the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 09 Jul 2026
| 09 Jul 2026
Biosignatures of microbial mats in Pleistocene coral reef cores from IODP Expedition 389 (Hawaiian Drowned Reefs)
Hildegard Westphal
Elisa Garuglieri
Gregory E. Webb
Luke Nothdurft
Anna Merkel
Pankaj Khanna
Poornima Karki
Theresa Nohl
Eberhard Gischler
Jody M. Webster
We systematically document surfaces and biosignatures of Pleistocene reefal microbialites (Marine Isotope Stages 7–6) recovered during IODP Expedition 389 (Hawaiian Drowned Reefs). Microbialites are abundant within Pleistocene coral reef successions and offer valuable archives of environmental change during Quaternary climate variability. However, relatively little is known about biofilm-forming microbial consortia, because biosignature preservation is usually very poor. The microbial crusts studied here form encrustations as much as 20 cm thick, ranging from laminated to thrombolitic, within the coral reef framework. Scanning electron microscopy (SEM) of samples from the windward (humid) Hilo and Kohala and the leeward (arid) Kawaihae sides of the Island of Hawai`i reveals exceptionally well-preserved microbial fabrics that developed during Marine Isotope Stages (MIS) 7–6. Humid side samples exhibit abundant putative calcified exopolymeric substance (EPS) matrices, mineralised filaments, and near spherical, multilobate aggregates that resemble protodolomite spherules formed by modern cyanobacteria or extant coccoid cyanobacteria (e.g., Gloeocapsa-type). In either case, the surfaces appear to have been formed with significant aid of cyanobacteria, suggesting formation in a euphotic setting. The microbialites from the arid side display peloidal microfabrics with fewer preserved physical biosignatures, typical of cryptic reefal microbialites, but the surfaces suggest confinement by an organic biofilm. The occurrence of pyrite framboids and huntite-like crystals in the wet-side samples suggests local redox gradients consistent with both sulphate reducing bacteria and cyanobacteria mediating carbonate precipitation. These findings provide the first direct evidence for euphotic microbial mat communities contributing to microbialite formation in Pleistocene drowned Hawai'ian reefs, and, to our knowledge, in Indopacific reefs and beyond. The outstanding petrological preservation of calcified EPS and microbial morphotypes highlights the potential of these Pleistocene reefal microbialites as sensitive archives of palaeoenvironmental conditions and microbial diversity under glacio-eustatic forcing and associated environmental changes.
- Article
(16981 KB) - Full-text XML
- BibTeX
- EndNote




Microbialites are organosedimentary deposits formed by benthic microbial communities through processes such as trapping, binding, and mineral precipitation. They represent some of the most ancient ecosystems in Earth's history and the first to form hard frameworks (Burne and Moore, 1987; Riding, 2006). Their fossil record spans at least 3.5 billion years, providing insights into early life, biogeochemical cycling, and environmental conditions across geologic time (Grotzinger and Knoll, 1999; Riding, 2000; Riding, 2006; Allwood et al., 2006; Noffke and Awramik, 2026). While marine stromatolites were globally dominant during the Precambrian and re-emerged prominently following mass extinction events (e.g., the end-Permian; Schubert and Bottjer, 1992; Baud et al., 1997; Webb, 2005; Pei et al., 2022), other microbialite types were important constituents of skeletal reefs throughout the Phanerozoic (e.g., Pratt, 1982; Webb, 1996). Microbialite occurrence in the Pleistocene in coral reef-associated, high-diversity, normal marine environments has drawn increasing attention for understanding carbonate platform dynamics and geomicrobiological processes under Quaternary climate variability (Reid et al., 2000; Dupraz and Visscher, 2005; Riding et al., 2014; Salas-Saavedra et al., 2018).
Pleistocene reefal microbialites provide archives of environmental change associated with glacio-eustatic and climatic fluctuations, since the microbial communities involved in their formation are highly sensitive to environmental parameters that affect metabolic processes and marine pH (cf., Riding, 2000b, 2014; Webb and Kamber, 2010). Specifically, Pleistocene reefal microbialites have been documented across a range of tropical and subtropical carbonate provinces, including the western Atlantic, Indo-Pacific, Arabian Gulf, and the Red Sea (Camoin et al., 1999; Reitner et al., 2000; Westphal et al., 2010; Heindel et al., 2012; Braga et al., 2019, among many others). Most, but not all, occurrences are linked to deglacial phases, transgressive reef systems, and possibly lagoonal back-reef environments (White et al., 1984; Camoin et al., 1999; Szilagyi et al., 2020). Holocene reefal microbialite crusts from Tahiti were interpreted early on as bio-accretionary features (Camoin and Montaggioni, 1994). In general, such microbialite formation requires: (1) a microbial consortium, (2) an organic carbon source (a local primary producer acting as energy source for microbial metabolism) that may, or may not, be a major part of the microbial community, (3) an appropriate nucleation template or scaffold to localize and support mineral precipitation (typically microbial exopolysaccharides – EPS), and (4) a local driver of increased pH and/or alkalinity, generally associated with microbial metabolism (Castanier et al., 1999; Visscher and Stolz, 2005; Dupraz et al., 2009; Heindel et al., 2010; Webb and Kamber, 2010; Zhu and Dittrich, 2016; Reid et al., 2024). A general perception exists that the best-known microbialites, benthic stromatolites, are made by cyanobacteria. Although photosynthetic removal of CO2 by cyanobacteria and filamentous green algae can increase pH within benthic biofilms, carbonate precipitation is also driven by processes deeper within the mat, where microbial heterotrophy processes organic matter from the overlying primary producers. Here, heterotrophs can modify potential nucleation scaffolds (i.e., EPS) and create anoxic microenvironments where sulphate reduction and other heterotrophic metabolic processes can deeply influence pH, increasing local alkalinity (Castanier et al., 1999; Dupraz et al., 2009; Heindel et al., 2010). The mechanisms involved in microbial carbonate formation within variably confined microenvironments (Dupraz et al., 2009; Webb and Kamber, 2010) are complex, ranging from rather passive calcification of fresh or degraded EPS to mineral precipitation induced by microbial activities (organomineralisation and biologically induced or influenced mineralisation, respectively; e.g., Reitner et al., 1995; Garuglieri et al., 2024). In all cases, the higher the ambient CaCO3 saturation state, the easier it is for microbial carbonate to accumulate (Riding and Liang, 2005), and microbialite mineral precipitation itself may occur only in highly chemically evolved microenvironments very different from the ambient one that surrounds them (e.g., anoxic versus fully oxygenated; Nothdurft et al., 2007). Hence, microbialites represent a combination of biotic and abiotic factors, thus providing a wealth of (paleo)environmentally significant morphological, textural, mineralogical and biogeochemical information (e.g., Riding, 2000b; Riding and Awramik, 2000; Dupraz and Strasser, 2002; Westphal et al., 2010; Webb and Kamber, 2010).
Reefal microbialites have a variety of morphologies and textures including laminated, thrombolitic, or dendritic structures formed mainly in aphotic (Reitner et al., 1995; Webb and Jell, 1997) to euphotic (shaded?) shallow cryptic reef settings (Westphal et al., 2010), while benthic examples occur in deeper fore-reef slope settings (Heindel et al., 2010; Riding, 2011; Braga et al., 2019; Szilagyi et al., 2020). The study of Pleistocene microbialites from Tahiti (IODP Expedition 310) and the Great Barrier Reef (IODP Expedition 325) pointed to sulphate-reducing bacteria as the main drivers of carbonate precipitation in reefal microbialites (Heindel et al., 2009a; Braga et al., 2019). Nevertheless, the roles of differing trophic levels and metabolic pathways for microbialite formation in general have not been unravelled, and many potential variables exist (e.g., benthic versus tight cryptic, and euphotic versus aphotic settings). Of particular interest for the IODP Expedition 389 successions from leeward and windward settings is that microbialites are affected by differing or fluctuating terrigenous sediment and nutrient flux caused by differences in geography and changing climatic (for example on islands with an exposed and a more protected side, such as Lizard Island, GBR: Reitner, 1993; Tahiti: Camoin et al., 1999) and oceanographic conditions (more intense circulation during deglaciation causing upwelling; as observed for example in the northern Great Barrier Reef: Marshall and Davies, 1988). Additionally, changing atmospheric pCO2 levels greatly affect marine carbonate saturation (Riding et al., 2014; Szilagyi et al., 2020).
A major hindrance to understanding microbialites is that different indicators highlight distinct aspects of microbialite formation. While biomarker analyses in previous studies point to heterotrophic bacteria (i.e., light-independent organisms) steering calcification, microbioerosion patterns in the surfaces of microbialites (Heindel et al., 2009b) and the taxa of interlayered red algae (Westphal et al., 2010) are consistent with euphotic conditions where the primary producers could have been cyanobacteria. Other reef microbialites formed entirely in aphotic crypts (Webb and Jell, 1997; Gischler et al., 2017) where primary producers may have been cryptic sponges and ascidians (Reitner et al., 1995; Braga et al., 2019; Maak et al., 2024). This emphasizes the broad range of environmental requirements and mechanisms for growth of different reefal microbialites, but also highlights an enigmatic gap in the record of possible photoautotrophic bacteria in the microbialites that formed in euphotic conditions; their presence currently can only be hypothesized. The questions remain as to what are the primary producers in reef microbialite consortia, and what types of communities or bulk metabolisms drive most mineral precipitation? Until we understand those issues, it will be difficult to retrieve all of the potential environmental data that microbialites record.
In most microbialites throughout Earth history, the preservation of identifiable microbial remains is generally poor except for calcimicrobes that were common only in Palaeozoic reefs (Webb, 1996). In that case, carbonate precipitated on or replaced EPS, thereby preserving microfossils (e.g., the preserved filament sheaths of Girvanella; Zhang et al., 2024). However, precipitation may occur only on degraded organic matter and EPS deeper within the mat, thus preserving organic matter that lacks easily identifiable morphology (Reitner et al., 1995). Additionally, continued alkalinity generation beneath the confining active mat may then drive additional precipitation, coarsening original crystals over time and further overprinting any original organic morphology (e.g., Webb et al., 1998). Thus, preservation of microfossils in microbialites is the exception to the rule (Webb and Kamber, 2010). However, where abundant precipitation occurs near the accretionary surface of the mat, recognizable and informative fossilized microbes may be more abundant.
Prior to the drilling of IODP Expedition 389, the reefal microbialites of Hawai'i attracted little attention (Webster et al., 2009). Limited to ROV and dredge samples, two microbial fabrics were observed in samples from the top of Hawai'ian drowned reefs: peloidal sediment-filled cavities and laminated to clotted micrite crusts, as much as 5 cm thick, in some cases forming digitate stromatolite overgrowths. Both fabrics were interpreted to postdate the deep-water coralline-algal facies (60–120 m), indicating deposition at greater depths as part of the drowning sequence (Webster et al., 2009). High recovery and extremely good core quality of IODP Expedition 389 now allow for a detailed study of the reef components and succession (Webster et al., 2025). The objective of the present study is to explore MIS 7–6 aged (approximately 133–225 kyr, Webster et al., 2025) reefal microbialites, for gaining insights into their architecture and to characterise the microbial contribution to microbialite formation in coral reefs.
IODP Expedition 389 aimed to explore the drowned coral reefs on the rapidly subsiding margins of Hawai'i as archives of sea level and climate change (Webster et al., 2023). As a direct result of Hawai'i's rapid and nearly constant subsidence caused by flexure of the oceanic lithosphere (2.5–2.6 m kyr−1; Watts, 1978; Webster et al., 2009; Puga-Bernabéu et al., 2016), a 100–200 m thick expanded sequence of shallow coral reef-dominated facies is preserved within the terraces off the Hawai'ian coast. Thus, Hawai'ian reefs cover the time intervals leading into, during, and out of the majority of the last five to six glacial cycles (Webster et al., 2023, 2024, 2025). One objective of this expedition was to gain a better understanding of ancient microbial communities within the reefs and their role in reef building. Based on compiled bathymetric data sets, 13 major terraces were identified around the Island of Hawai'i (designated H0–H12 from shallowest to deepest, where “H” is for Hawai'i) (Webster et al., 2009; Puga-Bernabéu et al., 2016; Sanborn et al., 2017; Taylor, 2019). Drilling operations took place from September to November 2023 with a seafloor drill (PROD5), achieving very good recovery (Webster et al., 2024). Besides well-preserved corals, the cores contain a high percentage of microbialites in the coral reef succession, forming crusts as much as 20 cm in thickness and making up a substantial portion of the recovered lithologies (Webster et al., 2025). Here we study the H2 terrace as recovered at Sites M0097, M0102, and M0104 (Figs. 1 and A1). The H2 terrace, with the top dated at 130 kyr, developed primarily during MIS 7–6, before drowning during the transition to MIS 5e (Fig. 2).
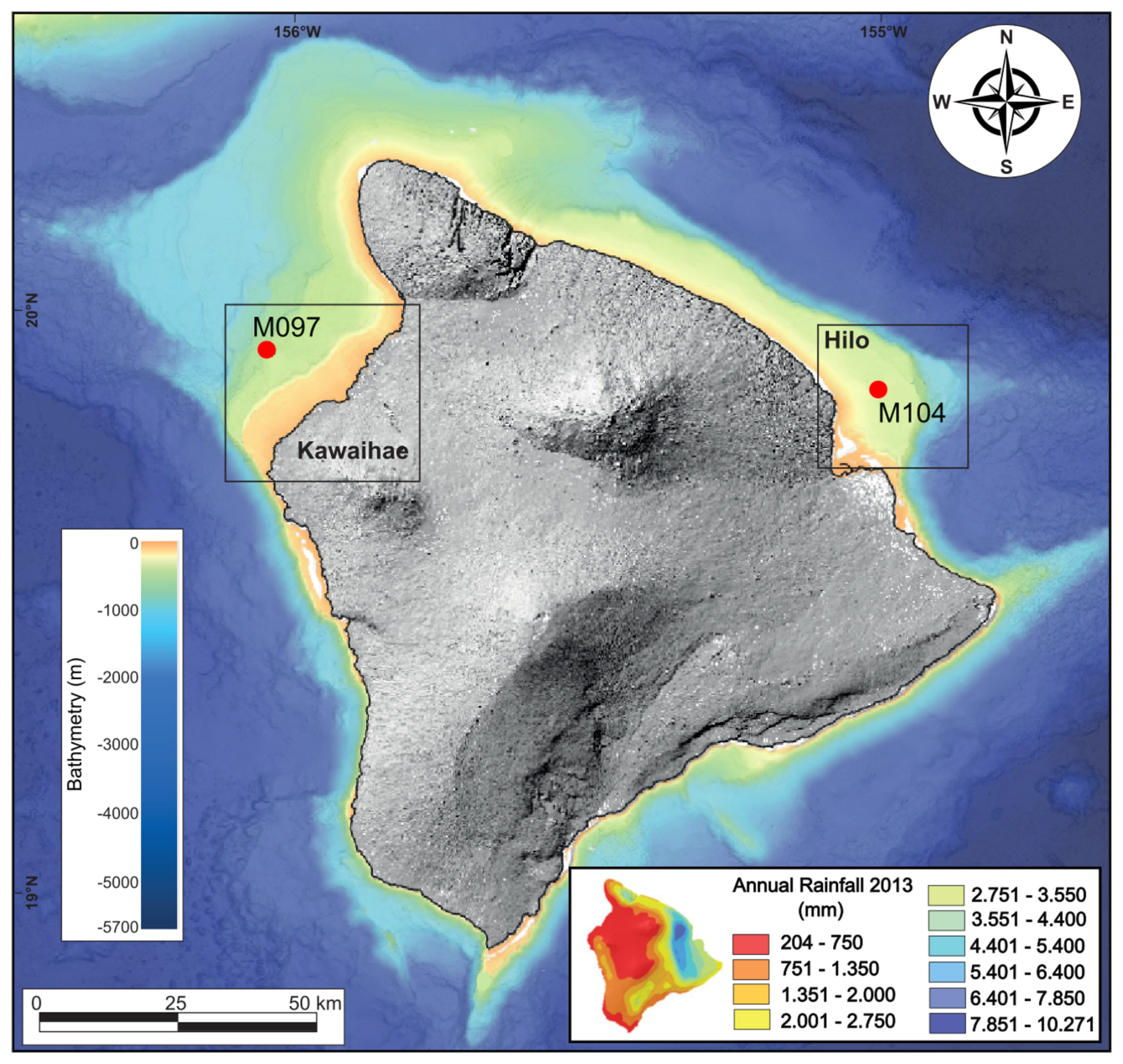
Figure 1IODP Expedition 389 drill sites M0097, M0102, M0104 studied here; bathymetric map from SOEST (2016; see Webster et al., 2024). Map of modern annual rainfall in the inset (Sanfilippo et al., 2024).
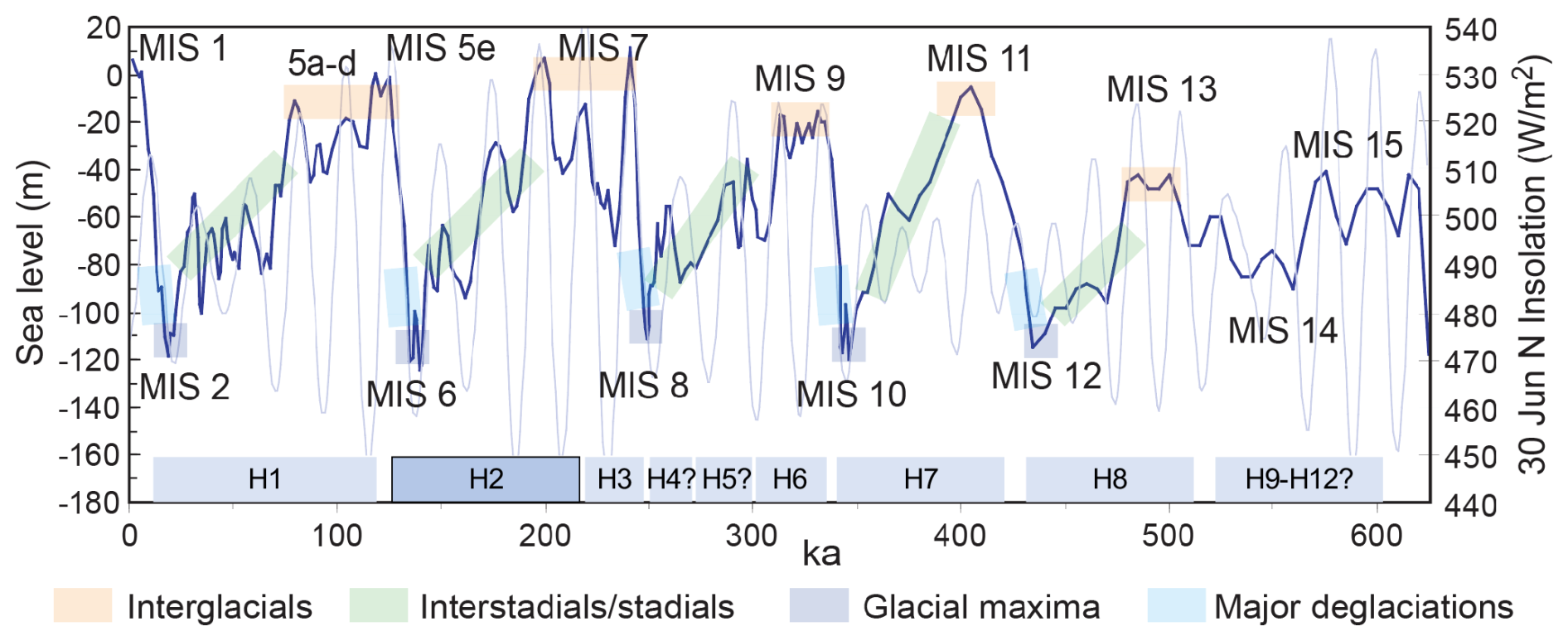
Figure 2Sea level, insolation and reef terraces around the Island of Hawai'i over the last 600 kyr (after Imbrie et al., 1984; Berger and Loutre, 1991; Lea et al., 2002). Observational and numerical modelling data indicate that the Hawai'ian reef successions (H1–H12) span each interglacial, interstadial/stadial, glacial maxima, and deglacial interval over the past 500–600 000 years with only minor hiatuses (Webster et al., 2009). The present study focuses on the H2 terrace.
The reefal microbialites in the Hawai'ian succession typically occur as the latest stage of framework sequence, overgrowing the coralgal framework and partly filling cavities in the reef or under ledges created by platy corals (Fig. 3). They form laminated and thrombolitic textures, in most cases occurring on top of red algae crusts that had previously encrusted corals (Webster et al., 2025). This is similar to previously described successions from other locations, such as Tahiti (e.g., Westphal et al., 2010) and the Great Barrier Reef (Braga et al., 2019), where microbialites were interpreted to develop coevally to, or immediately after coral growth, while the reef is still in the photic zone (Heindel et al., 2009b; Westphal et al., 2010; Jell and Webb, 2012). Although representing the latest stage of encrustation, these decimetres-thick microbialites do not completely fill the macropore space of the reef framework. In Tahiti, the microbial crusts were estimated to account for as much as 80 % of reef rock volume (Séard et al., 2011) and ∼43 %–72 % in the IODP 325 cores from the GBR (Szilagyi et al., 2020). As in the Tahitian microbialites, there is a distinct difference in coloration related to humidity and run-off patterns in the Hawai'ian microbialites, with the arid side of the island featuring lighter colours as compared to the humid side (Figs. 2 and A1; Webster et al., 2025).
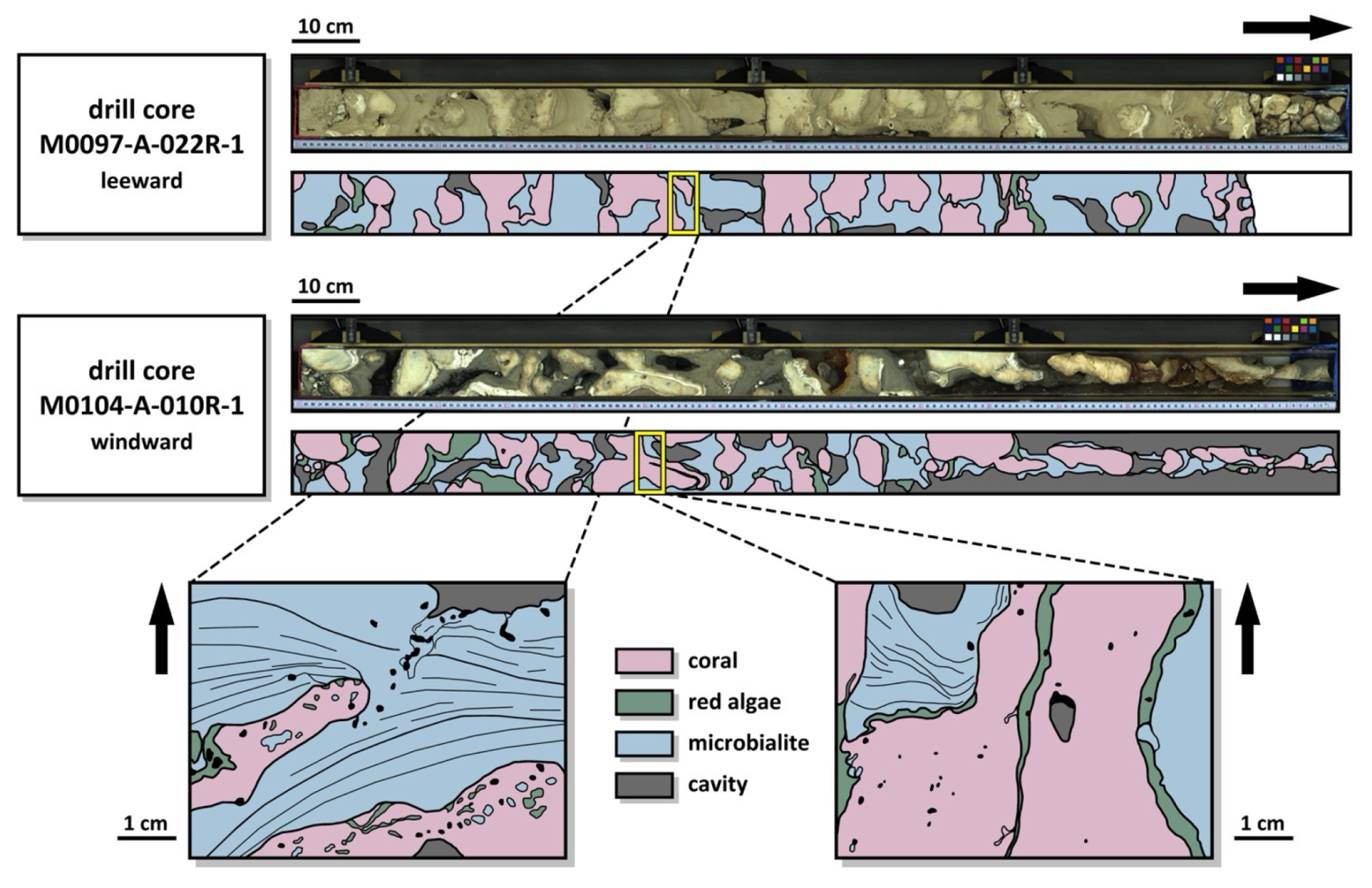
Figure 3Examples of core section line scans from the arid (M0097-A-022R-1) and humid side (M0104-A-010R-1) of the island of Hawai'i, along with the interpretation of coral, encrusting red algae, and microbialite contribution to the reef framework. The close-ups illustrate the occurrence of laminated microbialites within the reef framework.
In this study, we compared H2 terrace (MIS7 to MIS6; Webster et al., 2025) microbialites from the windward, humid side of the island (Sites M0102 and M0104) with those from the leeward, dry side (Site M0097) (Figs. 1, 3 and A1) specifically to identify any morphological biosignatures and document possible differences in microbial signatures between the two settings. A total of seven samples were chosen that have clear, undamaged accretionary surfaces where microbialites grew into reef framework voids, i.e., the final stage of growth (Fig. 4; Table B1). For identifying the microbialites, we followed the criteria of Webb and Kamber (2010), namely the macro-morphology, the micro-morphology, and the appropriate environment (see: Webster et al., 2025), with a special focus on the fourth criterion, i.e., the preservation of morphological bioindicators of microbes as part of the texture.

Figure 4Samples studied by SEM; blue quadrangles indicate the two subsamples prepared for SEM microscopy.
Samples were examined under a scanning electron microscope (SEM) to identify microbial features allowing for biological identification. With SEM imaging of microbialite surfaces and vertical polished sections, we characterize the ultrastructure of the crusts and identify trapped and bound particles (e.g., Cliona sponge chips from bored reef skeleton; de Bakker et al., 2024) as well as any microbial remains that might allow identification of the microbes, e.g., filaments that are indicative of taxonomic determination (cf., Melim et al., 2015).
Prior to analysis, the outermost growth surfaces of microbialites facing voids in the reef edifice were identified. These surfaces were sampled and mounted on SEM stubs. From the same samples, subsamples were cut perpendicular to the laminations and polished with corundum powder (borcarbid 400 and 600). All samples were then cleaned in an ultrasonic bath and rinsed with Millipore water, although in some cases, salts were not entirely removed. Cut samples were then etched with 0.3 % (0.1 N) hydrochloric acid for 20 s. Note that the preparation, including ultrasonic cleaning as well as the etching of the cut surfaces, prevents contamination. All samples were dried and coated with iridium (10 nm). SEM analysis with a secondary electron (SE) detector was performed with a Teneo-VS SEM at the KAUST Imaging and Characterization Core Lab, and a Tescan Vega LMU at the ZMT Geosciences Laboratory. Element concentrations were measured with an Oxford Energy Dispersive X-ray (EDX X-Max SDD; detector: 50 mm2) coupled to the Tescan Vega LMU. EDX working parameters were set to a beam voltage of 10 kV, a maximum of 5 million counts with a peak pile-up correction, and a process time of 4 min. Dead time was set to 30 %–50 %.
X-ray diffraction (XRD) pattern analyses were conducted in the laboratories of the Crystallography and Geomaterials Research Group at the Department of Geosciences, University of Bremen. Dried bulk samples were ground to a fine powder (<20 µm particle size) and prepared with a Philips backloading system. For XRD analysis, a Bruker D8 Discover diffractometer was used. The instrument was equipped with a Cu-tube (kα 1.541 Å, 40 kV, 40 mA), and a fixed divergence slit of °, a monochromatisation via energy discrimination on the highest resolution Linxeye detector system. Measurements were taken as a continuous scan from 3–65° 2θ, with a calculated step size of 0.016° 2θ. Mineral identification was done with the Philips software X'Pert HighScore™ Vs. 1.2 (Degen et al., 2014). The determination of well-crystallized minerals like quartz, calcite or aragonite has a standard deviation of ±1 %–3 % (Hardy and Tucker, 1988; Vogt et al., 2002).
4.1 Hilo and Kohala sites – Humid, windward margin
The microbialites from the humid side of Hawai'i are organized in layers, five of which can be distinguished on the basis of textures and biomorphic structures (Fig. 5A).

Figure 5Typical textures of layers composing microbialites on the humid side of the Island of Hawai'i. (A) Schematic of the five observed layers; (B, C) cut through the surface of the microbialite (Layer 1) showing fine crystalline texture and trapped grains; (D) collapsed surface layer revealing underlying Layer 2; (E) enlargement of Layer 2 showing filamentous biomorphs; (F) Layer 3 with altered and tightly cemented biomorphs; (G) view of Layers 1, 2, and 3 in a broken surface of a microbialite; (H, I) increase in crystal size of cements enclosing trapped and bound grains with cement showing peloidal textures in Layer 4; (J, K) peloidal texture and syntaxial aragonite needle cement on ascidian spicules in Layer 5 samples. Site M0104A, core 12.
Layer 1 (Fig. 5B and C): The surfaces of the microbialites are smooth, consisting of fine, poorly oriented anhedral crystals (Fig. 6A). This smooth, first layer exhibits a maximum thickness of 1 mm and consistently forms the uppermost element of the microbialite. The layer shows porous, cracked surfaces, which may represent a modern biofilm that has colonized the older microbialite (Fig. 6A–C). The samples contain abundant trapped and bound skeletal grains (e.g., Fig. 6B, black arrows). At high magnification, the outer layer locally appears to be composed of irregular material showing curved surfaces consistent with mineralised EPS or mucilage residues (Fig. 6D, black asterisks) and hollow filamentous microbial biomorphs emerging from beneath the layer (Fig. 6D, white arrows). Non-carbonate detritus is also present (Fig. 6E) showing an elemental composition pointing to silicates (Fig. C1). Cross-section samples from the windward, humid side of the island (Fig. 6F–J) reveal the deeper layers of the microbialite structure and show that the diffuse porosity observed on the surface continues below with interconnected voids (Fig. 6F).

Figure 6SEM images of samples from the humid side of the Island of Hawai'i. (A) Low magnification image of cross-section of the fine-crystalline surface. (B) Surface sample showing a collapsed area on the sample surface, revealing underlying Layer 2. (C) Higher magnification of the fine-crystalline surface layer material consistent with torn, desiccated, and mineralised EPS. (D) Mineralised EPS matrix (black asterisks) with mineralised filamentous microbial structures emerging from deeper layers as hollow tubes (white arrows), and residual embedded grains (black arrows). (E) Acicular microcrystals. (F) Porous spaces coated by microcrystals. (G) Details of torn, laminated matrix-like material consistent with fossilized EPS residues. (H) Hollow calcified sheaths of filamentous bacteria. (I) Microbial filaments located within a porous space (white arrow). (J) Calcified anastomosed tubes coated with crystals of various dimensions. (K) Flaky precipitates covering spherical biomorphs, (L) close-up of (K). (M) Pyrite aggregates. (E, J, M): Site M0102C, core section 22, all others: Site M0104A, core 12.
Layer 2 (Fig. 5D and E): Beneath the outer smooth, fine-crystalline Layer 1, calcified biofilms with preserved filaments occur. A remarkably well-preserved microbial mat shows calcified remains of EPS, isolated and aggregated filaments and a variety of spheroids (Fig. 7). Microbial bioindicators are abundant, including spheroidal and filamentous morphotypes (Fig. 7A–I; black and white arrows, respectively) embedded within the smooth, drape-like matrix. The filamentous morphotypes are horizontally oriented. Globular spheroids of variable size and morphology (Fig. 7B–F; white arrows) are consistently arranged in clustered aggregates. Smaller coccoid forms are also present (Fig. 7D and E). Filamentous structures exhibit a relatively uniform diameter (2–3 µm) and are predominantly preserved as hollow, calcified sheaths.
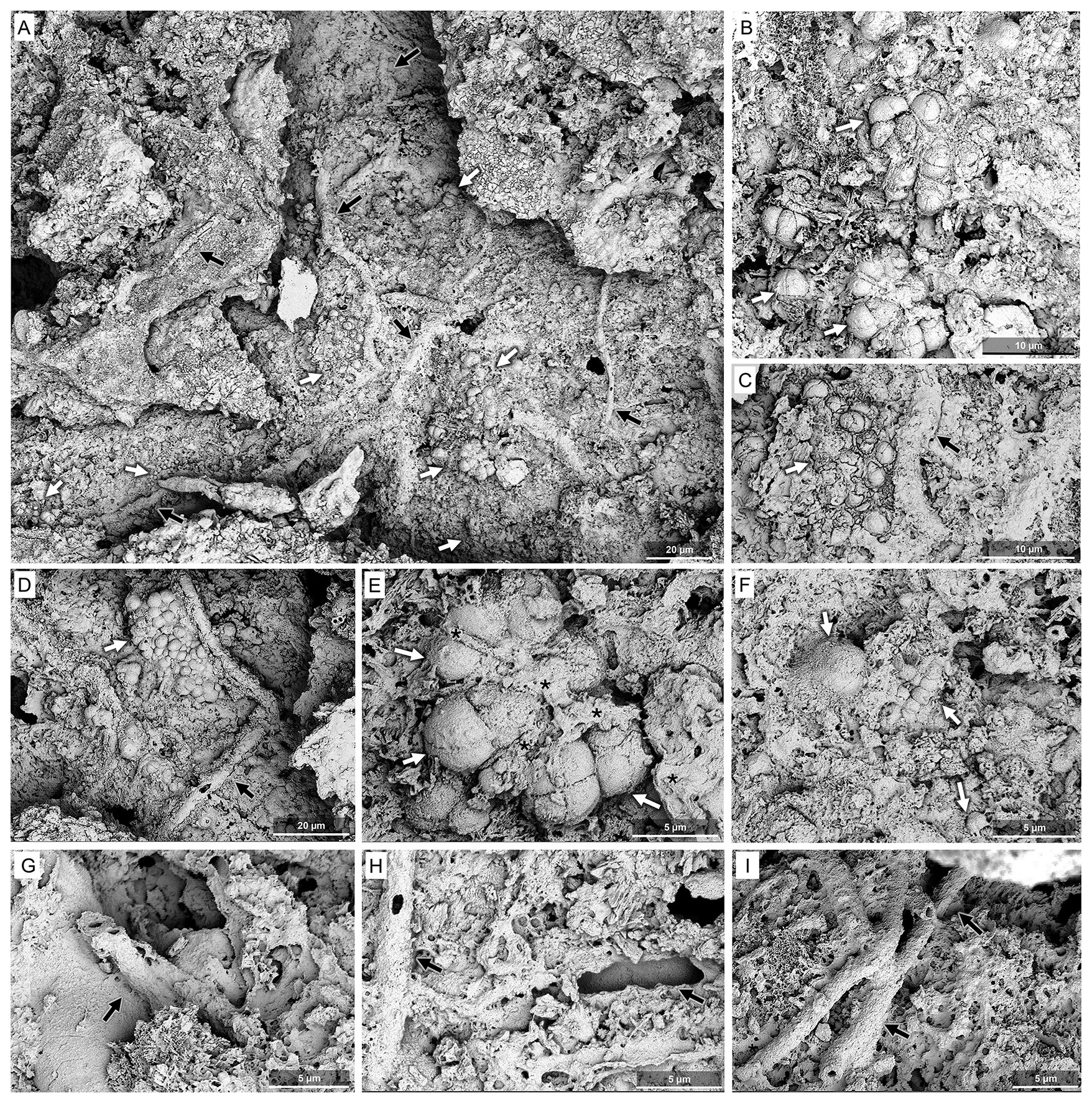
Figure 7SEM images of the microbial biofilm (Layer 2) observed on the surface sample from the humid side of the Island of Hawai'i. (A) General view of the crack within the surface layer exposing the biofilm; white arrows indicate coccid biomorphs and black arrows filamentous biomorphs. (B) Details of a group of coccoid biomorphs (white arrows) emerging from copious EPS matrix. (C, D) Details of a group of coccoid structures (white arrow) showing calcified EPS septa (black asterisks) and a filamentous biomorph (black arrow) in two different locations of the biofilm area. The quadrilobed biomorphs are similar to the modern colonial genus Gleocapsa. (E) Details of a group of coccoid biomorphs (white arrows) and hollow filaments (black arrow). (F) Details of a group of coccoid biomorphs (white arrows) and abundant EPS in a different spot of the biofilm area. (G–I) Mineralised hollow structures compatible with calcified sheaths of filamentous bacteria (black arrows). All images from Site M0104A, core 12.
Preserved biological structures at the surface include calcified EPS (Fig. 6G), hollow calcified microbial tubes and filaments (Fig. 6H and I, white arrows). The EPS residuals exhibited a filament-rich, matrix-like appearance, similar to that observed on the surface (Fig. 6G). Irregular needle-fibre crystal aggregates are consistent with thin fossilized EPS residuals, possibly at a more degraded stage compared to their surface counterparts (cf., Lin et al., 2019). The voids also contain anastomosed tubes (Fig. 6J). Locally, the tubes and spheroidal morphotypes show an overgrowth of flaky minerals (Fig. 6K and L, white arrows) with elemental composition of Ca, Mg, Al, and Si (Fig. C2).
Besides these general features, a remarkably well-preserved microbial mat shows clearly visible calcified EPS residuals, isolated and aggregated filaments and a variety of spheroids (Fig. 7). Microbial bioindicators are abundant, including spheroidal and filamentous morphotypes (Fig. 7A–I; black and white arrows, respectively) embedded within the smooth, drape-like matrix. The filamentous morphotypes are horizontally oriented. Globular spheroids of variable size and morphology (Fig. 7B–F; white arrows) are consistently arranged in clustered aggregates. Smaller coccoid forms are also present (Fig. 7D and E). Filamentous structures exhibit a relatively uniform diameter (2–3 µm) and are predominantly preserved as hollow, calcified sheaths (Fig. 7C, D, and G–I, black arrows).
Layer 3 (Fig. 5F and G): Further below that layer, microbial bioindicators such as filaments appear more altered, calcified and tightly cemented in the matrix.
Layer 4 (Fig. 5H and I): A downward coarsening of the cement crystals is observed in cross-section. Millimetres below the fine-crystalline surface layer, coarser crystals as well as peloidal textures appear (as seen in Figs. 5H, I and 6B, C). Peloidal textures are generally restricted to Layer 4 and below. The matrix encases rare nanocrystal aggregates (average aggregate dimension of 10 µm; as seen in Fig. 6D black arrows), as well as diffuse microcrystalline areas with irregularly oriented crystals forming peloids or aggregates (as seen in Fig. 6D, E, and M) and containing trapped and bound detritus, including arcuate chips of coral skeleton typical for the boring activity of the sponge Cliona, abundant ascidian spicules (Fig. 5H) and some calcareous sponge spicules. Pores are typically lined by more euhedral microcrystals (as seen in Fig. 6F, H, I, and J). Fine-grained meshes of acicular microcrystals (as seen in Fig. 6E and M) and bladed minerals (as seen in Fig. 6K and L) also occur. Pyrite framboids are present in Layer 4 (Fig. 6M).
Layer 5 (Fig. 5J and K): While otherwise showing the same textural characteristics as Layer 4, in Layer 5, aragonite needle cements are present as syntaxial overgrowth on aragonitic skeletal grains such as ascidian spicules and coral skeleton cut in chips by the boring sponge Cliona.
4.2 Interpretation of Hilo and Kohala sites – Humid, windward margin microbialites
The smooth undulating surface crusts (Layer 1) and increasing euhedral crystal growth as well as intergrown peloids with depth (downwards towards Layer 4, Fig. 5) are consistent with patterns observed in Holocene-modern microbialite growth (Webb et al., 1998). However, the well-preserved filaments and spheroids in Layer 2 with smooth, draping textures suggest abundant precipitation within the surface community where EPS and microbial sheaths were replaced by CaCO3 (Webb and Kamber, 2010, their Fig. 2). The large filament diameter and prostrate growth is consistent with cyanobacteria. The spheroids are more difficult to interpret. Some quadrilobed forms (Fig. 7B and E) closely resemble those observed in extant cyanobacteria of the order Chroococcales (e.g., the genus Gleocapsa, of similar dimensions). However, similar features have been produced in cyanobacterial biofilms as protodolomite spheroids, even mimicking a similar lobate morphology (e.g., Zhao et al., 2024).
The great difference in diameter of some of the spheroids in some clusters observed in Layer 2 (Fig. 7D) is more consistent with mineral spheroids, but other clusters are more uniform (Fig. 7B, C, and E). The mode of preservation may also favour mineral spheroids, as the filaments are preserved as calcified sheaths left hollow by degradation of the cells within, whereas the spheroids do not appear to be hollow, so would presumably represent preservation of the cells themselves. The large difference in diameter between spheroids of different clusters (Fig. 7A) suggests more than one process was operating in any case. Other holes in the EPS are consistent with microbial structure moulds. Regardless, both spheroid interpretations suggest the presence of cyanobacteria, so we suggest that the microbialites formed in open crypts with abundant light and adequate nutrients to support a cyanobacterial community as the primary producer of organic matter. Trapped and bound material still includes evidence of abundant cryptic reef dwellers (sponges and ascidians). Abundant coccoliths also occur, consistent with ambient water rich in nutrients. While precipitation occurred in the top layers within the microbial mat (Layers 1–3), thus preserving the filaments of cyanobacteria (oxygenic photoautotrophs), additional precipitation occurred deeper within the mat (Layer 4), where degraded EPS was possibly still preserved, and heterotrophs continued to increase alkalinity, coarsening microbial peloids and lining cavities with scalenohedra, through the activity of an anoxic community. The flaky precipitates with a silicate composition (Fig. C2) observed on biomorphs (Fig. 6K, L) could represent microbial autochthonous clay minerals. Preservation of pyrite framboids in deeper parts of the microbialite, for example, suggests ongoing sulphate reduction, but other processes, such as ammonification, also could have been important. The occurrence of syntaxial aragonite needle cements on skeletal grains suggests (Layer 5) a highly increased alkalinity allowing for precipitation below the main activity of sulphate-reducing bacteria.
4.3 Kawaihae site – Arid, leeward margin
The samples from the dry side of the island lack the distinctive layers observed on the humid side. While no fossilized microbial mats were observed here, other indicators of microbial activity are abundant. Both the growth surfaces (Fig. 8C and D) and the vertically cut samples (Fig. 8A, B, E, and F) show massive and peloidal textures composed of variably sized and typically irregularly oriented microcrystals. Peloidal structures as shown in Fig. 8D could have formed late in the microbialite owing to endolithic activity in the pore spaces under the rock surface (cf., Falkenberg et al., 2026). The bulk surface appears to be composed of irregularly overlapping laminae that are relatively smooth and undulose in places and more irregular with abundant trapped and bound sediment in others (Fig. 8C). Micropeloidal textures and confining structures are consistent with a biological origin (Fig. 8D–F) and occur in all samples coalescing to form the mineral matrix with minor trapped and bound detritus including both cryptic calcareous sponge spicules and possibly coccoliths (Fig. 8C). Much of the surface consists of irregular peloidal fabric composed of variably sized, mostly irregularly oriented crystals ranging from finer anhedral to coarser subhedral-euhedral shapes. Rare hollow, spherical structures coated in crystals (Fig. 8C; orange square) and tubular filamentous biomorphs (Fig. 8F) also occur, compatible with calcified bacterial cells and strings. One nannolith with a flowery appearance (Fig. 8C; yellow square) likely represents a coccolith.
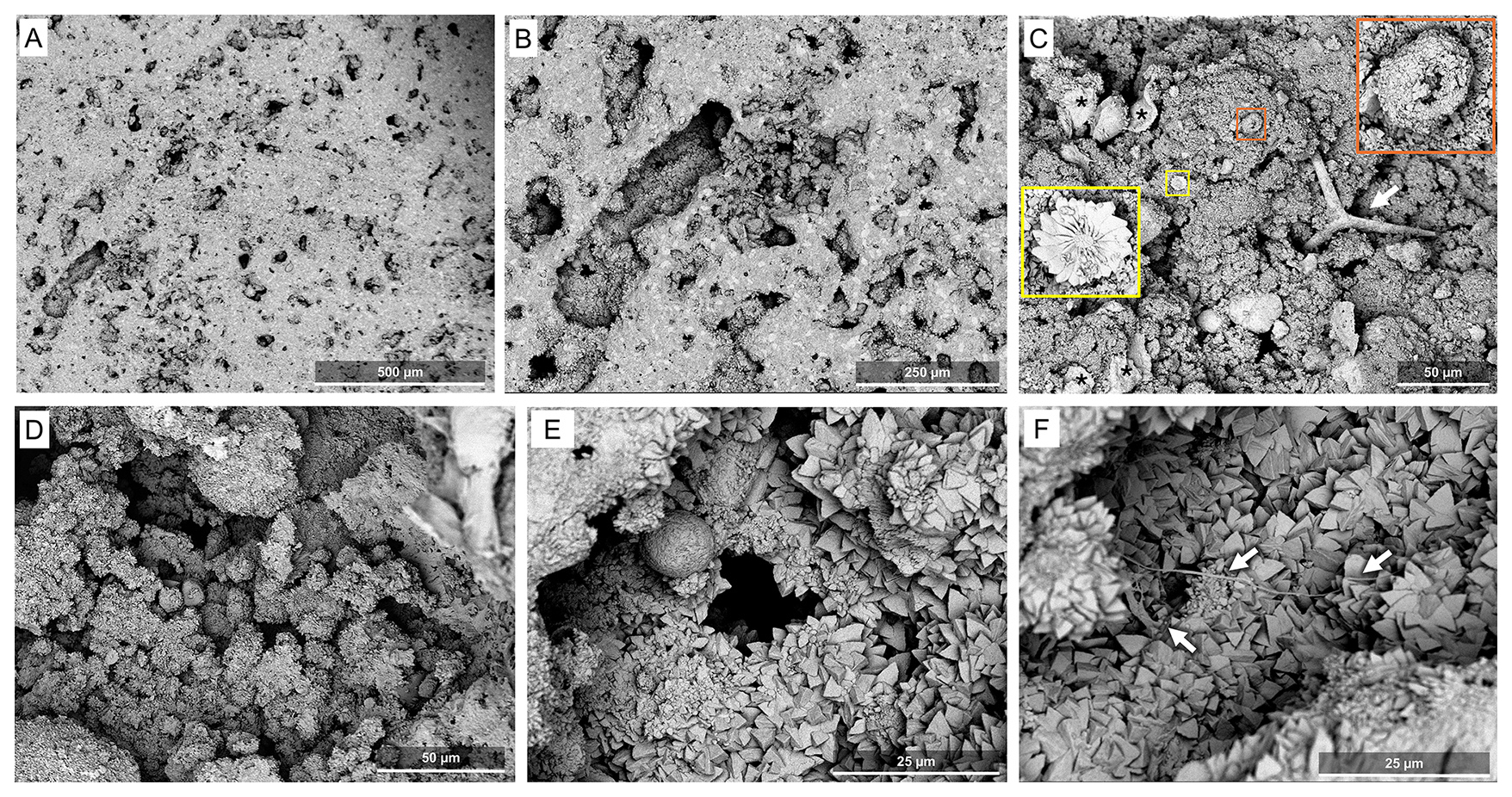
Figure 8SEM images of samples from the Kawaihae site, i.e., the dry side of the Island of Hawai'i. (A, B) Low magnification micrographs of the vertically cut sample. (C) Assemblage of structures of possible biological origin found on the surface of the sample; torn sheet-like matrices compatible with desiccated EPS (black asterisks), a spherical hollow structure consistent with a lithified coccoid microorganism (orange square), a calcareous sponge spicule (white arrow) and a coccolith flowery plate possibly affiliated to Discoasteraceae (yellow square). (D) Peloidal texture with abundant spheroidal structures coated in fine crystals. (E) Cavity exposed by the cut, covered in a microcrystalline coat. (F) Filaments consistent with EPS strings, indicated by white arrows (F). All images from Site M0097B, core 7.
Vertically cut samples are massive with porous zones serving to demarcate minor layering. Most of the micritic structure is composed of merged peloidal texture similar to that on the surface (Fig. 8A, B, and E). Trapped and bound detritus is relatively rare, but includes calcareous sponge spicules and chips excavated by the boring sponge Cliona from the skeleton of corals. The inner surfaces of voids are lined by coarser, more euhedral crystals (Fig. 8B, E, and F) as already described for the surface samples. Obvious biological structures and signatures are scarce here, consisting of spare microfilaments consistent with EPS fine strings (Fig. 8F). The presence of some characteristic smooth covers, in particular on the dry side of the islands, in both surface samples and cuts, may represent halite or organic matter remobilized during preparation.
4.4 Interpretation of Kawaihae site – Dry, leeward margin microbialites
The smooth undulating surfaces are consistent with formation at the base of a biofilm and are similar to other described reefal microbialites (Webb et al., 1998). The abundant peloids in deeper parts of the microbialites (millimeters depth) are typical of peloidal carbonates long associated with bacterial precipitation (e.g., Chafetz and Buczynski, 1992) and the coarsening of crystal texture below the surface is consistent with previous observations in reefal microbialites (Webb et al., 1998). The abundant holes or niches may have been colonized by endolithic microbial colonies, later covered with fine crystals coatings (cf., Falkenberg et al., 2026). The irregular fine-grained matrix is in places consistent with calcified EPS (Dohnalkova et al., 2011). Trapped and bound material from boring and other cryptic sponges, and the morphology described above is consistent with microbialite formation in semi-enclosed crypts within the reef framework, with no obvious evidence of cyanobacteria.
4.5 Mineralogy
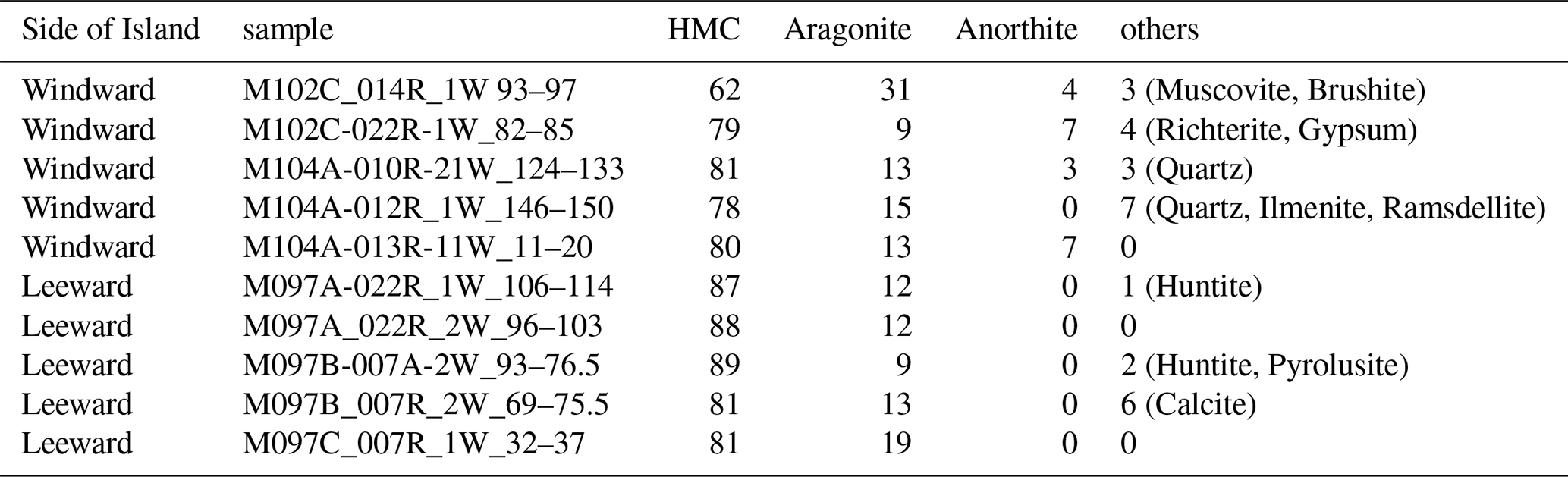
Bulk microbialite from both sides consists mainly of high-magnesium calcite (HMC), making as much as 89 % of the samples (Table D1). Aragonite amounts to as much as 15 %, while anorthite, i.e., Ca-rich plagioclase, is present (as much as 7 %) in samples from the windward side. Traces of quartz, huntite, and other minerals were also observed, but pyrite was not detected, despite framboidal pyrite being found in SEM analysis.
The genesis of reefal microbialites is still not fully understood. While the involvement of cyanobacteria in the formation of microbialites has been discussed for more than a century (e.g., Holtedahl, 1919), it became clear that complex consortia forming microbial mats and creating a succession of microbial activity are involved (e.g. Stolz et al., 2009). As Webb (2005) pointed out, microbialites are not necessarily the product of certain taxa but rather of a diverse consortium of autotrophic and heterotrophic microbes. Nevertheless, previous studies of Pleistocene reefal microbialites remained inconclusive regarding the presence of cyanobacteria (e.g., Heindel et al., 2009a).
The IODP Expedition 389 samples studied here resemble those presented in earlier studies (Heindel et al., 2010; Riding, 2011; Braga et al., 2019; Szilagyi et al., 2020) including the predominant composition of HMC and the textures observed in deeper parts of the microbialite, including the abundance of Cliona chips, originating from coral skeletons, as reflected, together with some cements, in the aragonite portion reaching 15 % of the bulk microbialite. However, in contrast to reefal microbialites studied earlier, the Hawai'ian microbialites exhibit an extraordinary preservation of microbial mats, featuring cyanobacterial bioindicators. In particular, the samples from the humid side of the Island of Hawai'i show rich bioindicative petrological features. The abundant occurrence of desiccated, cracked and calcified EPS or mucilage residues in surface samples indicates that they indeed are ancient features and have a distinctly different appearance than scattered spots of younger organic material accumulated during preparation on surfaces and cuts. This is also confirmed by the presence of degraded EPS signatures within the rock as exposed in vertical cuts of the samples.
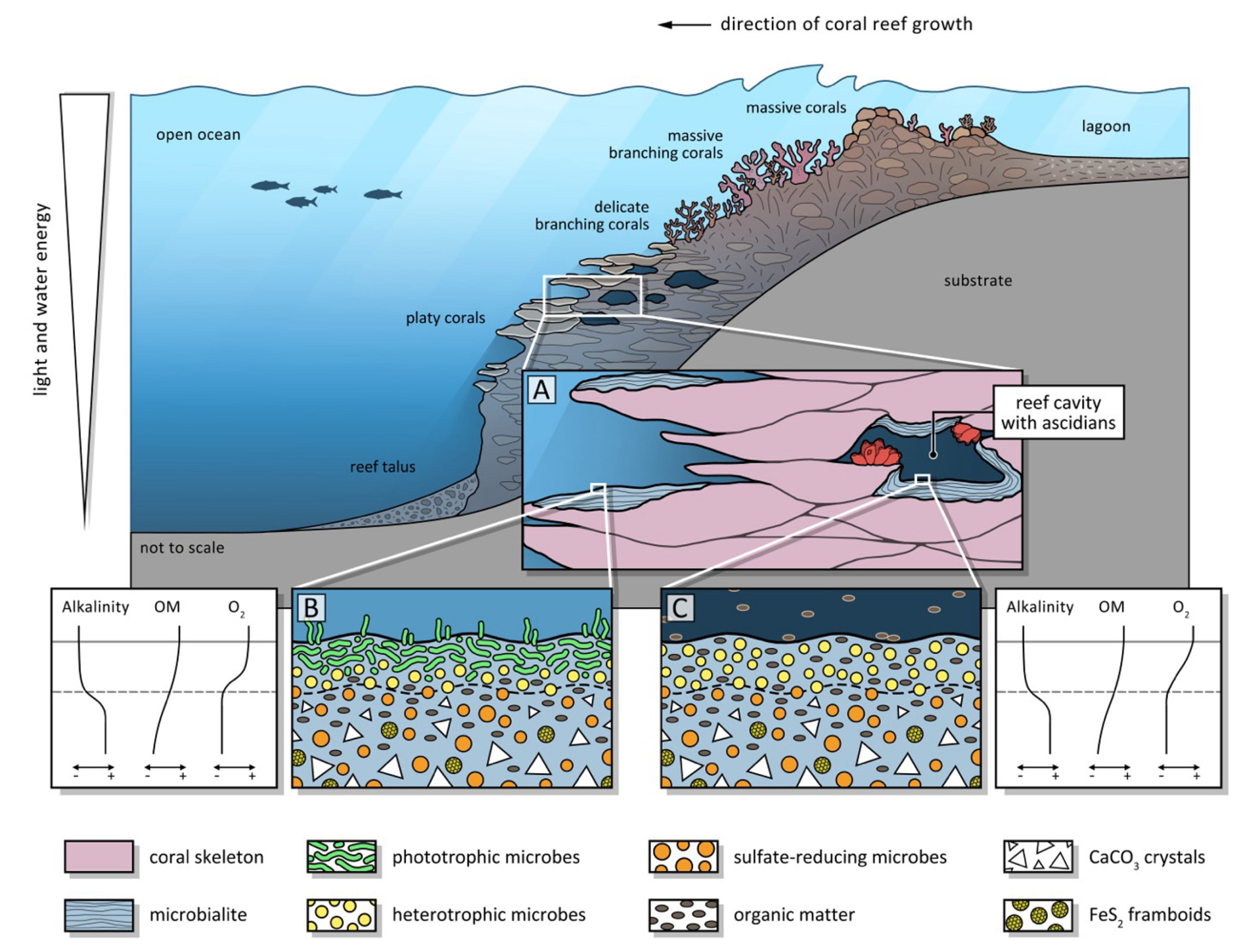
Figure 9Schematic diagram of loci of microbialite formation in a coral reef under slightly shaded euphotic versus oligophotic cryptic conditions. Section through a coral reef, (A) euphotic versus cryptic setting in a cavity, (B) close-up of euphotic setting with cyanobacterial mat at the surface of the microbialite, (C) close-up of detrital organic matter-driven microbial mat in a cavity. Diagrams illustrating gradients in alkalinity, organic matter, and oxygen throughout the upper layers of the microbial mats.
IODP 389 microbialite samples reveal a succession of textures and bioindicators allowing for reconstruction of the successive microbial activities in the succeeding layers (Fig. 5). Figure 7 shows a microbialite surface from the humid side that exhibits a specifically copious and well-preserved layer of calcified EPS. The calcified EPS and biofilm show a “cracked” appearance of the uppermost layer of the surface specimen which, at closer look, is made of torn EPS matrix embedding hollow filamentous sheaths. In the same specimens, pyrite aggregates indicate a reducing micromilieu, caused by heterotrophic oxygen removal, that allowed sulphate reduction within and/or beneath the microbial mats and the formation of local redox gradients. This is consistent with the findings of Gomes et al. (2022) who showed microbial communities are likely to play a major role in promoting pyrite formation at the surface of microbial mats. The origin of traces of quartz identified in XRD measurements of samples from the windward side of the island is inconclusive. It may be autochthonous, representing remineralised diatom and siliceous sponge skeletons, or may be allochthonous sourced from Asian eolian dust as identified previously on Hawai'i (Kurtz et al., 2001). Quartz was not observed directly in SEM, so its morphology remains unknown.
The observed biofilm areas (Figs. 5–7) are particularly well-preserved and allow characterization of a succession from fine-crystalline, smooth surface layers to “deeper” layers of the biofilm, followed by even deeper and more crystallized layers. Filaments that extend into the embedding EPS and sediments exclude their being modern contaminants. The putative coccoid colonies are also completely embedded in the matrix, which shows the typical appearance of dehydrated EPS with calcified septa in some cell clusters. These spectacular microbial mats with diverse cell types (no bacilli, only globular/cocci and filamentous) and colonies support the hypothesis of an actual microbial multispecies biofilm or microbial mat, some of them consistent with cyanobacteria indicating that the microbialite originated in the photic zone.
In the samples from the arid side of the Island of Hawai'i, discrete layers of biomats were not identified, but similar biosignatures are present, albeit less abundant than on the humid side. Also, fewer spicules of ascidians were identified. Encrusting ascidians are common inhabitants of reef cavities as shown for Lizard Island in the GBR, with their aragonitic spicules being incorporated in microbialites (Reitner, 1993). As ascidians thrive in moderate to elevated nutrient levels (e.g., Ribes et al., 2005; Eckhardt et al., 2024) a lower abundance on the arid side of the island might be expected.
The flaky and mesh-like minerals observed (Fig. 6K–M) resemble the magnesium carbonate huntite known from microbialites forming in high-salinity/alkalinity crater lakes (Caumartin et al., 2025), or the magnesium carbonate dypingite that is known to be associated with modern cyanobacteria in alkaline wetlands (Power et al., 2007; Shirokova et al., 2013). It is known that EPS secreted by cyanobacteria can bind Mg2+, creating microenvironments conducive to mineral nucleation (Robles-Fernández et al., 2022). Dypingite, however, does not form in marine waters and has never been confirmed in reefal microbialites. Also, Mg clays can have a similar appearance and are known to form in the context of cyanobacteria (Perri et al., 2018). Therefore, an interpretation remains speculative. However, XRD analyses indicate the presence of traces of huntite, which is known to form in microbialites (Caumartin et al., 2025). The observation that HMC is so ubiquitous in reefal microbialites in general (Reitner, 1993; Camoin et al., 1999; Gischler et al., 2017) and in the Hawai'ian examples studied here in particular lends evidence to the presence of heterotrophic bacteria as well. Sulfate-reducing bacteria apparently exert control on calcium carbonate polymorph mineralogy in that HMC and dolomite precipitation occur preferentially, both in experiments and in a modern coastal lagoon environment (van Lith et al., 2003).
Evidence found in the MIS 7–6 wet-side microbialite samples from IODP 389 allows us to reconstruct the microbialites as the product of photic biomats that featured abundant cyanobacteria, while a few millimeters below this exposed surface, sulphate reducing and other heterotrophic microbes are suspected to drive larger scale calcification, producing peloidal textures typical for microbialites, and evidenced by pyrite framboids (Fig. 5). They seem to drive oversaturation of the porewater with respect to HMC as evidenced in the peloidal texture, and with aragonite in deeper layers of the microbialite where it precipitates as syntaxial needle cement. While this model seems conclusive for the humid, windward side of the island, its applicability on the arid side is not fully coherent, because the bioindicators of cyanobacteria have not been found. The question remains open as to why the record recovered on the humid side of the Island of Hawai'i contains such well-preserved cyanobacterial traces compared to the arid side, as well as to the substantially younger, last deglacial successions recovered in Tahiti (IODP 310) and the glacial maximum to deglacial aged ones from the GBR (IODP 325) (Westphal et al., 2010; Braga et al., 2019). The microbialites lacking cyanobacterial remains could represent a fully cryptic, aphotic setting where cyanobacteria did not thrive. Figure 9 schematically illustrates the different environments in a coral reef where microbialites may form under varying photic conditions. While aphotic microbialites are known from modern reef cavities and are typically characterized by abundant sponge or ascidian spicules (Reitner, 1993; Gischler et al., 2017; Maak et al., 2024), in the present study, on the humid side of Hawai'i, ascidians cooccur with cyanobacteria, pointing to shaded and more nutrient-rich settings between platy corals in the euphotic zone. For the arid side of Hawai'i, either an aphotic cryptic origin of the microbialites or poor preservation of the surface mat could explain the absence of preserved biomorphs. This study emphasizes the point that reefal microbialites represent a diverse set of precipitates that may have different origins in different environmental settings within the reef. Understanding their differences and similarities should render them more useful as environmental archives.

Figure A1Line scans of H2-terrace reef cores studied here by means of SEM. Core width is 7.2 cm.
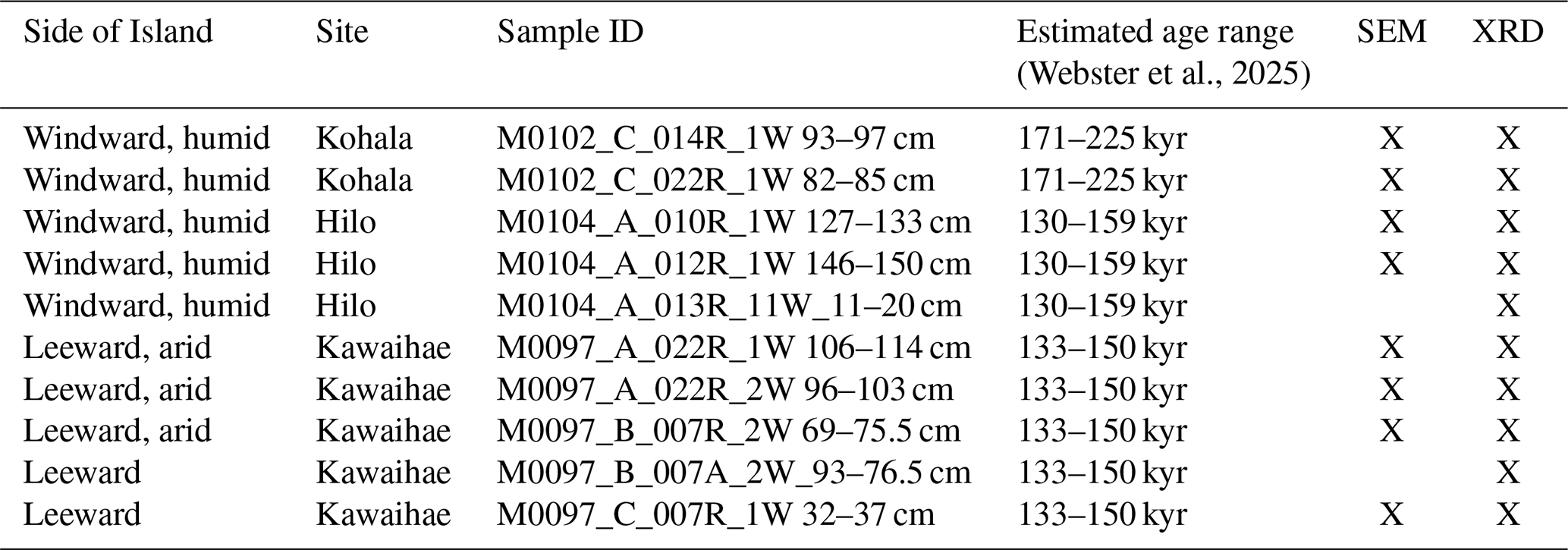
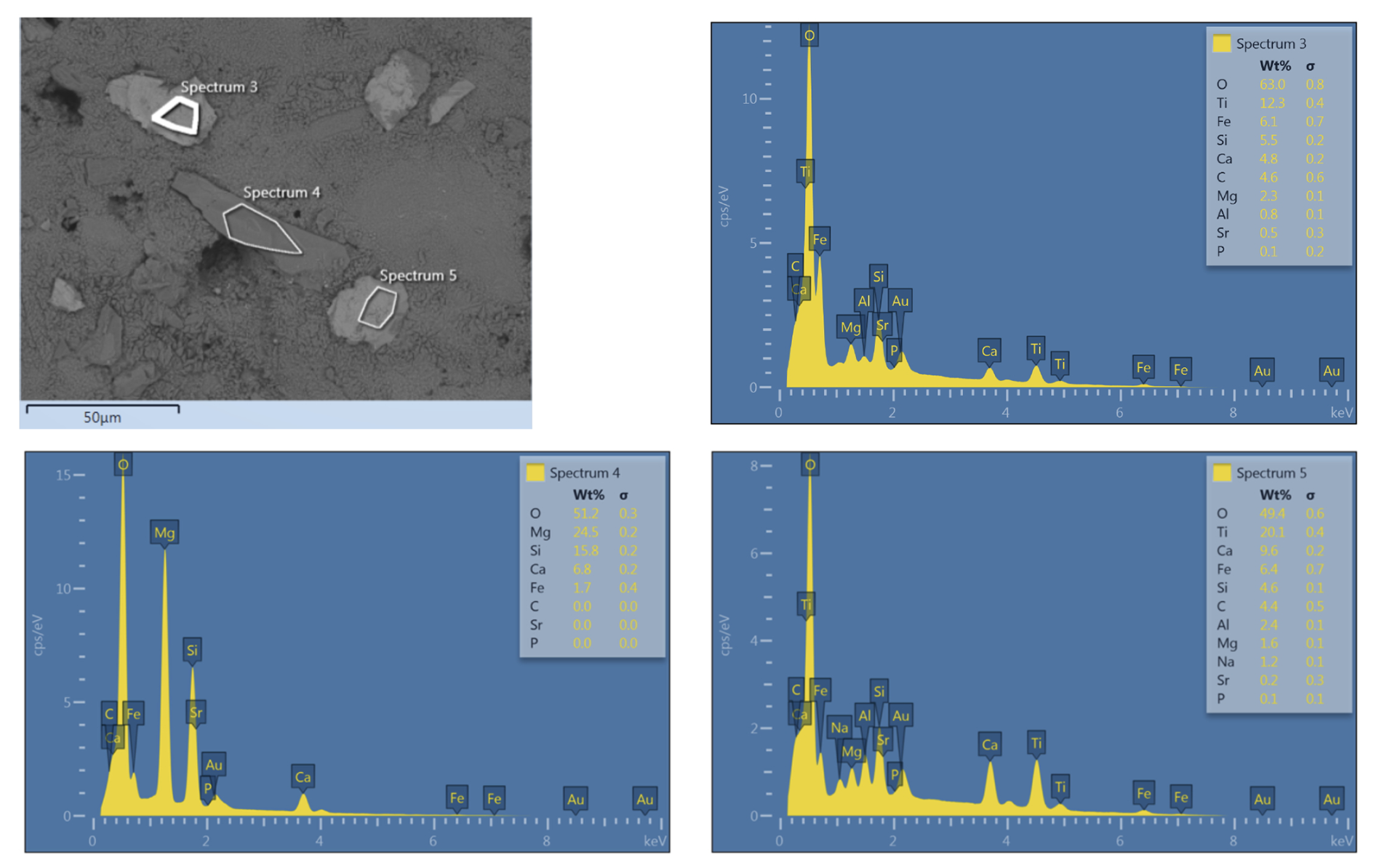
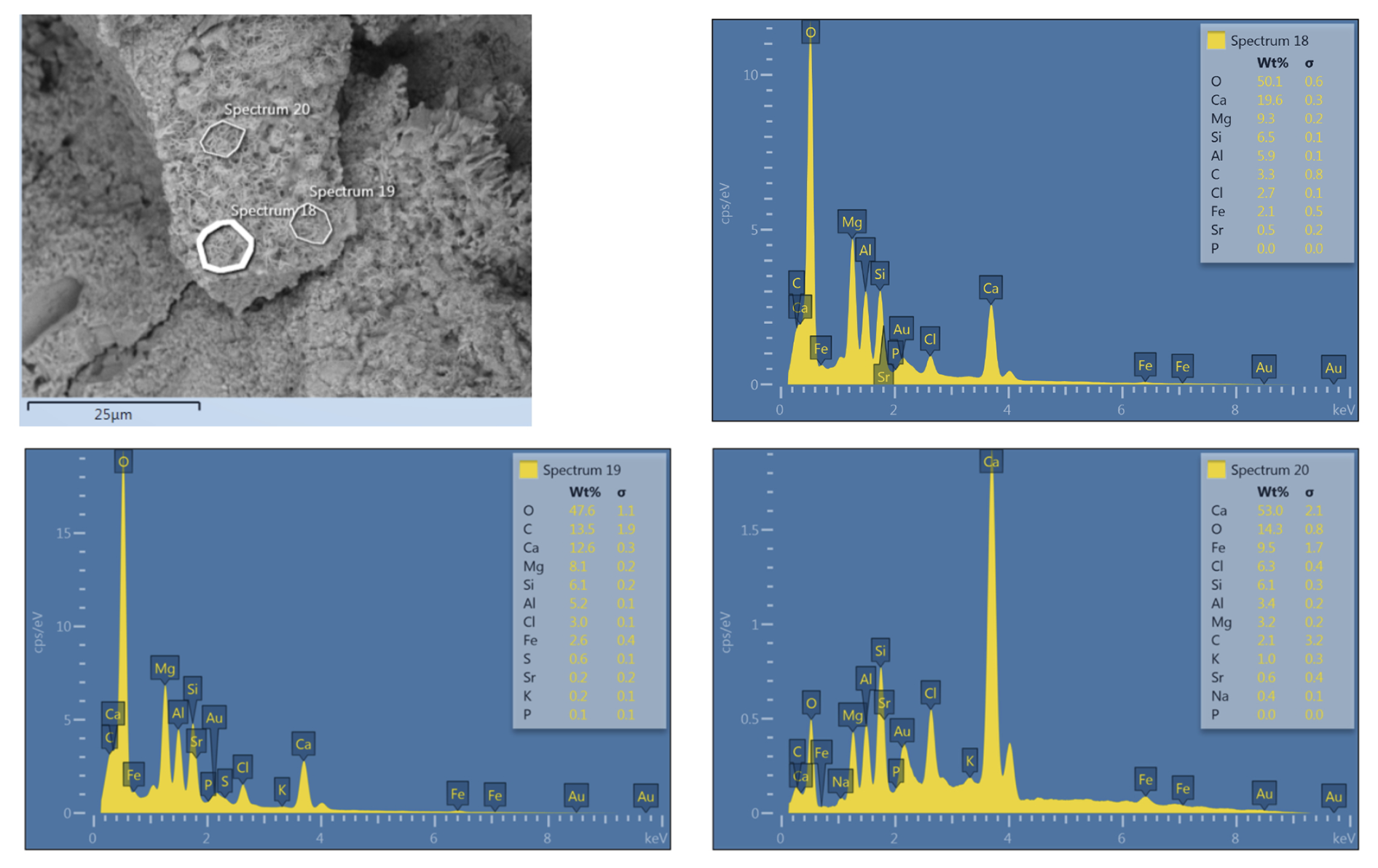
Table D1Mineralogical composition according to XRD results in %. HMC indicates high-Magnesium calcite.
The SEM micrographs are openly available from the PANGAEA data repository (https://doi.org/10.1594/PANGAEA.995587, Westphal et al., 2026).
JMW acted as co-chief scientist of expedition IODP 389 and was thus instrumental in designing and conducting the cruise and securing the core material; HW designed the present study; HW prepared the sample material; HW and EGa conducted the SEM and EDX analyses; HW, EGa and GEW interpreted the data with contributions by all authors; AM did the artwork in Figs. 5 and 9; HW drafted the first version of the manuscript; all co-authors substantially contributed to the manuscript and the revision.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
This publication benefitted from reviews by Nora Noffke and Joachim Reitner. We are grateful to Sebastian Flotow for support with the preparation of the samples and operation of the SEM at ZMT. Maximilian Göbel (Bremen University) helped with the preparation of the samples and the SEM work at ZMT. This work is part of the research to fulfil the obligations of the IODP X389 Science party, of which HW, PKh, TN, EGi, and JMW are members. We are indebted to the IODP, Bremen Core Repository (MARUM, University of Bremen), and British Geological Service personnel supporting the work on IODP X389, as well as to the PANGAEA team and Finn Opätz (ZMT) for support with the data management.
This research was funded by ZMT baseline funding to HW. JMW and GEW acknowledge support provided by the Australian Research Council (grant no. DP250102180) and ANZIC (grant no. PCAFX38901).
The article processing charges for this open-access publication were covered by the University of Bremen.
This paper was edited by Mark Lever and reviewed by Nora Noffke and Joachim Reitner.
Allwood, A. C., Walter, M. R., Kamber, B. S., Marshall, C. P., and Burch, I. W.: Stromatolite reef from the Early Archaean era of Australia, Nature, 441, 714–718, 2006.
Baud, A., Cirilli, S., and Marcoux, J.: Biotic response to mass extinction: the lowermost Triassic microbialites, Facies, 36, 238–242, 1997.
Berger, A. and Loutre, M. F.: Insolation values for the climate of the last 10 million years, Quaternary Sci. Rev., 10, 297–317, https://doi.org/10.1016/0277-3791(91)90033-Q, 1991.
Braga, J. C., Puga-Bernabéu, Á., Heindel, K., Patterson, M. A., Birgel, D., Peckmann, J., Sánchez-Almazo, I. M., Webster, J. M., Yokoyama, Y., and Riding, R.: Microbialites in Last Glacial Maximum and deglacial reefs of the Great Barrier Reef (IODP Expedition 325, NE Australia), Palaeogeogr. Palaeocl., 514, 1–17, 2019.
Burne, R. V. and Moore, L. S.: Microbialites: Organosedimentary deposits of benthic microbial communities, Palaios, 2, 241–254, 1987.
Camoin, G. F. and Montaggioni, L.: High energy coralgal-stromatolite frameworks from Holocene reefs (Tahiti, French Polynesia), Sedimentology, 41, 655–676, 1994.
Camoin, G. F., Gautret, P., Montaggioni, L., and Cabioch, G.: Nature and environmental significance of microbialites in Quaternary reefs: the Tahiti paradox, Sediment. Geol., 126, 271–304, 1999.
Castanier, S., Le Metayer-Levrel, G., and Perthuisot, J.-P.: Ca-carbonate precipitation and limestone genesis – the microbiogeologist point of view, Sediment. Geol., 126, 9–23, https://doi.org/10.1016/S0037-0738(99)00028-7, 1999.
Caumartin, J., Benzerara, K., Havas, R., Thomazo, C., Fogret, L., Betancourt, V., Tavera, R., Doisneau, B., Jézéquel, D., and Lopez-Garcia, P.: Huntite [CaMg3(CO3)4], a rare carbonate phase formed during early diagenesis in modern microbialites, ACS Earth Space Chem., 9, https://doi.org/10.1021/acsearthspacechem.5c00022, 2025.
Chafetz, H. and Buczynski, C.: Bacterially Induced Lithification of Microbial Mats. Palaios, 7, 277–293, https://doi.org/10.2307/3514973, 1992.
de Bakker, D. M., Perry, C. T., Magana-Gallegos, E., Perez-Cervantes, E., and Alvarez-Filip, L.: Fine-grained sediment production by endolithic sponges on Caribbean coral reefs, Limnol. Oceanogr., 69, 2015–2028, https://doi.org/10.1002/lno.12640, 2024.
Degen, T., Sadki, M., Bron, E., König, U., and Nénert, G.: The HighScore suite, Powder Diffr., 29, S13–S18, https://doi.org/10.1017/S0885715614000840, 2014.
Dohnalkova, A. C., Marshall, M. J., Arey, B. W., Williams, K. H., Buck, E. C., and Fredrickson, J. K.: Imaging hydrated microbial extracellular polymers: comparative analysis by electron microscopy, Appl. Environ. Microb., 77, 1254–1262, 2011.
Dupraz, C. and Strasser, A.: Nutritional modes in coral-microbialite reefs (Jurassic, Oxfordian, Switzerland): Evolution of trophic structure as a response to environmental change, Palaios, 17, 449–471, 2002.
Dupraz, C. and Visscher, P. T.: Microbial lithification in marine stromatolites and hypersaline mats, Trends Microbiol., 13, 429–438, 2005.
Dupraz, C., Reid, R. P., Braissant, O., Decho, A. W., Norman, R. S., and Visscher, P. T.: Processes of carbonate precipitation in modern microbial mats, Earth-Sci. Rev., 96, 141–162, 2009.
Eckhardt, S., Ainsworth, T. D., Leggat, W., and Page, C. E.: Colonial ascidian populations at inshore coral reefs of Norfolk Island, South Pacific, Diversity, 16, 384, https://doi.org/10.3390/d16070384, 2024.
Falkenberg, P., Vahrenkamp, S., Garuglieri, E., Petrovic, A., Hachmann, K., Chandra, V., and Vahrenkamp, V.: Polygonal tepee structures of Arabia, The Depositional Record, 12, e70043, https://doi.org/10.1002/dep2.70043, 2026.
Garuglieri, E., Marasco, R., Odobel, C., Chandra, V., Teillet, T., Areias, C., Sánchez-Román, M., Vahrenkamp, V., and Daffonchio, D.: Searching for microbial contribution to micritization of shallow marine sediments, Environ. Microbiol., 26, e16573, https://doi.org/10.1111/1462-2920.16573, 2024.
Gischler, E., Heindel, K., Birgel, D., Brunner, B., Reitner, J., and Peckmann, J.: Cryptic biostalactites in a submerged karst cave of the Belize Barrier Reef revisited: pendant bioconstructions cemented by microbial micrite, Palaeogeogr. Palaeocl., 468, 34–51, 2017.
Gomes, M. L., Klatt, J. M., Dick, G. J., Grim, S. L., Rico, K. I., Medina, M., Ziebis, W., Kinsman-Costello, L., Sheldon, N. D., and Fike, D. A.: Sedimentary pyrite sulfur isotope compositions preserve signatures of the surface microbial mat environment in sediments underlying low-oxygen cyanobacterial mats, Geobiology, 20, 60–78, https://doi.org/10.1111/gbi.12466, 2022.
Grotzinger, J. P. and Knoll, A. H.: Stromatolites in Precambrian carbonates: Evolutionary mileposts or environmental dipsticks?, Annu. Rev. Earth Pl. Sc., 27, 313–358, 1999.
Hardy, R. G. and Tucker, M.: X-ray powder diffraction of sediments, in: Techniques in sedimentology, edited by: Tucker, M., Oxford Blackwell, 191–228, ISBN 0632013729, 1988.
Heindel, K., Birgel, D., Peckmann, J., Kuhnert, H., and Westphal, H.: Sulfate-reducing bacteria as major players in the formation of reef-microbialites during the last sea-level rise (Tahiti, IODP 310), Geochim. Cosmochim. Ac., 73, Goldschmidt Conference A514-A514, 2009a.
Heindel, K., Westphal, H., and Wisshak, M.: Bioerosion in the reef framework, IODP Expedition #310 off Tahiti (Tiarei, Mara'a, and Faa'a), in: Proceedings IODP, 310, edited by: Camoin, G. F., Iryu, Y., McInroy, D. B., and the Expedition 310 Scientists, 28 p. https://doi.org/10.2204/iodp.proc.310.201.2009, 2009b.
Heindel, K., Birgel, D., Peckmann, J., Kuhnert, H., and Westphal, H.: Formation of deglacial microbialites in coral reefs off Tahiti (IODP 310) involving sulfate-reducing bacteria, Palaios, 25, 618–635, https://doi.org/10.2110/palo.2010.p10-032r, 2010.
Heindel, K., Birgel, D., Brunner, B., Thiel, V., Westphal, H., Gischler, E., Ziegenbalg, S. B., Cabioch, G., Sjövall, P., and Peckmann, J.: Post-glacial microbialite formation in coral reefs in the Pacific, Atlantic, and Indian Ocean, Chem. Geol., 304–305, 117–130, https://doi.org/10.1016/j.chemgeo.2012.02.009, 2012.
Holtedahl, O.: The Paleozoic formations of Finmarken in northern Norway, Am. J. Sci., 4, 85–107, 1919.
Imbrie, J., Hays, J. D., Martinson, D. G., McIntyre, A., Mix, A. C., Morley, J. J., Pisias, N. G., Prell, W. L., and Shackleton, N. J.: The orbital theory of Pleistocene climate: support from a revised chronology of the marine δ18O record, in: Milankovitch and climate: understanding the response to orbital forcing, edited by: Berger, A., Imbrie, J., Hays, J., and Kukla, G., D. Reidel Publishing Company, 269–305, ISBN 978-9027717771, 1984.
Jell, J. S. and Webb, G. E.: Geology of heron island and adjacent reefs, Great Barrier Reef, Australia, Episodes, 35, 110–119, 2012.
Kurtz, A. C., Derry, A. D., and Chadwick, O. A.: Accretion of Asian dust to Hawaiian soils: isotopic, elemental, and mineral mass balances, Geochim. Cosmochim. Ac., 65, 1971–1983, https://doi.org/10.1016/S0016-7037(01)00575-0, 2001.
Lea, D. W., Martin, P. A., Pak, D. K., and Spero, H. J.: Reconstructing a 350 kyr history of sea level using planktonic Mg/Ca and oxygen isotope records from a Cocos Ridge core, Quaternary Sci. Rev., 21, 283–293. https://doi.org/10.1016/S0277-3791(01)00081-6, 2002.
Lin, F., Zhu, X., Li, J., Yu, P., Luo, Y., and Liu, M.: Effect of extracellular polymeric substances (EPS) conditioned by combined lysozyme and cationic polyacrylamide on the dewatering performance of activated sludge, Chemosphere, 235, 679–689, 2019.
Maak, J. M., Birgel, D., Reitner, J., Gischler, E., Dullo, W. C., Foster, W., and Peckmann, J.: Molecular fossils in microbial carbonates and sponges of the deep fore reef of Mayotte and Moheli, Comoro Islands, Facies, 70, https://doi.org/10.1007/s10347-023-00678-3, 2024.
Marshall, J. F. and Davies, P. J.: Halimeda bioherms of the northern Great Barrier Reef, Coral Reefs, 6, 139–148, 1988.
Melim, L. A., Northup, D. E., Spilde, M. N., and Boston, P. J.: Update: Living reticulated filaments from Herbstlabyrinth-Adventhöhle Cave System Germany, J. Cave Karst Stud., 77, 87–90, 2015.
Noffke, N. and Awramik, S. M.: A tale of two microbialites: Stromatolites and microbially induced sedimentary structures, Sedimentology, https://doi.org/10.1111/sed.70109, 2026.
Nothdurft, L. D., Webb, G. E., Bostrom, T., and Rintoul, L.: Calcite-filled borings in the most recently deposited skeleton in live-collected Porites (Scleractinia): implications for trace element archives, Geochim. Cosmochim. Ac., 71, 5423–5438, https://doi.org/10.1016/j.gca.2007.09.025, 2007.
Pei, Y., Hagdorn, H., Voigt, T., Duda, J.-P., and Reitner, J.: Palaeoecological Implications of Lower-Middle Triassic Stromatolites and Microbe-Metazoan Build-Ups in the Germanic Basin: Insights into the Aftermath of the Permian–Triassic Crisis, Geosciences, 12, 133, https://doi.org/10.3390/geosciences12030133, 2022.
Perri, E., Tucker, M. E., Słowakiewicz, M., Whitaker, F., Bowen, L., and Perrotta, I. D.: Carbonate and silicate biomineralization in a hypersaline microbial mat (Mesaieed sabkha, Qatar): Roles of bacteria, extracellular polymeric substances and viruses, Sedimentology, 65, 1213–1245, https://doi.org/10.1111/sed.12419, 2018.
Power, I. M., Wilson, S. A., Thom, J. M., Dipple, G. M., and Southam, G.: Biologically induced mineralization of dypingite by cyanobacteria from an alkaline wetland near Atlin, British Columbia, Canada, Geochem. T., 8, 13, https://doi.org/10.1186/1467-4866-8-13, 2007.
Pratt, B. R.: Stromatolite decline – a reconsideration, Geology, 10, 512–515, 1982.
Puga-Bernabéu, Á., Webster, J. M., Braga, J. C., Clague, D. A., Dutton, A., Eggins, S., Fallon, S., Jacobsen, G., Paduan, J. B., and Potts, D. C.: Morphology and evolution of drowned carbonate terraces during the last two intergla-cial cycles, off Hilo, NE Hawaii, Mar. Geol., 371, 57–81, https://doi.org/10.1016/j.margeo.2015.10.016, 2016.
Reid, P., Visscher, T. P., Decho, A. W., Stolz, J. F., Bebout, B. M., Dupraz, C., MacIntyre, I. G., Paerl, H. W., Pinckney, J. L., Prufert-Bebout, L., Steppe, T. F., and DesMarais, D. J.: The role of microbes in accretion, lamination, and early lithification of modern marine stromatolites, Nature, 406, 989–992, 2000.
Reid, R. P., Suosaari, E. P., Oehlert, A. M., Pollier, C. G. L., and Dupraz, C.: Microbialite accretion and growth: lessons from Shark Bay and The Bahamas, Annu. Rev. Mar. Sci., 16, 487–511, 2024.
Reitner, J.: Modern cryptic microbialite-metazoan facies from Lizard Island (Great Barrier Reef, Australia), formation and concepts, Facies, 29, 3–40, 1993.
Reitner, J., Gautret, P., Marin, F., and Neuweiler, F.: Automicrites in a modern microbialite. Formation model via organic matrices (Lizard Island, Great Barrier Reef, Australia), Bull. Inst. Océanogr. Monaco, 14, 237–263, 1995.
Reitner, J., Thiel, V., Zankl, H., Michaelis, W., Wörheide, G., and Gautret, P.: Organic and biogeochemical patterns in cryptic microbialites, in: Microbial sediments, Springer Berlin Heidelberg, Berlin, Heidelberg, 149–160, https://doi.org/10.1007/978-3-662-04036-2_17, 2000.
Ribes, M., Coma, R., Atkinson, M. J., and Kinzie III, R. A.: Sponges and ascidians control removal of particulate organic nitrogen from coral reef water, Limnol. Oceanogr., 50, 2005, 1480–1489, 2005.
Riding, R.: Microbial carbonates: the geological record of calcified bacterial-algal mats and biofilms, Sedimentology, 47, 179–214, 2000.
Riding, R.: Microbial carbonate abundance compared with fluctuations in metazoan diversity over geological time, Sediment. Geol., 185, 229–238, 2006.
Riding, R.: Microbialites, stromatolites, and thrombolites, in: Reefs in Time and Space: The History and Evolution of the Reef-Building Biota, 103–120, https://doi.org/10.1007/978-1-4020-9212-1_196, 2011.
Riding, R. and Awramik, S. M. (Eds.): Microbial Sediments, Springer Verlag, Heidelberg, 331 pp., https://doi.org/10.1007/978-3-662-04036-2, 2000.
Riding, R. and Liang, L.: Geobiology of microbial carbonates: Metazoan and seawater saturation state influences on secular trends during the Phanerozoic, Geobiology: Objectives, Concepts, Perspectives, 101–115, https://doi.org/10.1016/B978-0-444-52019-7.50010-3, 2005.
Riding, R., Liang, L., and Braga, J. C.: Millennial-scale ocean acidification and late Quaternary decline of cryptic bacterial crusts in tropical reefs, Geobiol., 12, 387–405, 2014.
Robles-Fernández, A., Areias, C., Daffonchio, D., Vahrenkamp, V. C., and Sánchez-Román, M.: The Role of Microorganisms in the Nucleation of Carbonates, Environmental Implications and Applications, Mineral.-Basel, 12, 1562, https://doi.org/10.3390/min12121562, 2022.
Salas-Saavedra, M., Dechnik, B., Webb, G. E., Webster, J. M., Zhao, J.-X., Nothdurft, L. D., Clark, T. R., Graham, T., and Duce, S.: Holocene reef growth over irregular Pleistocene karst confirms major influence of hydrodynamic factors on Holocene reef development, Quaternary Sci. Rev., 180, 157–176, https://doi.org/10.1016/j.chemgeo.2022.120871, 2018.
Sanborn, K. L., Webster, J. M., Yokoyama, Y., Dutton, A., Braga, J. C., Clague, D. A., Paduan, J. B., Wagner, D., Rooney, J. J., and Hansen, J. R.: New evidence of Hawaiian coral reef drowning in response to meltwater pulse-1A, Quaternary Sci. Rev., 175, 60–72, https://doi.org/10.1016/j.quascirev.2017.08.022, 2017.
Sanfilippo, K., Timm, O. E., Frazier, A. G., and Giambelluca, T. W.: Effects of systematic predictor selection for statistical downscaling of rainfall in Hawai'i, Int. J. Climatol., 44, 571–591, https://doi.org/10.1002/joc.8345, 2024.
Schubert, J. K. and Bottjer, D. J.: Early Triassic stromatolites as post-mass extinction disaster forms, Geology, 20, 883–886, 1992.
Séard, C., Camoin, G., Yokoyama, Y., Matsuzaki, H., Durand, N., Bard, E., Sepulcre, S., and Deschamps, P.: Microbialite development patterns in the last deglacial reefs from Tahiti (French Polynesia; IODP Expedition #310): Implications on reef framework architecture, Mar. Geol., 279, 63–86, https://doi.org/10.1016/j.margeo.2010.10.013, 2011.
Shirokova, L. S., Mavromatis, V., Bundeleva, I. A., Pokrovsky, O. S., Bénézeth, P., Gérard, E., Pearce, C. R., and Oelkers, E. H.: Using Mg Isotopes to Trace Cyanobacterially Mediated Magnesium Carbonate Precipitation in Alkaline Lakes, Aquat. Geochem., 19, 1–24, https://doi.org/10.1007/s10498-012-9174-3, 2013.
Stolz, J., Reid, R. P., Visscher, P., Decho, A., Norman, R., Aspden, R., Bowlin, E., Franks, J., Foster, J., Paterson, D., Przekop, K., Underwood, G., and Prufert-Bebout, L.: The Microbial Communities of the Modern Marine Stromatolites at Highborne Cay, Bahamas, Atoll Research Bulletin, 567, https://doi.org/10.5479/si.00775630.567.1, 2009.
Szilagyi, Z., Webster, J. M., Patterson, M. A., Hips, K., Riding, R., Foley, M., Humblet, M., Yokoyama, Y., Liang, L., Gischler, E., Montaggioni, L., Gherardi, D., and Braga, J. C.: Controls on the spatio-temporal distribution of microbialite crusts on the Great Barrier Reef over the past 30 000 years, Mar. Geol., https://doi.org/10.1016/j.margeo.2020.106312, 2020.
Taylor, B.: Shoreline slope breaks revise understanding of Hawaiian shield volcanoes evolution, Geochem. Geophy. Geosy., 20, 4025–4045, https://doi.org/10.1029/2019GC008436, 2019.
van Lith, Y., Warthmann, R., Vasconselos, C., and McKenzie, J. A.: Sulphate-reducing bacteria induce low-temperature Ca-dolomite and high Mg-calcite formation, Geobiology, 1, 71–79, 2003.
Visscher, P. T. and Stolz, J. F.: Microbial mats as bioreactors: populations, processes, and products, Palaeogeogr. Palaeocl., 219, 87–100, 2005.
Vogt, C., Lauterjung, J., and Fischer, R. X.: Investigation of the clay fraction (<2 µm) of the clay mineral society reference clays, Clay. Clay Miner., 50, 388–400, 2002.
Watts, A. B.: An analysis of isostasy in the world's oceans, 1. Hawaiian-Emperor Seamount Chain, J. Geophys. Res.-Sol. Ea., 83, 5989–6004, https://doi.org/10.1029/JB083iB12p05989, 1978.
Webb, G. E.: Was Phanerozoic reef history controlled by the distribution of nonenzymatically secreted reef carbonates (microbial carbonate and biologically induced cement)?, Sedimentology, 43, 947–971, 1996.
Webb, G. E.: Quantitative Analysis and Paleoecology of Earliest Mississippian Microbial Reefs, Gudman Formation, Queensland, Australia: Not Just Post-Disaster Phenomena, J. Sediment. Res., 75, 875–894, https://doi.org/10.2110/jsr.2005.068, 2005.
Webb, G. E. and Jell, J. S.: Cryptic microbialite in subtidal reef framework and intertidal solution cavities in beachrock, Heron Reef, Great Barrier Reef, Australia: preliminary observations, in: Biosedimentology of microbial buildups, edited by: Neuweiler, F., Reitner, J., and Monty, C., IGCP Project No. 380, Proceedings of 2nd Meeting Göttingen/Germany 1996, Facies, 36, 219–223, 1997.
Webb, G. E. and Kamber, B. S.: Trace Element Geochemistry as a Tool for Interpreting Microbialites, in: Earliest Life on Earth: Habitats, Environments and Methods of Detection, edited by: Golding, S. and Glikson, M., Springer, Dordrecht, https://doi.org/10.1007/978-90-481-8794-2_6, 2010.
Webb, G. E., Baker, J. C., and Jell, J. S.: Inferred syngenetic textural evolution in Holocene cryptic reefal microbialites, Heron Reef, Great Barrier Reef, Australia, Geology, 26, 355–358, 1998.
Webster, J. M., Braga, J. C., Clague, D. A., Gallup, C., Hein, J. R., Potts, D. C., Renema, W., Riding, R., Riker-Coleman, K., Silver, E., and Wallace, L. M.: Coral reef evolution on rapidly subsiding margins, Global Planet. Change, 66, 129–148, https://doi.org/10.1016/j.gloplacha.2008.07.010, 2009.
Webster, J. M., Ravelo, A. C., and Grant, H. L. J.: Expedition 389 Scientific Prospectus: Hawaiian Drowned Reefs, International Ocean Discovery Program, https://doi.org/10.14379/iodp.sp.389.2023, 2023.
Webster, J. M., Ravelo, A. C., Grant, H. L. J., and the Expedition 389 Scientists: Expedition 389 Preliminary Report: Hawaiian Drowned Reefs, International Ocean Discovery Program, https://doi.org/10.14379/iodp.pr.389.2024, 2024.
Webster, J. M., Ravelo, A. C., Grant, H. L. J., and the Expedition 389 Scientists: Hawaiian Drowned Reefs, Proceedings of the International Ocean Discovery Program, 389, International Ocean Discovery Program, College Station, TX, https://doi.org/10.14379/iodp.proc.389.2025, 2025.
Westphal, H., Heindel, K., Brandano, M., and Peckmann, J.: Genesis of microbialites as contemporaneous framework components of coral reefs, deglacial of Tahiti (IODP 310), Facies, 56, 337–352, https://doi.org/10.1007/s10347-009-0207-3, 2010.
Westphal, H., Garuglieri, E., Webb, G. E., Nothdurft, L., Merkel, A., Khanna, P., Karki, P., Nohl, T., Gischler, E., and Webster, J. M.: Scanning Electron Micrographs of biosignatures of microbial mats (IODP Expedition 389), PANGAEA [data set], https://doi.org/10.1594/PANGAEA.995587, 2026.
White, B., Kurkjy, K. A., Curran, H. A., and Besom, K. A.: Shallowing-upward sequence in a Pleistocene coral reef and associated facies, San Salvador, Bahamas, AAPG Bull., 68, 539–539, 1984.
Zhang, H. S., Dai, M.-Y., Qi, Y.-A., Han, L.-L., Yin, Z.-L., Chen, S.-H., and Lin, L.-B.: Girvanella fossils from the Phanerozoic: Distribution, evolution and controlling factors, J. Palaeogeogr., 13, 924–938, 2024.
Zhao, Y., Wei, X., Gao, X., Li, J., Zhang, Y., Hu, K., Han, C., Wang, Q., and Han, Z.: Proto-dolomite spherulites with heterogeneous interior precipitated in brackish water cultivation of freshwater cyanobacterium Leptolyngbya boryana, Sci. Total Environ., 906, 167552, https://doi.org/10.1016/j.scitotenv.2023.167552, 2024.
Zhu, Y. and Dittrich, M.: Carbonate precipitation through microbial activities in natural environments, and their potential in biotechnology: a review, Front. Bioeng. Biotechnol., 4, 4, https://doi.org/10.3389/fbioe.2016.00004, 2016.