance for physiological traits is more challenging due to logistical constraints and the time-consuming measurements that are needed. Our approach, which combines diurnal cycle measurements of photosynthesis that can be easily taken in the field with computational and modelling methods, can help describe the intraspecific variance of tropical montane species.
4.2 Contribution of physiological traits to diurnal variation of leaf photosynthesis
Our results indicate that apparent plays a dominant role in regulating photosynthesis (An) in both montane and lowland tree species, contributing approximately 56 % of simulated An. The contribution of apparent increases when species are subjected to temperature changes (warming for montane species and cooling for lowland species), suggesting that carboxylation capacity is a key driver of Andean Forest responses to climate change. Conversely, apparent Jmax plays a more significant role in regulating An in species growing in their native environments, implying that its influence is linked to local environmental conditions. This raises an important question: will photosynthetic responses to increased temperatures under future climate be primarily driven by apparent as our results suggest, or could apparent Jmax regain dominance through acclimation processes? The fact that apparent Jmax is strongly coupled to the light environment in both montane and lowland species suggests that changes in light availability could modulate its role in photosynthetic regulation (Fyllas et al., 2017). Montane trees, which naturally experience low and fluctuating light conditions, may be more constrained by light interception traits, whereas lowland species, adapted to more stable but lower-intensity light conditions, might rely on different acclimation strategies (Marthews et al., 2012). Therefore, while leaf traits related to light interception appear crucial for in situ performance, their significance under climate warming remains uncertain. If temperature increases lead to shifts in light availability (e.g., through cloud displacement or canopy restructuring), species with traits optimizing light capture could gain a competitive advantage. However, further research is needed to determine whether light interception remains a limiting factor under warming or if apparent -driven biochemical responses will dominate photosynthetic acclimation.
The relative contribution of simulated stomatal conductance (g1) increases in the afternoon, mainly at the 22 °C site, in which Tair and VPD commonly increase with respect to the 14 °C site. This is likely a response to high leaf-to-air vapor pressure deficit, indicating that a rapid stomatal response (g1) is crucial for regulating water loss and gas exchange under conditions of high VPD, helping maintain photosynthetic efficiency and preventing excessive water loss. The high variability of g1 detected by our parametrization approach indicates that the species with high stomatal control ability can persist under the high environmental heterogeneity imposed by tropical highland conditions (Guo et al., 2022). Here, we suggest that the dynamic role of g1 in response to VPD should also be incorporated into models to better predict diurnal and seasonal photosynthetic performance and the differential responses of montane and lowland species to temperature and VPD highlighting the importance of biodiversity in maintaining ecosystem function (Smith et al., 2020).
4.3 Environmental influence on photosynthesis in the tropical highlands
We found differences in the role of environmental drivers on simulated leaf level photosynthesis at the two study sites, 14 °C (2500 m a.s.l.) and 22 °C (1400 m a.s.l.) (Figs. 4 and 5). At the 14 °C site, Photosynthetically Active Radiation (PAR) and air temperature (Tair) have a stronger influence on simulated photosynthesis, whereas at the 22 °C site, atmospheric demand (VPD) plays a more dominant role. This suggests that, at higher elevations, where temperatures are low and VPD is naturally reduced, radiation becomes the primary climatic driver of photosynthesis. Since Tair partly determines VPD, its lower influence at 14 °C may explain why PAR exerts a stronger control on simulated photosynthesis at this site. Tropical montane forests frequently experience low and fluctuating solar radiation due to persistent cloud cover and fog, leading to the adaptation of montane tree species to these conditions (Fyllas et al., 2017). In contrast, lowland species, which are beginning to colonize higher elevations, may compensate for the reduced and variable light by increasing their photosynthetic capacity, primarily through higher apparent values (Bahar et al., 2017; Yan et al., 2023). This raises an important question: how does temperature interact with radiation limitation across different thermal environments? While our findings suggest that PAR limits photosynthesis at high elevations (low Tair), it is also possible that radiation constrains photosynthesis at high temperatures, particularly if warming leads to increased cloud displacement along mountain slopes. If climate change results in increased solar radiation at higher elevations, this could raise leaf temperatures, negatively affecting montane species adapted to cooler conditions while favoring thermophilisation – the upward migration of lowland species (Duque et al., 2015; Fadrique et al., 2018; Cuni-Sanchez et al., 2024).
Our analyses suggest a key role for stomatal conductance (g1), a highly inter and intraspecific variable trait, in mediating the effects of PAR, VPD, and Tair on the diurnal variation of simulated An. Under low values of g1, photosynthesis is mainly influenced by Tair and VPD at both sites. However, the effect of Tair is larger at 14 °C, while VPD has a stronger effect at 22 °C. This suggests that under low stomatal conductance, photosynthesis at high elevations (14 °C site at 2500 m a.s.l.) is primarily constrained by biochemical limitations due to low temperatures, whereas at intermediate elevations (22 °C site at 1400 m a.s.l.), stomatal control dominates through the indirect effect of Tair on VPD. A key mechanism driving these differences may be the role of apparent in controlling gross primary productivity (GPP) responses to temperature (Marthews et al., 2012). At low Tair, biochemical limitations require a higher apparent to sustain high GPP, as enzymatic reactions slow down at colder temperatures. If apparent does not scale accordingly, GPP remains limited by temperature (Marthews et al., 2012). However, maintaining high apparent is metabolically costly, meaning some montane species may instead favor a strategy of lower apparent combined with rapid stomatal responses to optimize carbon gain and water use efficiency. Conversely, under warming conditions, apparent appears less responsive to temperature than apparent Jmax, suggesting that lack of apparent acclimation could negatively impact GPP. While Jmax can respond dynamically to increased temperatures, a relatively unacclimated apparent may impose a biochemical bottleneck, further limiting photosynthetic potential under future warming (Marthews et al., 2012). This introduces two critical processes: (i) the need for a higher apparent at low temperatures to compensate for temperature-related biochemical constraints on GPP, balanced against the metabolic costs of maintaining such high enzymatic activity. (ii) The lower responsiveness of apparent compared to apparent Jmax under warming, which may restrict acclimation potential, making tropical montane species increasingly vulnerable to climate change (Smith et al., 2020). These findings reinforce the necessity of incorporating thermal acclimation of temperature-dependent responses of both apparent and apparent Jmax into models predicting future forest productivity under climate change.
On the other hand, when apparent and apparent Jmax are high (under all g1 cases tested), photosynthesis is primarily controlled by PAR and the influence of VPD is significantly reduced with increasing g1. This suggest that high g1 species with high photosynthetic capacity are less constrained by VPD and temperature, with PAR being the dominant factor. This indicates that species with higher photosynthetic capacities can better exploit available light for photosynthesis, reducing their sensitivity to other environmental stressors. Montane and lowland species have evolved different mechanisms to optimize photosynthesis, reflecting their adaptation to their unique environments. For lowland species, maximising photosynthesis capacity could be an advantageous strategy. However, this strategy is unstable on tropical highlands because of the fluctuating nature of light conditions, due to frequent fog and cloudiness. Therefore, a more favourable, i.e. resource-efficient, photosynthesis strategy under these conditions can be low apparent combined to high stomatal control for rapid response to changing conditions (Bahar et al., 2017; van de Weg et al., 2012). This highlights the importance of understanding the physiological and anatomical features that determine the ability to control stomatal conductance (g1) response under stressful conditions such as tropical mountains for maintaining photosynthesis.
4.4 Recommendations for photosynthesis modelling in montane forests
We found that total photosynthesis in montane forests is related to local conditions (e.g., elevational and climate) but also to species composition at each site (montane and lowland species). Although, we expected the contribution of montane species to be larger at 14 °C and vice versa for lowland species at 22 °C, we found that combining the parametrization of both species groups increases the photosynthesis estimation of montane forests and a reduction in lowlands (Bahar et al., 2017; Cox et al., 2023a; Dusenge et al., 2025). High-elevation conditions can be adverse for lowland species, and their modelled photosynthetic responses likely underestimate actual forest productivity at these sites. This may lead to a misinterpretation of the effects of low solar radiation and persistent cloud cover, as montane tree species tend to perform better under such conditions. Conversely, photosynthesis in lowland forests between 1000 and 1800 m a.s.l. may be overestimated in models that do not account for the presence of montane species. This result highlights the importance of accounting for community composition (e.g., proportion of montane vs. lowland with their corresponding parameters) to model photosynthesis at landscape scale. Common large modelling approaches are restricted to use mean values of physiological parameters and exclude the high observed species level variability. Also, the differential response of each species group to environmental conditions needs to be accounted for in large scale modelling because the response of species to Tair, VPD and PAR varies along elevation and by species adaptations to their origin climates (Yan et al., 2023). Our results demonstrate large variability in g1 and suggest that it is important to incorporate this high variability when modelling stomatal conductance under the heterogeneous environmental conditions of tropical elevational gradients. In sum, we recommend using specific parameterisations for montane and lowland species when modelling montane forest and caution for modelling approaches with respect to using invariant physiological parameters when modelling along complex environmental gradients such as in tropical montane forests as this can obscure photosynthesis response to rapidly changing conditions. However, our findings are based on juvenile trees growing without competition and thus should be interpreted as reflecting potential physiological responses rather than realised ecosystem-scale productivity. Nevertheless, because juveniles often exhibit higher photosynthetic capacity and represent the future cohorts of these forests, they provide valuable insights into the mechanisms that will likely shape mature forest responses to ongoing warming.
The code associated with this study is available on Zenodo at https://doi.org/10.5281/zenodo.20138234 (González-Caro, 2026).
The physiological data that support the findings of this study are openly available in Figshare at https://doi.org/10.6084/m9.figshare.29084276.v1 (Dusenge, 2025) and from https://doi.org/10.5285/60dd0b8f-f0c3-4e30-841b-1c2067052974 (Cox et al., 2023b). The tree growth data are available at NERC EDS Environmental Information Data Centre at https://doi.org/10.5285/c7ce1610-aba3-4a09-bf7c-1b6c774d597a (Restrepo and Mercado, 2024).
The supplement related to this article is available online at https://doi.org/10.5194/bg-23-3347-2026-supplement.
SGC and LMM conceptualized this study. Data were collected by MED, ZR, AJFC, AS, SGC and LMM. LMM, IH and PM with support from AS, DR and ZR wrote the grant that funded the project that funded the work and collected the data. SGC analysed data. SGC drafted the manuscript with significant inputs from LMM, IH and PM. All authors provided inputs to the final version of the manuscript.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
Sebastián González-Caro, Mirindi Eric Dusenge, Zorayda Restrepo, Andrew J. F. Cox, Ian P. Hartley, Patrick Meir, Adriana Sánchez, Daniel Ruiz-Carrascal and Lina M. Mercado acknowledge funding from the UK Natural Environment Research Council (NE/R001928/1). Sebastián González-Caro, Zorayda Restrepo, Ian P. Hartley, Patrick Meir, and Lina M. Mercado acknowledge funding from the UK Natural Environment Research Council (NE/X001172/1.) For the purpose of open access, the author has applied a “Creative Commons Attribution” (CC BY) licence to any Author Accepted Manuscript version arising from this submission.
This research has been supported by the Natural Environment Research Council, Centre for Ecology and Hydrology (grant nos. NE/R001928/1 and NE/X001172/1).
This paper was edited by Kirsten Thonicke and reviewed by David Ellsworth and one anonymous referee.
Anderegg, L. D. L., Berner, L. T., Badgley, G., Sethi, M. L., Law, B. E., and HilleRisLambers, J.: Within-species patterns challenge our understanding of the leaf economics spectrum, Ecol. Lett., 21, 734–744, https://doi.org/10.1111/ele.12945, 2018.
Bahar, N. H. A., Ishida, F. Y., Weerasinghe, L. K., Guerrieri, R., O'Sullivan, O. S., Bloomfield, K. J., Asner, G. P., Martin, R. E., Lloyd, J., Malhi, Y., Phillips, O. L., Meir, P., Salinas, N., Cosio, E. G., Domingues, T. F., Quesada, C. A., Sinca, F., Escudero Vega, A., Zuloaga Ccorimanya, P. P., del Aguila-Pasquel, J., Quispe Huaypar, K., Cuba Torres, I., Butrón Loayza, R., Pelaez Tapia, Y., Huaman Ovalle, J., Long, B. M., Evans, J. R., and Atkin, O. K.: Leaf-level photosynthetic capacity in lowland Amazonian and high-elevation Andean tropical moist forests of Peru, New Phytol., 214, 1002–1018, https://doi.org/10.1111/nph.14079, 2017.
Bassow, S. L. and Bazzaz, F. A.: Intra- and inter-specific variation in canopy photosynthesis in a mixed deciduous forest, Oecologia, 109, 507–515, https://doi.org/10.1007/s004420050111, 1997.
Blonder, B., Escobar, S., Kapás, R. E., and Michaletz, S. T.: Low predictability of energy balance traits and leaf temperature metrics in desert, montane and alpine plant communities, Funct. Ecol., 34, 1882–1897, 2020.
Cox, A. J. F., Hartley, I. P., Meir, P., Sitch, S., Dusenge, M. E., Restrepo, Z., González-Caro, S., Villegas, J. C., Uddling, J., and Mercado, L. M.: Acclimation of photosynthetic capacity and foliar respiration in Andean tree species to temperature change, New Phytol., 238, 2329–2344, https://doi.org/10.1111/nph.18900, 2023a.
Cox, A. J. F., Hartley, I. P., Meir, P., Sitch, S., Dusenge, M. E., Restrepo, Z., González-Caro, S., Villegas, J. C., Uddling, J., and Mercado, L. M.: A–Ci, photosynthetic capacity and dark respiration data along an elevation/temperature gradient in the Colombian Andes, 2019, NERC EDS Environmental Information Data Centre [data set], https://doi.org/10.5285/60dd0b8f-f0c3-4e30-841b-1c2067052974, 2023b.
Cox, A. J. F., González-Caro, S., Meir, P., Hartley, I. P., Restrepo, Z., Villegas, J. C., Sanchez, A., and Mercado, L. M.: Variable thermal plasticity of leaf functional traits in Andean tropical montane forests, Plant Cell Environ., 47, 731–750, https://doi.org/10.1111/pce.14778, 2024.
Cuni-Sanchez, A., Sullivan, M. J. P., Platts, P. J., Lewis, S. L., Marchant, R., Imani, G., Hubau, W., Abiem, I., Adhikari, H., Albrecht, T., Altman, J., Amani, C., Aneseyee, A. B., Avitabile, V., Banin, L., Batumike, R., Bauters, M., Beeckman, H., Begne, S. K., Bennett, A. C., Bitariho, R., Boeckx, P., Bogaert, J., Bräuning, A., Bulonvu, F., Burgess, N. D., Calders, K., Chapman, C., Chapman, H., Comiskey, J., de Haulleville, T., Decuyper, M., DeVries, B., Dolezal, J., Droissart, V., Ewango, C., Feyera, S., Gebrekirstos, A., Gereau, R., Gilpin, M., Hakizimana, D., Hall, J., Hamilton, A., Hardy, O., Hart, T., Heiskanen, J., Hemp, A., Herold, M., Hiltner, U., Horak, D., Kamdem, M. N., Kayijamahe, C., Kenfack, D., Kinyanjui, M. J., Klein, J., Lisingo, J., Lovett, J., Lung, M., Makana, J. R., Malhi, Y., Marshall, A., Martin, E. H., Mitchard, E. T. A., Morel, A., Mukendi, J. T., Muller, T., Nchu, F., Nyirambangutse, B., Okello, J., Peh, K. S. H., Pellikka, P., Phillips, O. L., Plumptre, A., Qie, L., Rovero, F., Sainge, M. N., Schmitt, C. B., Sedlacek, O., Ngute, A. S. K., Sheil, D., Sheleme, D., Simegn, T. Y., Simo-Droissart, M., Sonké, B., Soromessa, T., Sunderland, T., Svoboda, M., Taedoumg, H., Taplin, J., Taylor, D., Thomas, S. C., Timberlake, J., Tuagben, D., Umunay, P., Uzabaho, E., Verbeeck, H., Vleminckx, J., Wallin, G., Wheeler, C., González-Caro, S., Dusenge, M. E., Restrepo, Z., Cox, A. J. F., Hartley, I. P., Meir, P., Sánchez, A., Ruiz-Carrascal, D., and Mercado, L. M.: High aboveground carbon stock of African tropical montane forests, Nature, 596, 536–542, https://doi.org/10.1038/s41586-021-03728-4, 2021.
Cuni-Sanchez, A., Martin, E. H., Uzabaho, E., Ngute, A. S. K., Bitariho, R., Kayijamahe, C., Marshall, A. R., Mohamed, N. A., Mseja, G. A., Nkwasibwe, A., Rovero, F., Sheil, D., Tinkasimire, R., Tumugabirwe, L., Feeley, K. J., and Sullivan, M. J. P.: Evidence of thermophilization in Afromontane forests, Nat. Commun., 15, https://doi.org/10.1038/s41467-024-48520-w, 2024.
Duque, A., Stevenson, P. R., and Feeley, K. J.: Thermophilization of adult and juvenile tree communities in the northern tropical Andes, P. Natl. Acad. Sci. USA, 112, 10744–10749, https://doi.org/10.1073/pnas.1506570112, 2015.
Duque, A., Peña, M. A., Cuesta, F., González-Caro, S., Kennedy, P., Phillips, O. L., Calderón-Loor, M., Blundo, C., Carilla, J., Cayola, L., Farfán-Ríos, W., Fuentes, A., Grau, R., Homeier, J., Loza-Rivera, M. I., Malhi, Y., Malizia, A., Malizia, L., Martínez-Villa, J. A., Myers, J. A., Osinaga-Acosta, O., Peralvo, M., Pinto, E., Saatchi, S., Silman, M., Tello, J. S., Terán-Valdez, A., and Feeley, K. J.: Mature Andean forests as globally important carbon sinks and future carbon refuges, Nat. Commun., 12, 1–10, https://doi.org/10.1038/s41467-021-22459-8, 2021.
Dusenge, M. E.: Dataset for the paper: “Unexpected large photosynthetic thermal plasticity of montane Andean trees”, figshare [data set], https://doi.org/10.6084/m9.figshare.29084276.v1, 2025.
Dusenge, M. E., Wittemann, M., Mujawamariya, M., Ntawuhiganayo, E. B., Zibera, E., Ntirugulirwa, B., Way, D. A., Nsabimana, D., Uddling, J., and Wallin, G.: Limited thermal acclimation of photosynthesis in tropical montane tree species, Global Change Biol., 27, 4860–4878, https://doi.org/10.1111/gcb.15790, 2021.
Dusenge, M. E., González-Caro, S., Restrepo, Z., Gardner, A., Meir, P., Hartley, I. P., Sitch, S., Sanchez, A., Villegas, J. C., and Mercado, L. M.: Unexpected Large Photosynthetic Thermal Plasticity of Montane Andean Trees, Global Change Biol., 31, e70266, https://doi.org/10.1111/gcb.70266, 2025.
Duursma, R. A.: Plantecophys – An R package for analysing and modelling leaf gas exchange data, PLoS ONE, 10, 1–13, https://doi.org/10.1371/journal.pone.0143346, 2015.
Fadrique, B., Báez, S., Duque, Á., Malizia, A., Blundo, C., Carilla, J., Osinaga-Acosta, O., Malizia, L., Silman, M., Farfán-Ríos, W., Malhi, Y., Young, K. R., Cuesta C, F., Homeier, J., Peralvo, M., Pinto, E., Jadan, O., Aguirre, N., Aguirre, Z., and Feeley, K. J.: Widespread but heterogeneous responses of Andean forests to climate change, Nature, 564, 207–212, https://doi.org/10.1038/s41586-018-0715-9, 2018.
Farquhar, G. D., von Caemmerer, S. V., and Berry, J. A.: A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species, Planta, 149, 78–90, 1980.
Fisher, R. A., Koven, C. D., Anderegg, W. R. L., Christoffersen, B. O., Dietze, M. C., Farrior, C. E., Holm, J. A., Hurtt, G. C., Knox, R. G., Lawrence, P. J., Lichstein, J. W., Longo, M., Matheny, A. M., Medvigy, D., Muller-Landau, H. C., Powell, T. L., Serbin, S. P., Sato, H., Shuman, J. K., Smith, B., Trugman, A. T., Viskari, T., Verbeeck, H., Weng, E., Xu, C., Xu, X., Zhang, T., and Moorcroft, P. R.: Vegetation demographics in Earth System Models: A review of progress and priorities, Global Change Biol., 24, 35–54, https://doi.org/10.1111/gcb.13910, 2018.
Fyllas, N. M., Bentley, L. P., Shenkin, A., Asner, G. P., Atkin, O. K., Díaz, S., Enquist, B. J., Farfan-Rios, W., Gloor, E., Guerrieri, R., Huasco, W. H., Ishida, Y., Martin, R. E., Meir, P., Phillips, O., Salinas, N., Silman, M., Weerasinghe, L. K., Zaragoza-Castells, J., and Malhi, Y.: Solar radiation and functional traits explain the decline of forest primary productivity along a tropical elevation gradient, Ecol. Lett., 20, 730–740, https://doi.org/10.1111/ele.12771, 2017.
Girardin, C. A. J., Malhi, Y., Aragao, L. E. O. C., Mamani, M., Huaraca Huasco, W., Durand, L., Feeley, K. J., Rapp, J., Silva-Espejo, J. E., Silman, M., Salinas, N., and Whittaker, R. J.: Net primary productivity allocation and cycling of carbon along a tropical forest elevational transect in the Peruvian Andes, Global Change Biol., 16, 3176–3192, 2010.
González-Caro, S.: Sensitivity of a mechanistic photosynthesis model to tropical Andean species and environments. In Biogeoscience, Zenodo [code], https://doi.org/10.5281/zenodo.20138234, 2026.
González-Caro, S., Duque, Á., Feeley, K. J., Cabrera, E., Phillips, J., Ramirez, S., and Yepes, A.: The legacy of biogeographic history on the composition and structure of Andean forests, Ecology, 101, e03131, https://doi.org/10.1002/ecy.3131, 2020.
González-Caro, S., Tello, J. S., Myers, J. A., Feeley, K., Blundo, C., Calderón-Loor, M., Carilla, J., Cayola, L., Cuesta, F., Farfán, W., Fuentes, A. F., Garcia-Cabrera, K., Grau, R., Idarraga, Á., Loza, M. I., Malhi, Y., Malizia, A., Malizia, L., Osinaga-Acosta, O., Pinto, E., Salinas, N., Silman, M., Terán-Valdéz, A., and Duque, Á.: Historical Assembly of Andean Tree Communities, Plants, 12, https://doi.org/10.3390/plants12203546, 2023.
Guo, Z., Yan, Z., Majcher, B. M., Lee, C. K. F., Zhao, Y., Song, G., Wang, B., Wang, X., Deng, Y., Michaletz, S. T., Ryu, Y., Ashton, L. A., Lam, H. M., Wong, M. S., Liu, L., and Wu, J.: Dynamic biotic controls of leaf thermoregulation across the diel timescale, Agr. Forest Meteorol., 315, 108827, https://doi.org/10.1016/j.agrformet.2022.108827, 2022.
Harper, A. B., Wiltshire, A. J., Cox, P. M., Friedlingstein, P., Jones, C. D., Mercado, L. M., Sitch, S., Williams, K., and Duran-Rojas, C.: Vegetation distribution and terrestrial carbon cycle in a carbon cycle configuration of JULES4.6 with new plant functional types, Geosci. Model Dev., 11, 2857–2873, https://doi.org/10.5194/gmd-11-2857-2018, 2018.
Knauer, J., Cuntz, M., Smith, B., Canadell, J. G., Medlyn, B. E., Bennett, A. C., Caldararu, S., and Haverd, V.: Higher global gross primary productivity under future climate with more advanced representations of photosynthesis, Sci. Adv., 9, 24–28, https://doi.org/10.1126/SCIADV.ADH9444, 2023.
Kumarathunge, D. P., Medlyn, B. E., Drake, J. E., Tjoelker, M. G., Aspinwall, M. J., Battaglia, M., Cano, F. J., Carter, K. R., Cavaleri, M. A., Cernusak, L. A., Chambers, J. Q., Crous, K. Y., De Kauwe, M. G., Dillaway, D. N., Dreyer, E., Ellsworth, D. S., Ghannoum, O., Han, Q., Hikosaka, K., Jensen, A. M., Kelly, J. W. G., Kruger, E. L., Mercado, L. M., Onoda, Y., Reich, P. B., Rogers, A., Slot, M., Smith, N. G., Tarvainen, L., Tissue, D. T., Togashi, H. F., Tribuzy, E. S., Uddling, J., Vårhammar, A., Wallin, G., Warren, J. M., and Way, D. A.: Acclimation and adaptation components of the temperature dependence of plant photosynthesis at the global scale, New Phytol., 222, 768–784, https://doi.org/10.1111/nph.15668, 2019.
Lin, Y. S., Medlyn, B. E., Duursma, R. A., Prentice, I. C., Wang, H., Baig, S., Eamus, D., De Dios, V. R., Mitchell, P., Ellsworth, D. S., De Beeck, M. O., Wallin, G., Uddling, J., Tarvainen, L., Linderson, M. L., Cernusak, L. A., Nippert, J. B., Ocheltree, T. W., Tissue, D. T., Martin-StPaul, N. K., Rogers, A., Warren, J. M., De Angelis, P., Hikosaka, K., Han, Q., Onoda, Y., Gimeno, T. E., Barton, C. V. M., Bennie, J., Bonal, D., Bosc, A., Löw, M., Macinins-Ng, C., Rey, A., Rowland, L., Setterfield, S. A., Tausz-Posch, S., Zaragoza-Castells, J., Broadmeadow, M. S. J., Drake, J. E., Freeman, M., Ghannoum, O., Hutley, L. B., Kelly, J. W., Kikuzawa, K., Kolari, P., Koyama, K., Limousin, J. M., Meir, P., Da Costa, A. C. L., Mikkelsen, T. N., Salinas, N., Sun, W., and Wingate, L.: Optimal stomatal behaviour around the world, Nat. Clim. Change, 5, 459–464, https://doi.org/10.1038/nclimate2550, 2015.
Malhi, Y., Girardin, C. A., Goldsmith, G. R., Doughty, C. E., Salinas, N., Metcalfe, D. B., Huaraca Huasco, W., Silva-Espejo, J. E., del Aguilla-Pasquell, J., Farfán Amézquita, F., Aragão, L. E. O. C., Guerrieri, R., Ishida, F. Y., Bahar, N. H. A., Farfan-Rios, W., Phillips, O. L., Meir, P., and Silman, M.: The variation of productivity and its allocation along a tropical elevation gradient: a whole carbon budget perspective, New Phytol., 214, 1019–1032, 2017.
Maréchaux, I., Fischer, F. J., Schmitt, S., and Chave, J.: TROLL 4.0: representing water and carbon fluxes, leaf phenology, and intraspecific trait variation in a mixed-species individual-based forest dynamics model – Part 1: Model description, Geosci. Model Dev., 18, 5143–5204, https://doi.org/10.5194/gmd-18-5143-2025, 2025.
Marthews, T. R., Malhi, Y., Girardin, C. A. J., Silva Espejo, J. E., Aragão, L. E. O. C., Metcalfe, D. B., Rapp, J. M., Mercado, L. M., Fisher, R. A., Galbraith, D. R., Fisher, J. B., Salinas-Revilla, N., Friend, A. D., Restrepo-Coupe, N., and Williams, R. J.: Simulating forest productivity along a neotropical elevational transect: Temperature variation and carbon use efficiency, Global Change Biol., 18, 2882–2898, https://doi.org/10.1111/j.1365-2486.2012.02728.x, 2012.
Martínez, R., Ruiz, D., Andrade, M., Blacutt, L., and Pabón, D.: Synthesis of the Climate of the Tropical Andes BT – Climate Change and Biodiversity in the Tropical Andes, Climate Change and Biodiversity in the Tropical Andes, Inter-American Institute for Global Change Research, ISBN 978-92-95046-03-0, 97–109, 2011.
Medlyn, B. E., Dreyer, E., Ellsworth, D., Forstreuter, M., Harley, P. C., Kirschbaum, M. U. F., Le Roux, X., Montpied, P., Strassemeyer, J., Walcroft, A., Wang, K., and Loustau, D.: Temperature response of parameters of a biochemically based model of photosynthesis. II. A review of experimental data, Plant Cell Environ., 25, 1167–1179, https://doi.org/10.1046/j.1365-3040.2002.00891.x, 2002.
Medlyn, B. E., Duursma, R. A., Eamus, D., Ellsworth, D. S., Prentice, I. C., Barton, C. V. M., Crous, K. Y., De Angelis, P., Freeman, M., and Wingate, L.: Reconciling the optimal and empirical approaches to modelling stomatal conductance, Global Change Biol., 17, 2134–2144, https://doi.org/10.1111/j.1365-2486.2010.02375.x, 2011.
Mercado, L. M., Medlyn, B. E., Huntingford, C., Oliver, R. J., Clark, D. B., Sitch, S., Zelazowski, P., Kattge, J., Harper, A. B., and Cox, P. M.: Large sensitivity in land carbon storage due to geographical and temporal variation in the thermal response of photosynthetic capacity, New Phytol., 218, 1462–1477, https://doi.org/10.1111/nph.15100, 2018.
Montaño-Centellas, F., Fuentes, A. F., Cayola, L., Macía, M. J., Arellano, G., Loza, M. I., Nieto-Ariza, B., and Tello, J. S.: Elevational range sizes of woody plants increase with climate variability in the Tropical Andes, J. Biogeogr., 51, 814–826, https://doi.org/10.1111/jbi.14783, 2024.
Mujawamariya, M., Wittemann, M., Dusenge, M. E., Manishimwe, A., Ntirugulirwa, B., Zibera, E., Nsabimana, D., Wallin, G., and Uddling, J.: Contrasting warming responses of photosynthesis in early- and late-successional tropical trees, Tree Physiol., 43, 1104–1117, https://doi.org/10.1093/treephys/tpad035, 2023.
Myers, N., Mittermeier, R. A., Mittermeier, C. G., Da Fonseca, G. A., and Kent, J.: Biodiversity hotspots for conservation priorities, Nature, 403, 853-=–858, 2000.
Myers, N., Russell, M. A., Mittermeier, C. G., Fonseca, G. A. B. da, Kent, J., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. B., and Kent, J.: Biodiversity hotspots for conservation priorities Norman, Nature, 403, 854–858, 2010.
Oliver, R. J., Mercado, L. M., Clark, D. B., Huntingford, C., Taylor, C. M., Vidale, P. L., McGuire, P. C., Todt, M., Folwell, S., Shamsudheen Semeena, V., and Medlyn, B. E.: Improved representation of plant physiology in the JULES-vn5.6 land surface model: photosynthesis, stomatal conductance and thermal acclimation, Geosci. Model Dev., 15, 5567–5592, https://doi.org/10.5194/gmd-15-5567-2022, 2022.
Poulter, B., Ciais, P., Hodson, E., Lischke, H., Maignan, F., Plummer, S., and Zimmermann, N. E.: Plant functional type mapping for earth system models, Geosci. Model Dev., 4, 993–1010, https://doi.org/10.5194/gmd-4-993-2011, 2011.
Puy, A., Becker, W., Lo Piano, S., and Saltelli, A.: a Comprehensive Comparison of Total-Order Estimators for Global Sensitivity Analysis, Int. J. Uncertain. Quant., 12, 1–18, https://doi.org/10.1615/Int.J.UncertaintyQuantification.2021038133, 2022.
Restrepo, Z. and Mercado, L. M.: Leaf structural traits and allometry of juvenile trees across temperature treatments in an experimental plantation in the Colombian Andes, 2019–2022, NERC EDS Environmental Information Data Centre [data set], https://doi.org/10.5285/c7ce1610-aba3-4a09-bf7c-1b6c774d597a, 2024.
Siefert, A., Violle, C., Chalmandrier, L., Albert, C. H., Taudiere, A., Fajardo, A., Aarssen, L. W., Baraloto, C., Carlucci, M. B., Cianciaruso, M. V., de L. Dantas, V., de Bello, F., Duarte, L. D. S., Fonseca, C. R., Freschet, G. T., Gaucherand, S., Gross, N., Hikosaka, K., Jackson, B., Jung, V., Kamiyama, C., Katabuchi, M., Kembel, S. W., Kichenin, E., Kraft, N. J. B., Lagerström, A., Bagousse-Pinguet, Y. Le, Li, Y., Mason, N., Messier, J., Nakashizuka, T., Overton, J. M., Peltzer, D. A., Pérez-Ramos, I. M., Pillar, V. D., Prentice, H. C., Richardson, S., Sasaki, T., Schamp, B. S., Schöb, C., Shipley, B., Sundqvist, M., Sykes, M. T., Vandewalle, M., and Wardle, D. A.: A global meta-analysis of the relative extent of intraspecific trait variation in plant communities, Ecol. Lett., 18, 1406–1419, https://doi.org/10.1111/ele.12508, 2015.
Smith, M. N., Taylor, T. C., van Haren, J., Rosolem, R., Restrepo-Coupe, N., Adams, J., Wu, J., de Oliveira, R. C., da Silva, R., de Araujo, A. C., de Camargo, P. B., Huxman, T. E., and Saleska, S. R.: Empirical evidence for resilience of tropical forest photosynthesis in a warmer world, Nat. Plants, 6, 1225–1230, https://doi.org/10.1038/s41477-020-00780-2, 2020.
Smith, N. G., Keenan, T. F., Colin Prentice, I., Wang, H., Wright, I. J., Niinemets, Ü., Crous, K. Y., Domingues, T. F., Guerrieri, R., Yoko Ishida, F., Kattge, J., Kruger, E. L., Maire, V., Rogers, A., Serbin, S. P., Tarvainen, L., Togashi, H. F., Townsend, P. A., Wang, M., Weerasinghe, L. K., and Zhou, S.-X.: Global photosynthetic capacity is optimized to the environment, Ecol. Lett., 22, 506–517, https://doi.org/10.1111/ele.13210, 2019.
van de Weg, M. J., Meir, P., Grace, J., and Ramos, G. D.: Photosynthetic parameters, dark respiration and leaf traits in the canopy of a Peruvian tropical montane cloud forest, Oecologia, 168, 23–34, https://doi.org/10.1007/s00442-011-2068-z, 2012.
van de Weg, M. J., Meir, P., Williams, M., Girardin, C., Malhi, Y., Silva-Espejo, J., and Grace, J.: Gross Primary Productivity of a High Elevation Tropical Montane Cloud Forest, Ecosystems, 17, 751–764, https://doi.org/10.1007/s10021-014-9758-4, 2014.
Yan, Z., Sardans, J., Peñuelas, J., Detto, M., Smith, N. G., Wang, H., Guo, L., Hughes, A. C., Guo, Z., Lee, C. K. F., Liu, L., and Wu, J.: Global patterns and drivers of leaf photosynthetic capacity: The relative importance of environmental factors and evolutionary history, Global Ecol. Biogeogr., 32, 668–682, https://doi.org/10.1111/geb.13660, 2023.
Zaka, S., Frak, E., Julier, B., Gastal, F., and Louarn, G.: Intraspecific variation in thermal acclimation of photosynthesis across a range of temperatures in a perennial crop, AoB Plants, 8, https://doi.org/10.1093/aobpla/plw035, 2016.