the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 13 Feb 2026
| 13 Feb 2026
Vulnerability of soil organic carbon in Amazonian Podsols to changes in environmental conditions
Claire Chenu
Valérie Pouteau
André Soro
Kevin Potard
Célia R. Montes
Patricia Merdy
Adolpho J. Melfi
Yves Lucas
It has recently been shown that the C stocks in Amazonian podzols are very large. They are much larger than was previously thought, particularly in the Bh horizon, which has been estimated to contain in excess of 10 Pg C for Amazonia alone. It is predicted that changes in the regional climate will result in a drier soil moisture regime, which may affect the C dynamics in these generally waterlogged soils. In order to determine the vulnerability to decomposition of the organic C contained in the Amazonian podzols as a result of environmental changes, we established a series of incubation experiments in which the effects of different environmental factors were measured. The direct effect of drier soil moisture regimes was tested by incubating undisturbed cores from the Bh horizon at a range of matric potentials. Contrary to what is usually found in soils, no significant difference in mineralisation was found among matric potentials, suggesting that other factors control microbial mineralisation of this organic C. In a second series of incubations, the effect of nitrogen additions, of anoxic conditions and of labile C substrate additions were also tested on undisturbed cores of the Bh horizon. Samples incubated under oxic conditions produced more than twice as much CO2 as samples incubated under anoxic conditions, whilst the mineralisation rates of samples incubated under oxic conditions with the addition of N increased more than four-fold relative to the anoxic samples. The addition of labile C did not have a significant effect on C mineralisation. The data suggest that the large pool of C in Amazonian Podzols may be vulnerable to increases in N and O2 availability.
- Article
(522 KB) - Full-text XML
-
Supplement
(1100 KB) - BibTeX
- EndNote




Hydromorphic Podzols are widespread in equatorial regions (Bernoux et al., 2002; Quesada et al., 2011). They are characterised by a deep sandy horizon on top of clayey horizons. A small portion of the dissolved organic matter from the upper organic horizon moves freely through the sandy horizon and accumulates at the transition with the clay horizon below, forming a deep Bh horizon that can reach thicknesses of several meters (Montes et al., 2011; Sierra et al., 2013; Doupoux et al., 2017). In the Amazon basin, the majority of the dissolved organic carbon is transferred to the Amazon river via a perched water-table in the sandy E horizon above the impermeable Bh horizon (Doupoux et al., 2017). Despite the C loss to the river network, the C stocks in the Amazonian Podzols have been estimated to exceed 13 Pg C, the majority of which is contained in the deep Bh horizons (Montes et al., 2011). This C represents a significant portion of the C stored in the Amazon basin: the total woody biomass of Amazonian forests having been estimated to be between 121 and 126 Pg C (Malhi et al., 2006). Furthermore, 14C dating of the Bh horizon C suggests that the Bh C is very old, reaching ages of up to 25 thousand years (Doupoux et al., 2017). The Bh horizons are also characterised by organic matter with ratios that can be exceptionally high, with values sometimes exceeding 80 (Montes et al., 2023), and by the fact that they are cemented and relatively impermeable (Sierra et al., 2013; Montes et al., 2023). The E horizon above it is therefore generally waterlogged, thus preventing oxygen from penetrating down the soil profile to the Bh horizon.
The vulnerability of this C to changes in environmental conditions is still poorly characterised. The 14C ages in the Bh horizons suggest that it is very stable and resistant to decomposition. However, this does not mean that it is not vulnerable to decomposition if environmental conditions change. Old soil C can be mineralised quite rapidly, as was shown, for example, by Fontaine et al. (2007): the simple addition of cellulose to subsoil samples of a Cambisol stimulated the mineralisation of old organic C with an apparent 14C age of 2500 years, a phenomenon known as the “priming effect”. The decomposition of old, millennial organic C was shown to be as responsive to warming as fast cycling C in high latitude soils (Vaughn and Torn, 2019). These data suggest, therefore, that old organic C may not be intrinsically resistant to decomposition, but rather that it is not decomposed under the prevailing conditions.
Regional climate models, which downscale global climate projections to regions of interest, all predict reductions in precipitation levels and longer or more frequent periods of drought in the Amazon region (Avila-Diaz et al., 2020) and, indeed, the Amazon experienced record-breaking droughts in 2023 and 2024 (Marengo et al., 2024). A significant potential consequence of prolonged periods of drought is that the perched water table above the Bh horizon could dry out, thus leading to its aeration. It is well established that decomposition rates in aerobic conditions are far greater than those in anoxic conditions (Linn and Doran, 1984; Moyano et al., 2012), so we might expect significant increases in Bh horizon mineralisation rates subsequent to such changes in conditions.
The projected climate changes, compounded by deforestation and fires, may also lead to changes in the structure and composition of the forest (Esquivel-Muelbert et al., 2019; Flores et al., 2024) and, ultimately, to forest dieback (Boulton et al., 2022; Flores et al., 2024). There are already precursor signs of resilience loss in the Amazon forest related to reductions in mean annual precipitation and human land-use (Boulton et al., 2022). Forests on hydromorphic podzols are particularly vulnerable to dieback due to their shallow rooting depth, which is probably a consequence of the lack of nutrients in the E horizon, due to its very low exchange capacity, as well as the mostly waterlogged conditions. According to Sierra et al. (2013), the high bulk density of this E horizon below the organic horizon may also play a role. Such dramatic changes in forest dynamics are known to alter element cycling and potentially result in nutrient losses, including N, from the vegetation and the surface organic horizon to the soil horizons below (Xiong et al., 2011). The very high ratio of the Bh horizon organic matter suggests that the mineralisation of this organic C is constrained by N availability and increases in N flux from overlaying horizons, due to the decomposition of dead biomass or increased N deposition (Galloway et al., 2008), may unlock the Bh horizon C. The death of plant biomass and its subsequent decomposition is also likely to release significant amounts of labile organic matter which could stimulate the mineralisation of the Bh horizon organic matter through a priming effect.
Previous studies have suggested that large quantities of C could be released to the atmosphere from these Bh horizons under certain conditions (Sierra et al., 2013; Montes et al., 2023). However, in these studies the structural integrity of samples was not preserved, which is likely to have allowed far greater oxygenation of the samples than would occur under natural circumstances. Furthermore, the disruption of the physical structure of soils is known to stimulate the mineralisation of organic C (Rovira and Greacen, 1957; Salomé et al., 2010).
Despite the Bh carbon pool being very large, its sensitivity to individual environmental drivers remains poorly constrained under realistic physical conditions. Here, we incubated undisturbed soil cores in order to obtain more realistic estimations of the vulnerability of organic C in hydromorphic Podzols, particularly that in the Bh horizon, to a range of potential future disruptions to the present environmental conditions. The conditions tested were changes in moisture status, and in O2 and N availability. We also tested the effect of the addition of a cocktail of labile organic molecules that sought to mimic the arrival of soluble, labile organic matter from the soil surface.
The hypotheses were fourfold. The first was that reductions in moisture content from saturation to a matric potential of approximately −31.6 hPa would result in increases in CO2 emissions, due to increased O2 availability in the pore space (Moyano et al., 2012; Sierra et al., 2017), but that further decreases in moisture content would result in lower CO2 emissions, due to reductions in the diffusion of C-substrates towards enzymes or the diffusion of enzymes towards insoluble C-substrates (Davidson et al., 2014). The second hypothesis was that anoxic conditions were responsible for the slow decomposition rates (Sierra et al., 2017; Davidson et al., 2014) and that decomposition would be stimulated by increases in O2 levels. The third hypothesis was that decomposition is N limited, as indicated by the characteristically high C:N ratios of Podzol Bh horizons (Montes et al., 2023) and, therefore, that the addition of mineral N would stimulate decomposition. The fourth and final hypothesis was that the addition of readily available sources of C would result in a priming effect (Fontaine et al., 2007) that releases CO2 from Bh horizon organic matter.
Samples were taken from three sites circa one hundred meters apart in the region of Cabeça do Cachorro, Amazonas state, Brazil, near the town of São Gabriel da Cachoeira (Fig. S1 in the Supplement). The profiles at the three sites were typical Amazonian Podzols. They were made up of a waterlogged O horizon of about 15 cm, an E horizon that was also waterlogged and slightly less than a meter thick, a silt-loam Bh horizon that was slightly over a meter thick underneath which there was a C horizon. Bulk soil and undisturbed soil cores were collected from the OH, E, Bh and C horizons to a depth of 3 m using an augur inserted into a metal tube that was used to prevent the sandy, waterlogged E horizon from collapsing into the bore hole. This sampling procedure was necessary due to the perched water table above the Bh horizon. The samples were placed in medical sample containers and closed. They were thus maintained in anoxic conditions prior to use. One undisturbed sample was taken from each site x horizon per incubation treatment (see below) and for the establishment of moisture release characteristics, resulting in 10 undisturbed samples being taken from each horizon at each site, and a total of 120 samples. Total C and N contents of the soils were determined by elemental analysis, and pH measurements were carried out in a 1:5 soil : water mixture.
Soil moisture retention curves were established on three replicate samples from each horizon using a suction table and ceramic pressure plates (Eijkelkamp). All samples used in the subsequent incubation experiments were adjusted to the desired matric potential using a suction table and pressure plates.
Two microcosm incubation experiments were set up in order to measure CO2 emissions from samples in response to changes in environmental conditions. The first incubation measured the response to differences in moisture content in O, E, Bh and C horizons. This incubation lasted 68 d. The second incubation measured changes in CO2 emission in response to O2, mineral N or substrate-C availability in the Bh horizon and lasted for 72 d. Both sets of incubations were carried out at 28 °C in the dark. There were three replicate microcosms for every treatment, resulting in a total of 60 microcosms (4 horizons × 5 moisture contents × 3 replicates) for the first incubation and 12 microcosms (4 treatments × 3 replicates) for the second incubation. In both incubation experiments the samples were placed on sample holders in 1 L air-tight jars that were sealed with rubber gaskets and firmly closed with spring-lock catches. The glass lids of the jars were fitted with a septum that allowed for headspace sampling. The concentration of CO2 in the headspace of sample microcosms was analysed on 18 and 23 occasions during the first and second incubations, respectively. The headspace of the microcosms was flushed with CO2-free air at regular intervals to ensure that the conditions did not become anoxic, except for the anoxic treatment in the second incubation experiment (see below). The moisture content of the samples was adjusted gravimetrically when necessary. The CO2 concentrations in the microcosm headspaces were determined by gas chromatography (Agilent 3000A) and the isotopic signature of the CO2 by gas chromatography coupled to an Isochrome III isotope ratio mass spectrometer (Micromass-GVI Optima). The amount of soil organic C and 13C-labelled substrate mineralised in the samples that received a cocktail of 13C-labelled substrates (see below) was determined by isotopic mass balance (e.g. Ruamps et al., 2011).
In the first incubation experiment, the three replicate samples from each horizon were incubated at matric potentials of 0, −5, −31.6, −316 or −1585 kPa. The matric potentials were chosen in order to have a broad range of potentials from saturation (0 kPa) to the permanent wilting point (−1585 kPa). The range of matric potentials was centered on −31.6 kPa because respiration maxima are generally reached at approximately this potential (Moyano et al., 2012). In the second experiment a series of treatments were imposed in order to determine whether microbial decomposition of the Bh horizon organic matter was limited by O2, N or energy availability. The samples were first pre-incubated for two weeks under oxic conditions in order to ensure that there were no differences in soil respiration among the samples chosen for the different treatments. After the pre-incubation, four treatments were imposed: an anoxic treatment, an O2 treatment, an O2 + N treatment and an O2 + simple organic substrate treatment.
The anoxic treatment was established by replacing the microcosm headspace with N2. In the oxic treatments samples were incubated in the presence of ambient O2 levels, alone or with the addition of N (1.6 mg g−1 soil, in order to bring the ratio of the soil to approximately 20) or a cocktail of substrates (15 mg g−1 soil C). The cocktail was made up of 13C-labelled 30 % glucose, 50 % vanillin and 20 % pyruvic acid. These proportions were chosen to approximate the soil solution of forest soils (Kaiser et al., 2001). On the 34th day of incubation, a second addition of N (half the amount previously added) and of the substrate cocktail was carried out. The other two treatments received water. The matric potential of the samples was set at −1585 kPa.
Due to the difficulty and expense of the sampling exercise, there were not a sufficient number of undisturbed cores for a fully factorial experimental design (i.e. there were no treatment combinations) and, therefore, treatment and horizon differences in the cumulative amount of CO2 evolved from the soil samples during the incubations were tested by one-way ANOVA. Data were log-transformed prior to analysis, where necessary. Differences in soil properties among horizons were also tested by one-way ANOVA. In order to estimate the annual CO2 flux from the Bh horizon of Amazonian Podzols to the atmosphere under the different conditions tested here, a first-order decay model with one pool (Manzoni et al., 2012) was fitted to the cumulative CO2 emission curves (Eq. 1):
where a is the pool of mineralisable C, α is the rate at which the organic C is mineralised and t is time. The model fitting was done using the nls command in R (R Core Team, 2022).
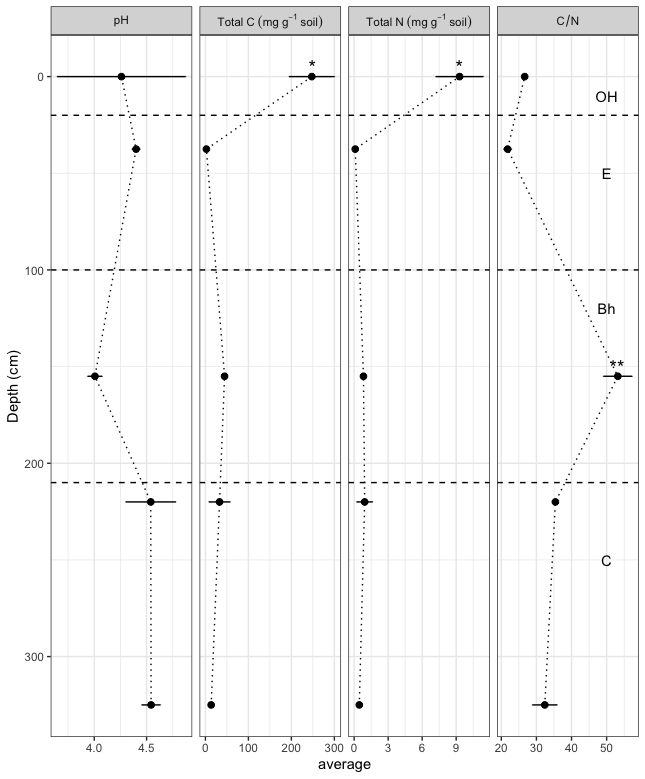
Figure 1Properties of different horizons of the Podzols. The bars indicate the standard deviation of the mean where it is larger than the size of the symbols. Stars indicate horizons with values that are significantly (* <0.05, ** <0.01) higher than in other horizons.
The soil properties were typical of Amazonian Podzols (Montes et al., 2011; Sierra et al., 2013; Doupoux et al., 2017). The pH was acidic (<4.6) throughout the profile, without showing any significant differences among horizons (Fig. 1). The C and N contents ranged from 2 to 248 and from 0.1 to 9.3 mg g−1 soil, respectively, with significantly higher values for both variables in the OH horizon (Fig. 1). The ratios were all >20 but the Bh horizon showed by far the highest ratio at 53, which was significantly higher than in the other horizons. No other statistically significant differences were observed.
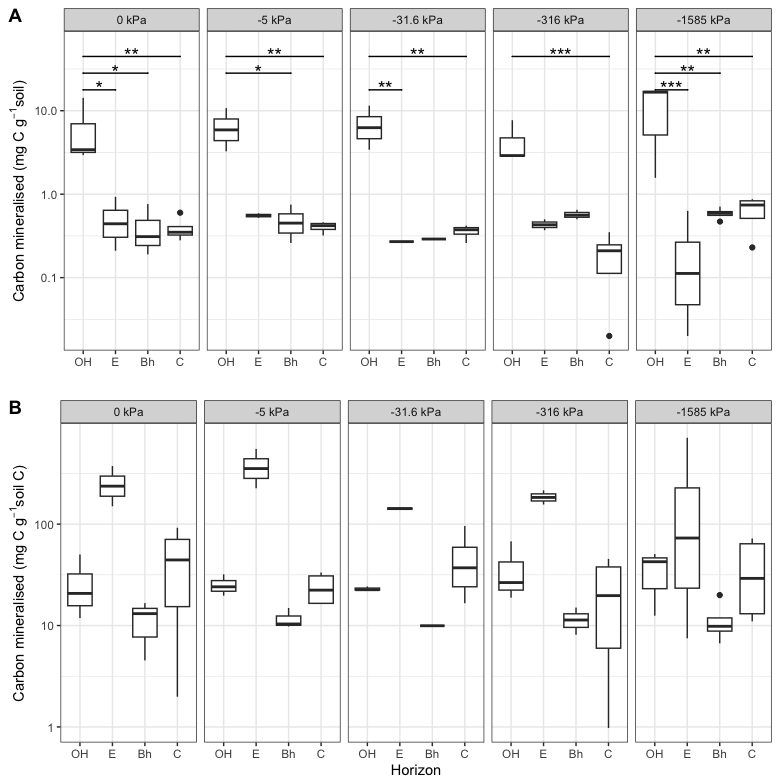
Figure 2Total C mineralisation in the different Podzol horizons at different matric potentials during the 68 d incubation. Mineralisation is expressed per g soil (A) and per g soil C (B). Note that the y axes are in log scale. Across all matric potentials, the carbon mineralised (per g soil) from the OH horizon was significantly (P<0.001) higher than in all other horizons. Differences within each matric potential are shown in the graph (* <0.05, ** <0.01, *** <0.001). The carbon mineralised per g soil C from the E horizon was significantly (P<0.001) higher than in all other horizons, but showed no significant differences within each matric potential.
3.1 Effect of moisture status and horizon
We first tested the effects of changes in moisture status on the mineralisation rates of the organic C in each of the OH, E, Bh and C horizons. This was achieved by adjusting the matric potential of the samples (based on the water retention curves, Fig. S2) and measuring CO2 emissions during a subsequent incubation at 28 °C and under aerobic conditions. There were no clear trends with matric potential in any of the horizons (Fig. 2), contrary to what might be expected (Moyano et al., 2012; Sierra et al., 2017). The first hypothesis was, thus, rejected. Whilst this is surprising, the water retention curves (Fig. S2) show that sample moisture content varied little as a function of matric potential, suggesting that the saturation of the pore network changed little and that the environmental conditions of microbial decomposers in the samples were relatively unaffected. Sierra et al. (2017) showed that moisture effects on decomposition rates are strongly modulated by O2 availability. As the changes in moisture content in this experiment were small, O2 availability inside the cores may not have changed much, thus reducing any matric potential effect on decomposition.
Overall, the amounts of C mineralised from the Oh horizon were an order of magnitude higher (P<0.001) than from the other horizons (Fig. 2A), reflecting the higher organic C content of this horizon. However, the high variability in the data meant that the differences among horizons within each matric potential were not always significant. At the matric potentials of −5 and −31.6 kPa significant differences were not found with the E and Bh horizons, respectively, and neither of these horizons showed a significant difference with the OH horizon at −316 kPa.
The specific mineralisation rates (the amount of CO2 produced per unit organic C) were significantly (P<0.001) higher in the E horizon (Fig. 2B). Here also, the high variability in the data meant that specific respiration rates in the E horizon within a matric potential were not significantly different from the other horizons. The specific mineralisation rate is indicative of the mineralisability of the organic C (Fierer et al., 2003; Salomé et al., 2010). With the exception of the E horizon, the specific mineralisation rates were generally in the range of what has been found elsewhere in incubations of approximately two months (usually between 1 %–3 %, e.g. Salomé et al., 2010; Autret et al., 2020; Kan et al., 2020), although the rate of the Bh horizon was at the lower end of this range. The specific mineralisation rates in the E horizon were an order of magnitude higher however. These very high rates may be linked to the low total C content of the E horizon (2.3 mg g−1 soil), although others have not found such high specific mineralisation rates, even in deep soil where total C contents were lower than what was observed here (Fierer et al., 2003; Salomé et al., 2010). It is more likely therefore, that the high specific mineralisation rates suggest that the organic C in this horizon was highly labile and readily available. The high decomposability of the organic C in the E horizon may be due to the nature of the organic matter or due to the sandy texture of the horizon. In general, organic C is less persistent in sandy soils, possibly due to lower rates of mineral-associated organic matter formation (Haddix et al., 2020) or to more oxic conditions.
The lack of difference in the mineralisation rates across matric potentials suggests that oxygen availability did not play a major role in this particular experiment, possibly because the cores were in oxic conditions within the microcosms. The fact that the organic C in the E horizon was more mineralisable than in the Bh horizon (Fig. 2B) suggests that the specific conditions in the Bh horizon had a strong limiting effect on the mineralisation, as the C in the Bh horizon was likely transferred from the E horizon above it. The second series of incubations was carried out in order to investigate the causes of the relatively low specific mineralisation rates in the Bh horizon.
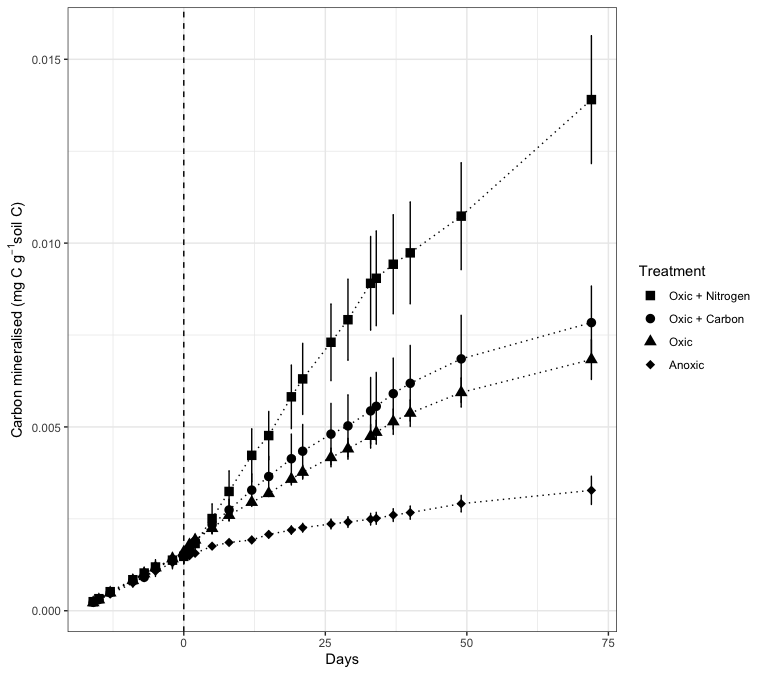
Figure 3Soil organic C mineralisation in Podzol Bh horizons prior to and after imposition of treatments (dashed line indicates day at which treatments commenced). Bars indicate standard error of the mean where error is larger than the size of the symbols. At the end of the incubation, the amount of C mineralised in the oxic + N treatment was significantly higher than the oxic and the oxic + C treatments (P<0.05) as well as the anoxic treatment (P<0.001). Both oxic treatments without N were also significantly (P<0.01) higher than the anoxic treatment.
3.2 Effects of N, C and O2 availability
The second incubation experiment was carried out to test whether the decomposition of the organic matter in the Bh horizon was N, O2 or energy limited. The undisturbed samples were incubated for two weeks under oxic conditions and at 28 °C prior to initiating the treatments, in order to ensure that there were no differences among the samples selected for each treatment (Fig. 3). There was a rapid divergence in mineralisation rates following the initiation of the treatments and, by the end of the incubation, there were significant differences (P<0.001) in the amounts of CO2 released from the samples subjected to the different treatments (Fig. 4A). The samples that received N under oxic conditions mineralised approximately twice as much organic C as the oxic control (P<0.01) and the samples that received a cocktail of simple substrates (P<0.05), and in excess of four times as much C as the samples incubated under anoxic conditions (P<0.001; Figs. 3 and S3 for O2 contents in all treatments). No significant differences were observed between the oxic treatments with or without the addition of the simple substrate cocktail, even though the substrate cocktail was rapidly mineralised after both additions (Fig. S4). Both oxic treatments without N mineralised significantly (P<0.01) more than the anoxic treatment. These data suggest that the mineralisation of the Bh horizon organic C is constrained by N and O2 availability (hypotheses 2 and 3) rather than by energetic deficiencies in the organic matter. The very large ratio of the Bh horizon organic matter (Fig. 1) tends to confirm that low N availability to soil microbial decomposers is a major limiting factor of organic matter decomposition in these soils. An analysis of the 14C-age of the organic C in other Amazonian Podzols showed that there was a negative relationship between the N content and the 14C-age of the organic matter (Montes et al., 2023), confirming that N limitation is a major factor in the dynamics of C in such soils.
Although the CO2 emissions in the anoxic treatment were significantly lower than the other treatments, they were not negligible (Fig. 3). Fairbairn et al. (2023) have suggested that CO2 emissions can remain high under anoxic conditions due to the anaerobic degradation of soil organic matter, via processes such as fermentation or anaerobic respiration. It is likely that the microbial communities in this soil were adapted to anoxic conditions, due to the water saturation conditions, and would therefore have had the capacity for anaerobic degradation. Nevertheless, the CO2 emission levels were significantly lower than in the oxic treatments, confirming the work of Sierra et al. (2017), who showed that the O2 levels impose a significant limitation on CO2 emissions.
There was a CO2 pulse after both additions of the substrate cocktail, but this was due to the mineralisation of the substrate-C that was added rather than an increase in the mineralisation of Bh horizon organic C (Figs. 3 and S4), meaning that hypothesis 4 was rejected. The lack of a priming effect may be due to the low pH of the soil (Fig. 1). The priming effect is more common in soils with pHs between 5.5 and 7.5 but tends to be lower at the pH values found here (Wang and Kuzyakov, 2024). Furthermore, it has been shown that soils that are characterised by high levels of mineral associated organic C, as is the case in the Bh horizons of Podzols (Schmidt et al., 2000; Doupoux et al., 2017), also tend to be less prone to the priming effect (Chen et al., 2019). It should also be noted that a month after the first addition of the substrate cocktail, only 22 % of the added C was mineralised, and only 15 % was mineralised slightly more than a month after the second addition (Fig. S4). These mineralisation rates are lower than what is usually found. The mineralisation of glucose often exceeds 60 % after a month's incubation (e.g. Hamer and Marschner, 2005), while that of pyruvate and vanilin can exceed 30 % (Chenu et al., 2025) and 20 % (Juarez et al., 2013), respectively. These low mineralisation rates may also have been due to an N limitation, but this would have to be confirmed experimentally. Nitrogen limitation can arise due to microbial cells being unable to produce proteins, such as enzymes or membrane transport proteins, necessary for activity, as proteins are N rich molecules (Nunan et al., 2020).
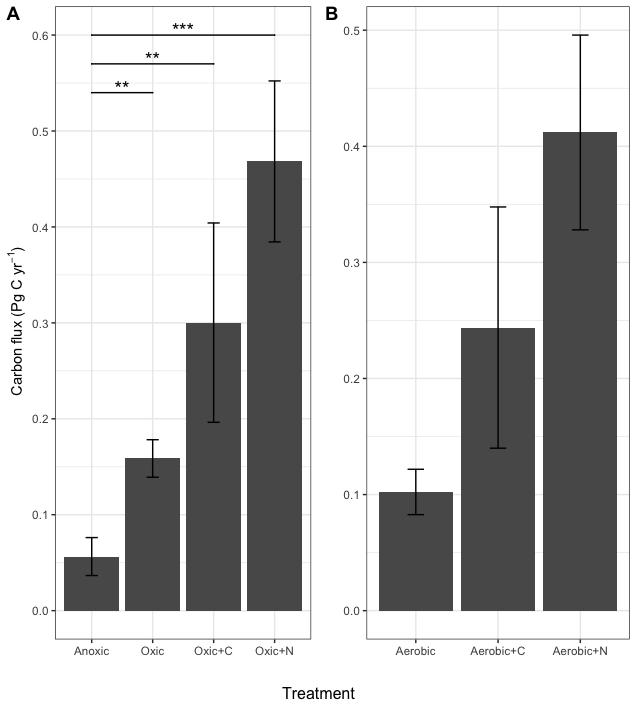
Figure 4Estimated annual carbon flux from Bh horizons to the atmosphere (A) and estimated increase in C flux to the atmosphere if the present anoxic conditions were to change (B). The bars indicate the standard error of the mean. The oxic + N and the oxic + C treatments resulted in significantly (** <0.01) more C flux than the anoxic treatment.
In view of the very large quantities of organic C that are stored in the Bh horizon of the Amazonian Podzols (Montes et al., 2011), we sought to estimate the annual CO2 flux from these horizons to the atmosphere under the different conditions tested here. We first fitted a first order decay model with a single pool to the respiration data (Fig. S5 – best fit based on the Akaike Information Criterion) and extrapolated the mineralisation curves to a year. Montes et al. (2011) estimated that 78.8 % of the 13 Pg C in the Podzol profiles is found in the Bh horizon (10.45 Pg C), which we used to estimate the potential total C fluxes from the Bh Horizon of the Amazonian Podzols (Fig. 4). The increase in CO2 flux in the oxic treatment with N translates to an extra 0.41 Pg C yr−1 being released to the atmosphere compared to the anoxic treatment (P<0.001). The other two oxic treatments also resulted in significant increases in the amount of C released (P<0.01).
Global soil respiration estimates are subject to large uncertainties, due to the complex set of biogeochemical and biophysical processes that are involved. These uncertainties are one of the major causes of uncertainty in terrestrial ecosystem models (He et al., 2022). Nevertheless, a recent study has estimated global soil heterotrophic respiration to be 48.8±0.9 Pg C yr−1 (He et al., 2022). The potential increase in CO2 flux from Amazonian Podzols could therefore be equivalent to 0.8 % of global soil heterotrophic respiration.
There are a number of uncertainties associated with the estimates put forward in this study. The first is that it is a laboratory study and, even though the samples were undisturbed, the experiment and the treatments are somewhat artificial. For example, the Bh horizons can often be found at depths greater than 1 m (Doupoux et al., 2017) and the degree to which O2 or N would reach it is uncertain. Ideally, an experiment testing similar treatments should be carried out in situ in order to determine the magnitude of the vulnerability of the organic C to N and O2 availability, though this would be extremely difficult.
The second uncertainty is the use of a first order decay model with only one pool to extrapolate to yearly CO2 carbon fluxes. The use of single pool models has been criticised in the past because it can mask the presence of smaller C pools with faster turnover times (Davidson et al., 2000). This criticism is less relevant here because the pool is small and even if it did respond differently to the treatments, it would not change the overall trend of the results. Furthermore, one might expect this smaller pool to respond more rapidly to N availability and, if anything, increase the observed N effect. Nitrogen additions are often associated with an increase in enzymes that catalyse carbohydrate hydrolysis but a decrease in oxidative enzymes that catalyse the breakdown of polyphenols (Moorhead and Sinsabaugh, 2006), thus increasing the decomposition rates of the fast-cycling C pool and slowing down the decomposition of the slow-cycling C pool. Although, the decomposition of fast-cycling C pools is likely less sensitive to variations in O2 levels than that of slow-cycling C pools (Lin et al., 2021), the smaller size of the pool is unlikely to change the overall conclusions, as suggested above.
The third uncertainty lies in the duration of the incubations (72 d), which may not have been long enough to detect the response of the slower-cycling C pools in these soils. However, the parameters obtained from the first order decay model suggest that the treatments increased the size of the mineralisable pool rather than the rate at which the pool was mineralised (Table S1 in the Supplement). This suggests that the treatments increased the amount of C that was readily mineralisable in the samples and, therefore, not detecting the slow C pool's response may not be problematic as this change was detected. Furthermore, Montes et al. (2023) measured CO2 emissions from Bh horizon samples of Amazonian Podzols and found that the fast pool always accounted for <0.5 % soil C, far less than in any of the treatments here, other than the anoxic treatment.
A fourth uncertainty associated with the study is that the amount of N arriving in the Bh horizons annually may be lower than what was added here, despite the fact that future N deposition projections indicate significant increases in reactive N deposition (Galloway et al., 2004) and forest dieback might also result in higher soil N concentrations (Xiong et al., 2011).
The CO2 flux from the Amazonian Podzols may be further enhanced by increases in average temperature that will occur with climate change, due to the very high ratio of the Bh horizon: CO2 fluxes from soils with high ratios show a positive response to temperature (Karhu et al., 2014). Overall, the significant increases in CO2 emissions under oxic and N amended treatments suggest that the organic C contained in equatorial Podzols is vulnerable to N and O2 availability and that significant amounts of CO2 could be released to the atmosphere, thus exacerbating atmospheric CO2 levels.
The data that support the findings of this study are openly available in Zenodo at https://doi.org/10.5281/zenodo.15824641 (Nunan, 2025).
The supplement related to this article is available online at https://doi.org/10.5194/bg-23-1279-2026-supplement.
NN: Conceptualization, Investigation, Methodology, Data curation, Formal analysis, Writing – original draft, Writing – review & editing, Funding acquisition. CC: Conceptualization, Investigation, Writing – review & editing. VP: Investigation, Data curation, Methodology, Writing – review & editing. AS: Investigation, Data curation, Methodology, Writing – review & editing. KP: Investigation, Data curation, Methodology, Writing – review & editing. CRM: Investigation, Writing – review & editing. PM: Investigation, Writing – review & editing. AJM: Investigation, Writing – review & editing. YL: Conceptualization, Investigation, Writing – review & editing, Funding acquisition.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
The authors would like to acknowledge the contribution of two anonymous referees whose constructive comments helped improve the paper.
This research has been supported by the Agence Nationale de la Recherche (grant no. ANR-12-IS06-0002).
This paper was edited by Sara Vicca and reviewed by two anonymous referees.
Autret, B., Guillier, H., Pouteau, V., Mary, B., and Chenu, C.: Similar specific mineralization rates of organic carbon and nitrogen in incubated soils under contrasted arable cropping systems, Soil Tillage Res., 204, 104712, https://doi.org/10.1016/j.still.2020.104712, 2020.
Avila-Diaz, A., Benezoli, V., Justino, F., Torres, R., and Wilson, A.: Assessing current and future trends of climate extremes across Brazil based on reanalyses and earth system model projections, Clim. Dynam., 55, 1403–1426, https://doi.org/10.1007/s00382-020-05333-z, 2020.
Bernoux, M., Da Conceição Santana Carvalho, M., Volkoff, B., and Cerri, C. C.: Brazil's Soil Carbon Stocks, Soil Sci. Soc. Am. J., 66, 888–896, https://doi.org/10.2136/sssaj2002.8880, 2002.
Boulton, C. A., Lenton, T. M., and Boers, N.: Pronounced loss of Amazon rainforest resilience since the early 2000s, Nat. Clim. Change, 12, 271–278, https://doi.org/10.1038/s41558-022-01287-8, 2022.
Chen, L., Liu, L., Qin, S., Yang, G., Fang, K., Zhu, B., Kuzyakov, Y., Chen, P., Xu, Y., and Yang, Y.: Regulation of priming effect by soil organic matter stability over a broad geographic scale, Nat. Commun., 10, 5112, https://doi.org/10.1038/s41467-019-13119-z, 2019.
Chenu, C., Pouteau, V., and Nunan, N.: Pore scale microbial biogeography across different soil types, Soil Biol. Biochem., 109896, https://doi.org/10.1016/j.soilbio.2025.109896, 2025.
Davidson, E. A., Trumbore, S. E., and Amundson, R.: Biogeochemistry – Soil warming and organic carbon content, Nature, 408, 789–790, 2000.
Davidson, E. A., Savage, K. E., and Finzi, A. C.: A big-microsite framework for soil carbon modeling, Glob. Change Biol., 20, 3610–3620, 2014.
Doupoux, C., Merdy, P., Montes, C. R., Nunan, N., Melfi, A. J., Pereira, O. J. R., and Lucas, Y.: Modelling the genesis of equatorial podzols: age and implications for carbon fluxes, Biogeosciences, 14, 2429–2440, https://doi.org/10.5194/bg-14-2429-2017, 2017.
Esquivel‐Muelbert, A., Baker, T. R., Dexter, K. G., Lewis, S. L., Brienen, R. J. W., Feldpausch, T. R., Lloyd, J., Monteagudo‐Mendoza, A., Arroyo, L., Álvarez-Dávila, E., Higuchi, N., Marimon, B. S., Marimon-Junior, B. H., Silveira, M., Vilanova, E., Gloor, E., Malhi, Y., Chave, J., Barlow, J., Bonal, D., Davila Cardozo, N., Erwin, T., Fauset, S., Hérault, B., Laurance, S., Poorter, L., Qie, L., Stahl, C., Sullivan, M. J. P., Ter Steege, H., Vos, V. A., Zuidema, P. A., Almeida, E., Almeida De Oliveira, E., Andrade, A., Vieira, S. A., Aragão, L., Araujo‐Murakami, A., Arets, E., Aymard, C. G. A., Baraloto, C., Camargo, P. B., Barroso, J. G., Bongers, F., Boot, R., Camargo, J. L., Castro, W., Chama Moscoso, V., Comiskey, J., Cornejo Valverde, F., Lola Da Costa, A. C., Del Aguila Pasquel, J., Di Fiore, A., Fernanda Duque, L., Elias, F., Engel, J., Flores Llampazo, G., Galbraith, D., Herrera Fernández, R., Honorio Coronado, E., Hubau, W., Jimenez‐Rojas, E., Lima, A. J. N., Umetsu, R. K., Laurance, W., Lopez‐Gonzalez, G., Lovejoy, T., Aurelio Melo Cruz, O., Morandi, P. S., Neill, D., Núñez Vargas, P., Pallqui Camacho, N. C., Parada Gutierrez, A., Pardo, G., Peacock, J., Peña‐Claros, M., Peñuela‐Mora, M. C., Petronelli, P., Pickavance, G. C., Pitman, N., Prieto, A., Quesada, C., Ramírez‐Angulo, H., Réjou‐Méchain, M., Restrepo Correa, Z., Roopsind, A., Rudas, A., Salomão, R., Silva, N., Silva Espejo, J., Singh, J., Stropp, J., Terborgh, J., Thomas, R., Toledo, M., Torres‐Lezama, A., Valenzuela Gamarra, L., Van De Meer, P. J., Van Der Heijden, G., Van Der Hout, P., Vasquez Martinez, R., Vela, C., Vieira, I. C. G., and Phillips, O. L.: Compositional response of Amazon forests to climate change, Glob. Change Biol., 25, 39–56, https://doi.org/10.1111/gcb.14413, 2019.
Fairbairn, L., Rezanezhad, F., Gharasoo, M., Parsons, C. T., Macrae, M. L., Slowinski, S., and Van Cappellen, P.: Relationship between soil CO2 fluxes and soil moisture: Anaerobic sources explain fluxes at high water content, Geoderma, 434, 116493, https://doi.org/10.1016/j.geoderma.2023.116493, 2023.
Fierer, N., Allen, A. S., Schimel, J. P., and Holden, P. A.: Controls on microbial CO2 production: a comparison of surface and subsurface soil horizons, Glob. Change Biol., 9, 1322–1332, 2003.
Flores, B. M., Montoya, E., Sakschewski, B., Nascimento, N., Staal, A., Betts, R. A., Levis, C., Lapola, D. M., Esquível-Muelbert, A., Jakovac, C., Nobre, C. A., Oliveira, R. S., Borma, L. S., Nian, D., Boers, N., Hecht, S. B., Ter Steege, H., Arieira, J., Lucas, I. L., Berenguer, E., Marengo, J. A., Gatti, L. V., Mattos, C. R. C., and Hirota, M.: Critical transitions in the Amazon forest system, Nature, 626, 555–564, https://doi.org/10.1038/s41586-023-06970-0, 2024.
Fontaine, S., Barot, S., Barre, P., Bdioui, N., Mary, B., and Rumpel, C.: Stability of organic carbon in deep soil layers controlled by fresh carbon supply, Nature, 450, 277–280, https://doi.org/10.1038/nature06275, 2007.
Galloway, J. N., Dentener, F. J., Capone, D. G., Boyer, E. W., Howarth, R. W., Seitzinger, S. P., Asner, G. P., Cleveland, C. C., Green, P. A., Holland, E. A., Karl, D. M., Michaels, A. F., Porter, J. H., Townsend, A. R., and Vöosmarty, C. J.: Nitrogen Cycles: Past, Present, and Future, Biogeochemistry, 70, 153–226, https://doi.org/10.1007/s10533-004-0370-0, 2004.
Galloway, J. N., Townsend, A. R., Erisman, J. W., Bekunda, M., Cai, Z., Freney, J. R., Martinelli, L. A., Seitzinger, S. P., and Sutton, M. A.: Transformation of the Nitrogen Cycle: Recent Trends, Questions, and Potential Solutions, Science, 320, 889–892, https://doi.org/10.1126/science.1136674, 2008.
Haddix, M. L., Gregorich, E. G., Helgason, B. L., Janzen, H., Ellert, B. H., and Francesca Cotrufo, M.: Climate, carbon content, and soil texture control the independent formation and persistence of particulate and mineral-associated organic matter in soil, Geoderma, 363, 114160, https://doi.org/10.1016/j.geoderma.2019.114160, 2020.
Hamer, U. and Marschner, B.: Priming effects in soils after combined and repeated substrate additions, Geoderma, 128, 38–51. https://doi.org/10.1016/j.geoderma.2004.12.014, 2005.
He, Y., Ding, J., Dorji, T., Wang, T., Li, J., and Smith, P.: Observation-based global soil heterotrophic respiration indicates underestimated turnover and sequestration of soil carbon by terrestrial ecosystem models, Glob. Change Biol., 28, 5547–5559, https://doi.org/10.1111/gcb.16286, 2022.
Juarez, S., Nunan, N., Duday, A.-C., Pouteau, V., Schmidt, S., Hapca, S., Falconer, R., Otten, W., and Chenu, C.: Effects of different soil structures on the decomposition of native and added organic carbon, Eur. J. Soil Biol., 58, 81–90, https://doi.org/10.1016/j.ejsobi.2013.06.005, 2013.
Kaiser, K., Guggenberger, G., Haumaier, L., and Zech, W.: Seasonal variations in the chemical composition of dissolved organic matter in organic forest floor layer leachates of old-growth Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) stands in northeastern Bavaria, Germany, Biogeochemistry, 55, 103–143. https://doi.org/10.1023/A:1010694032121, 2001.
Kan, Z.-R., He, C., Liu, Q.-Y., Liu, B.-Y., Virk, A. L., Qi, J.-Y., Zhao, X., and Zhang, H.-L.: Carbon mineralization and its temperature sensitivity under no-till and straw returning in a wheat-maize cropping system, Geoderma, 377, 114610, https://doi.org/10.1016/j.geoderma.2020.114610, 2020.
Karhu, K., Auffret, M. D., Dungait, J. A. J., Hopkins, D. W., Prosser, J. I., Singh, B. K., Subke, J.-A., Wookey, P. A., Ågren, G. I., Sebastià, M.-T., Gouriveau, F., Bergkvist, G., Meir, P., Nottingham, A. T., Salinas, N., and Hartley, I. P.: Temperature sensitivity of soil respiration rates enhanced by microbial community response, Nature, 513, 81–84, https://doi.org/10.1038/nature13604, 2014.
Lin, Y., Campbell, A. N., Bhattacharyya, A., DiDonato, N., Thompson, A. M., Tfaily, M. M., Nico, P. S., Silver, W. L., and Pett-Ridge, J.: Differential effects of redox conditions on the decomposition of litter and soil organic matter, Biogeochemistry, 154, 1–15, https://doi.org/10.1007/s10533-021-00790-y, 2021.
Linn, D. M. and Doran, J. W.: Effect of Water-Filled Pore Space on Carbon Dioxide and Nitrous Oxide Production in Tilled and Nontilled Soils, Soil Sci. Soc. Am. J., 48, 1267–1272, https://doi.org/10.2136/sssaj1984.03615995004800060013x, 1984.
Malhi, Y., Wood, D., Baker, T. R., Wright, J., Phillips, O. L., Cochrane, T., Meir, P., Chave, J., Almeida, S., Arroyo, L., Higuchi, N., Killeen, T. J., Laurance, S. G., Laurance, W. F., Lewis, S. L., Monteagudo, A., Neill, D. A., Vargas, P. N., Pitman, N. C. A., Quesada, C. A., Salomao, R., Silva, J. N. M., Lezama, A. T., Terborgh, J., Martinez, R. V., and Vinceti, B.: The regional variation of aboveground live biomass in old-growth Amazonian forests, Glob. Change Biol., 12, 1107–1138, https://doi.org/10.1111/j.1365-2486.2006.01120.x, 2006.
Manzoni, S., Piñeiro, G., Jackson, R. B., Jobbágy, E. G., Kim, J. H., and Porporato, A.: Analytical models of soil and litter decomposition: Solutions for mass loss and time-dependent decay rates, Soil Biol. Biochem., 50, 66–76, https://doi.org/10.1016/j.soilbio.2012.02.029, 2012.
Marengo, J. A., Cunha, A. P., Espinoza, J. C., Fu, R., Schöngart, J., Jimenez, J. C., Costa M. C., Ribeiro, J. M., Wongchuig, S., and Zhao, S.: The Drought of Amazonia in 2023–2024, Am. J. Climate Change, 13, 567–597, 2024.
Montes, C. R., Lucas, Y., Pereira, O. J. R., Achard, R., Grimaldi, M., and Melfi, A. J.: Deep plant-derived carbon storage in Amazonian podzols, Biogeosciences, 8, 113–120, https://doi.org/10.5194/bg-8-113-2011, 2011.
Montes, C. R., Merdy, P., Da Silva, W. T. L., Ishida, D., Melfi, A. J., Santin, R. C., and Lucas, Y.: Mineralization of soil organic matter from equatorial giant podzols submitted to drier pedoclimate: A drainage topochronosequence study, CATENA, 222, 106837, https://doi.org/10.1016/j.catena.2022.106837, 2023.
Moorhead, D. L. and Sinsabaugh, R. L.: A theoretical model of litter decay and microbial interaction, Ecol. Monogr., 76, 151–174, https://doi.org/10.1890/0012-9615(2006)076[0151:ATMOLD]2.0.CO;2, 2006.
Moyano, F. E., Vasilyeva, N., Bouckaert, L., Cook, F., Craine, J., Curiel Yuste, J., Don, A., Epron, D., Formanek, P., Franzluebbers, A., Ilstedt, U., Kätterer, T., Orchard, V., Reichstein, M., Rey, A., Ruamps, L., Subke, J.-A., Thomsen, I. K., and Chenu, C.: The moisture response of soil heterotrophic respiration: interaction with soil properties, Biogeosciences, 9, 1173–1182, https://doi.org/10.5194/bg-9-1173-2012, 2012.
Nunan, N.: Amazonian Podzols – a carbon time bomb? – dataset, Zenodo [data set], https://doi.org/10.5281/zenodo.15824641, 2025.
Nunan, N., Schmidt, H., and Raynaud, X.: The ecology of heterogeneity: soil bacterial communities and C dynamics, Philos. Trans. R. Soc. B Biol. Sci., 375, 20190249, https://doi.org/10.1098/rstb.2019.0249, 2020.
Quesada, C. A., Lloyd, J., Anderson, L. O., Fyllas, N. M., Schwarz, M., and Czimczik, C. I.: Soils of Amazonia with particular reference to the RAINFOR sites, Biogeosciences, 8, 1415–1440, https://doi.org/10.5194/bg-8-1415-2011, 2011.
R Core Team: R: A language and environment for statistical computing, R Foundation for Statistical Computing, Vienna, Austria, https://www.R-project.org/ (last access: 6 February 2026), 2022.
Rovira, A. and Greacen, E.: The effect of aggregate disruption on the activity of microorganisms in the soil, Aust. J. Agric. Res., 8, 659, https://doi.org/10.1071/AR9570659, 1957.
Ruamps, L. S., Nunan, N., and Chenu, C.: Microbial biogeography at the soil pore scale, Soil Biol. Biochem., 43, 280–286, https://doi.org/10.1016/j.soilbio.2010.10.010, 2011.
Salomé, C., Nunan, N., Pouteau, V., Lerch, T. Z., and Chenu, C.: Carbon dynamics in topsoil and in subsoil may be controlled by different regulatory mechanisms, Glob. Change Biol., 16, 416–426, 2010.
Schmidt, M. W. I., Knicker, H., and Kögel-Knabner, I.: Organic matter accumulating in Aeh and Bh horizons of a Podzol – chemical characterization in primary organo-mineral associations, Org. Geochem., 31, 727–734, https://doi.org/10.1016/S0146-6380(00)00045-0, 2000.
Sierra, C. A., Jiménez, E. M., Reu, B., Peñuela, M. C., Thuille, A., and Quesada, C. A.: Low vertical transfer rates of carbon inferred from radiocarbon analysis in an Amazon Podzol, Biogeosciences, 10, 3455–3464, https://doi.org/10.5194/bg-10-3455-2013, 2013.
Sierra, C. A., Malghani, S., and Loescher, H. W.: Interactions among temperature, moisture, and oxygen concentrations in controlling decomposition rates in a boreal forest soil, Biogeosciences, 14, 703–710, https://doi.org/10.5194/bg-14-703-2017, 2017.
Vaughn, L. J. S. and Torn, M. S.: 14C evidence that millennial and fast-cycling soil carbon are equally sensitive to warming, Nat. Clim. Change, 9, 467–471, https://doi.org/10.1038/s41558-019-0468-y, 2019.
Wang, C. and Kuzyakov, Y.: Soil organic matter priming: The pH effects, Glob. Change Biol., 30, e17349, https://doi.org/10.1111/gcb.17349, 2024.
Xiong, Y., D'Atri, J. J., Fu, S., Xia, H., and Seastedt, T. R.: Rapid soil organic matter loss from forest dieback in a subalpine coniferous ecosystem, Soil Biol. Biochem., 43, 2450–2456, https://doi.org/10.1016/j.soilbio.2011.08.013, 2011.