Articles | Volume 17, issue 14
https://doi.org/10.5194/bg-17-3815-2020
© Author(s) 2020. This work is distributed under
the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
https://doi.org/10.5194/bg-17-3815-2020
© Author(s) 2020. This work is distributed under
the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
Research article
| 
23 Jul 2020
Research article |
| 23 Jul 2020
| 23 Jul 2020
The Southern Annular Mode (SAM) influences phytoplankton communities in the seasonal ice zone of the Southern Ocean
Bruce L. Greaves
CORRESPONDING AUTHOR
Institute for Marine and Antarctic Studies, University of Tasmania, Private Bag 129, Hobart, Tasmania 7001, Australia
Andrew T. Davidson
Australian Antarctic Division, Department of the Environment and Energy, 203 Channel Highway, Kingston, Tasmania 7050, Australia
Antarctic Climate & Ecosystems Cooperative Research Centre (ACE CRC), University of Tasmania, Private Bag 80, Hobart, Tasmania 7001, Australia
Alexander D. Fraser
Antarctic Climate & Ecosystems Cooperative Research Centre (ACE CRC), University of Tasmania, Private Bag 80, Hobart, Tasmania 7001, Australia
Institute for Marine and Antarctic Studies, University of Tasmania, Private Bag 129, Hobart, Tasmania 7001, Australia
John P. McKinlay
Australian Antarctic Division, Department of the Environment and Energy, 203 Channel Highway, Kingston, Tasmania 7050, Australia
Andrew Martin
Institute for Marine and Antarctic Studies, University of Tasmania, Private Bag 129, Hobart, Tasmania 7001, Australia
Andrew McMinn
Institute for Marine and Antarctic Studies, University of Tasmania, Private Bag 129, Hobart, Tasmania 7001, Australia
Simon W. Wright
Institute for Marine and Antarctic Studies, University of Tasmania, Private Bag 129, Hobart, Tasmania 7001, Australia
Australian Antarctic Division, Department of the Environment and Energy, 203 Channel Highway, Kingston, Tasmania 7050, Australia
Antarctic Climate & Ecosystems Cooperative Research Centre (ACE CRC), University of Tasmania, Private Bag 80, Hobart, Tasmania 7001, Australia
Related authors
No articles found.
Robert Massom, Phillip Reid, Stephen Warren, Bonnie Light, Donald Perovich, Luke Bennetts, Petteri Uotila, Siobhan O'Farrell, Michael Meylan, Klaus Meiners, Pat Wongpan, Alexander Fraser, Alessandro Toffoli, Giulio Passerotti, Peter Strutton, Sean Chua, and Melissa Fedrigo
EGUsphere, https://doi.org/10.5194/egusphere-2025-3166, https://doi.org/10.5194/egusphere-2025-3166, 2025
This preprint is open for discussion and under review for The Cryosphere (TC).
Short summary
Short summary
Ocean waves play a previously-neglected role in the rapid annual melting of Antarctic sea ice by flooding and pulverising floes, removing the snow cover and reducing the albedo by an estimated 0.38–0.54 – to increase solar absorption and enhance the vertical melt rate by up to 5.2 cm/day. Ice algae further decrease the albedo, to increase the melt-rate enhancement to up to 6.1 cm/day. Melting is accelerated by four previously-unconsidered wave-driven positive feedbacks.
Joey J. Voermans, Alexander D. Fraser, Jill Brouwer, Michael H. Meylan, Qingxiang Liu, and Alexander V. Babanin
EGUsphere, https://doi.org/10.5194/egusphere-2024-2104, https://doi.org/10.5194/egusphere-2024-2104, 2024
Short summary
Short summary
Limited measurements of waves in sea ice exist, preventing our understanding of wave attenuation in sea ice under a wide range of ice conditions. Using satellite observations from ICESat-2 we observe an overall linear increase of the wave attenuation rate with distance into the marginal ice zone. While attenuation may vary greatly locally, this finding may provide opportunities for the modelling of waves in sea ice at global and climate scales when such fine detail may not be needed.
Kazuya Kusahara, Daisuke Hirano, Masakazu Fujii, Alexander D. Fraser, Takeshi Tamura, Kohei Mizobata, Guy D. Williams, and Shigeru Aoki
The Cryosphere, 18, 43–73, https://doi.org/10.5194/tc-18-43-2024, https://doi.org/10.5194/tc-18-43-2024, 2024
Short summary
Short summary
This study focuses on the Totten and Moscow University ice shelves, East Antarctica. We used an ocean–sea ice–ice shelf model to better understand regional interactions between ocean, sea ice, and ice shelf. We found that a combination of warm ocean water and local sea ice production influences the regional ice shelf basal melting. Furthermore, the model reproduced the summertime undercurrent on the upper continental slope, regulating ocean heat transport onto the continental shelf.
Lingwei Zhang, Tessa R. Vance, Alexander D. Fraser, Lenneke M. Jong, Sarah S. Thompson, Alison S. Criscitiello, and Nerilie J. Abram
The Cryosphere, 17, 5155–5173, https://doi.org/10.5194/tc-17-5155-2023, https://doi.org/10.5194/tc-17-5155-2023, 2023
Short summary
Short summary
Physical features in ice cores provide unique records of past variability. We identified 1–2 mm ice layers without bubbles in surface ice cores from Law Dome, East Antarctica, occurring on average five times per year. The origin of these bubble-free layers is unknown. In this study, we investigate whether they have the potential to record past atmospheric processes and circulation. We find that the bubble-free layers are linked to accumulation hiatus events and meridional moisture transport.
Lea Pesjak, Andrew McMinn, Zanna Chase, and Helen Bostock
Clim. Past, 19, 419–437, https://doi.org/10.5194/cp-19-419-2023, https://doi.org/10.5194/cp-19-419-2023, 2023
Short summary
Short summary
This study uses diatom assemblages, biogenic silica and Si/Al data over the last 140 kyr from core TAN1302-44 (64°54' S, 144°32' E) to define glacial-to-interglacial paleoenvironments near Antarctica with respect to sea ice duration and ocean circulation. It has found that the sea ice season increased gradually during the last glacial, reaching a maximum before decreasing at the end of MIS 2. Following this, Circumpolar Deep Water increased relative to other times prior to ice sheet retreat.
Jill Brouwer, Alexander D. Fraser, Damian J. Murphy, Pat Wongpan, Alberto Alberello, Alison Kohout, Christopher Horvat, Simon Wotherspoon, Robert A. Massom, Jessica Cartwright, and Guy D. Williams
The Cryosphere, 16, 2325–2353, https://doi.org/10.5194/tc-16-2325-2022, https://doi.org/10.5194/tc-16-2325-2022, 2022
Short summary
Short summary
The marginal ice zone is the region where ocean waves interact with sea ice. Although this important region influences many sea ice, ocean and biological processes, it has been difficult to accurately measure on a large scale from satellite instruments. We present new techniques for measuring wave attenuation using the NASA ICESat-2 laser altimeter. By measuring how waves attenuate within the sea ice, we show that the marginal ice zone may be far wider than previously realised.
Tian R. Tian, Alexander D. Fraser, Noriaki Kimura, Chen Zhao, and Petra Heil
The Cryosphere, 16, 1299–1314, https://doi.org/10.5194/tc-16-1299-2022, https://doi.org/10.5194/tc-16-1299-2022, 2022
Short summary
Short summary
This study presents a comprehensive validation of a satellite observational sea ice motion product in Antarctica by using drifting buoys. Two problems existing in this sea ice motion product have been noticed. After rectifying problems, we use it to investigate the impacts of satellite observational configuration and timescale on Antarctic sea ice kinematics and suggest the future improvement of satellite missions specifically designed for retrieval of sea ice motion.
Jessica Cartwright, Alexander D. Fraser, and Richard Porter-Smith
Earth Syst. Sci. Data, 14, 479–490, https://doi.org/10.5194/essd-14-479-2022, https://doi.org/10.5194/essd-14-479-2022, 2022
Short summary
Short summary
Due to the scale and remote nature of the polar regions, it is essential to use satellite remote sensing to monitor and understand them and their dynamics. Here we present data from the Advanced Scatterometer (ASCAT), processed in a manner proven for use in cryosphere studies. The data have been processed on three timescales (5 d, 2 d and 1 d) in order to optimise temporal resolution as each of the three MetOp satellites is launched.
Alexander D. Fraser, Robert A. Massom, Mark S. Handcock, Phillip Reid, Kay I. Ohshima, Marilyn N. Raphael, Jessica Cartwright, Andrew R. Klekociuk, Zhaohui Wang, and Richard Porter-Smith
The Cryosphere, 15, 5061–5077, https://doi.org/10.5194/tc-15-5061-2021, https://doi.org/10.5194/tc-15-5061-2021, 2021
Short summary
Short summary
Landfast ice is sea ice that remains stationary by attaching to Antarctica's coastline and grounded icebergs. Although a variable feature, landfast ice exerts influence on key coastal processes involving pack ice, the ice sheet, ocean, and atmosphere and is of ecological importance. We present a first analysis of change in landfast ice over an 18-year period and quantify trends (−0.19 ± 0.18 % yr−1). This analysis forms a reference of landfast-ice extent and variability for use in other studies.
Sazlina Salleh and Andrew McMinn
Biogeosciences, 18, 5313–5326, https://doi.org/10.5194/bg-18-5313-2021, https://doi.org/10.5194/bg-18-5313-2021, 2021
Short summary
Short summary
The benthic diatom communities in Tanjung Rhu, Malaysia, were regularly exposed to high light and temperature variability during the tidal cycle, resulting in low photosynthetic efficiency. We examined the impact of high temperatures on diatoms' photosynthetic capacities, and temperatures beyond 50 °C caused severe photoinhibition. At the same time, those diatoms exposed to temperatures of 40 °C did not show any sign of photoinhibition.
Camilla K. Crockart, Tessa R. Vance, Alexander D. Fraser, Nerilie J. Abram, Alison S. Criscitiello, Mark A. J. Curran, Vincent Favier, Ailie J. E. Gallant, Christoph Kittel, Helle A. Kjær, Andrew R. Klekociuk, Lenneke M. Jong, Andrew D. Moy, Christopher T. Plummer, Paul T. Vallelonga, Jonathan Wille, and Lingwei Zhang
Clim. Past, 17, 1795–1818, https://doi.org/10.5194/cp-17-1795-2021, https://doi.org/10.5194/cp-17-1795-2021, 2021
Short summary
Short summary
We present preliminary analyses of the annual sea salt concentrations and snowfall accumulation in a new East Antarctic ice core, Mount Brown South. We compare this record with an updated Law Dome (Dome Summit South site) ice core record over the period 1975–2016. The Mount Brown South record preserves a stronger and inverse signal for the El Niño–Southern Oscillation (in austral winter and spring) compared to the Law Dome record (in summer).
Richard Porter-Smith, John McKinlay, Alexander D. Fraser, and Robert A. Massom
Earth Syst. Sci. Data, 13, 3103–3114, https://doi.org/10.5194/essd-13-3103-2021, https://doi.org/10.5194/essd-13-3103-2021, 2021
Short summary
Short summary
This study quantifies the characteristic complexity
signaturesaround the Antarctic outer coastal margin, giving a multiscale estimate of the magnitude and direction of undulation or complexity at each point location along the entire coastline. It has numerous applications for both geophysical and biological studies and will contribute to Antarctic research requiring quantitative information about this important interface.
Kazuya Kusahara, Daisuke Hirano, Masakazu Fujii, Alexander D. Fraser, and Takeshi Tamura
The Cryosphere, 15, 1697–1717, https://doi.org/10.5194/tc-15-1697-2021, https://doi.org/10.5194/tc-15-1697-2021, 2021
Short summary
Short summary
We used an ocean–sea ice–ice shelf model with a 2–3 km horizontal resolution to investigate ocean–ice shelf/glacier interactions in Lützow-Holm Bay, East Antarctica. The numerical model reproduced the observed warm water intrusion along the deep trough in the bay. We examined in detail (1) water mass changes between the upper continental slope and shelf regions and (2) the fast-ice role in the ocean conditions and basal melting at the Shirase Glacier tongue.
Alexander D. Fraser, Robert A. Massom, Kay I. Ohshima, Sascha Willmes, Peter J. Kappes, Jessica Cartwright, and Richard Porter-Smith
Earth Syst. Sci. Data, 12, 2987–2999, https://doi.org/10.5194/essd-12-2987-2020, https://doi.org/10.5194/essd-12-2987-2020, 2020
Short summary
Short summary
Landfast ice, or
fast ice, is a form of sea ice which is mechanically fastened to stationary parts of the coast. Long-term and accurate knowledge of its extent around Antarctica is critical for understanding a number of important Antarctic coastal processes, yet no accurate, large-scale, long-term dataset of its extent has been available. We address this data gap with this new dataset compiled from satellite imagery, containing high-resolution maps of Antarctic fast ice from 2000 to 2018.
Stefanie Arndt, Mario Hoppmann, Holger Schmithüsen, Alexander D. Fraser, and Marcel Nicolaus
The Cryosphere, 14, 2775–2793, https://doi.org/10.5194/tc-14-2775-2020, https://doi.org/10.5194/tc-14-2775-2020, 2020
Stacy Deppeler, Kai G. Schulz, Alyce Hancock, Penelope Pascoe, John McKinlay, and Andrew Davidson
Biogeosciences, 17, 4153–4171, https://doi.org/10.5194/bg-17-4153-2020, https://doi.org/10.5194/bg-17-4153-2020, 2020
Short summary
Short summary
Our study showed how ocean acidification can exert both direct and indirect influences on the interactions among trophic levels within the microbial loop. Microbial grazer abundance was reduced at CO2 concentrations at and above 634 µatm, while microbial communities increased in abundance, likely due to a reduction in being grazed. Such changes in predator–prey interactions with ocean acidification could have significant effects on the food web and biogeochemistry in the Southern Ocean.
Alyce M. Hancock, Andrew T. Davidson, John McKinlay, Andrew McMinn, Kai G. Schulz, and Rick L. van den Enden
Biogeosciences, 15, 2393–2410, https://doi.org/10.5194/bg-15-2393-2018, https://doi.org/10.5194/bg-15-2393-2018, 2018
Short summary
Short summary
Absorption of carbon dioxide (CO2) realized by humans is decreasing the ocean pH (ocean acidification). Single-celled organisms (microbes) support the Antarctic ecosystem, yet little is known about their sensitivity to ocean acidification. This study shows a shift in a natural Antarctic microbial community, with CO2 levels exceeding 634 μatm changing the community composition and favouring small cells. This would have significant flow effects for Antarctic food webs and elemental cycles.
Stacy Deppeler, Katherina Petrou, Kai G. Schulz, Karen Westwood, Imojen Pearce, John McKinlay, and Andrew Davidson
Biogeosciences, 15, 209–231, https://doi.org/10.5194/bg-15-209-2018, https://doi.org/10.5194/bg-15-209-2018, 2018
Short summary
Short summary
We combined productivity and photophysiology measurements to investigate the effects of ocean acidification on a natural Antarctic marine microbial community. Our study identifies a threshold for CO2 tolerance in the phytoplankton community between 953 and 1140 μatm of CO2, above which productivity declines. Bacteria were tolerant to CO2 up to 1641 μatm. We identify physiological changes in the phytoplankton at high CO2 that allowed them to acclimate to the high CO2 treatment.
Andrew McMinn
Biogeosciences, 14, 3927–3935, https://doi.org/10.5194/bg-14-3927-2017, https://doi.org/10.5194/bg-14-3927-2017, 2017
Short summary
Short summary
Dissolved carbon dioxide levels in the oceans are rising and this is causing a drop in the pH (ocean acidification). This potentially effects all marine organisms, including those in polar regions. Sea ice algae are naturally exposed to a wide range of pH and CO2 concentrations, particularly during the ice formation and melting cycles. However, all studies so far have shown ice algae to be quite resilient to change. This includes the effects of co-stressors such as light, iron and temperature.
Mana Inoue, Mark A. J. Curran, Andrew D. Moy, Tas D. van Ommen, Alexander D. Fraser, Helen E. Phillips, and Ian D. Goodwin
Clim. Past, 13, 437–453, https://doi.org/10.5194/cp-13-437-2017, https://doi.org/10.5194/cp-13-437-2017, 2017
Short summary
Short summary
A 120 m ice core from Mill Island, East Antarctica, was studied its chemical components. The Mill Island ice core contains 97 years of climate record (1913–2009) and has a mean snow accumulation of 1.35 m yr−1 (ice equivalent). Trace ion concentrations were generally higher than other Antarctic ice core sites. Nearby sea ice concentration was found to influence the annual mean sea salt record. The Mill Island ice core records are unexpectedly complex, with strong modulation of the trace chemistry.
J. L. Lieser, M. A. J. Curran, A. R. Bowie, A. T. Davidson, S. J. Doust, A. D. Fraser, B. K. Galton-Fenzi, R. A. Massom, K. M. Meiners, J. Melbourne-Thomas, P. A. Reid, P. G. Strutton, T. R. Vance, M. Vancoppenolle, K. J. Westwood, and S. W. Wright
The Cryosphere Discuss., https://doi.org/10.5194/tcd-9-6187-2015, https://doi.org/10.5194/tcd-9-6187-2015, 2015
Revised manuscript has not been submitted
Related subject area
Biogeochemistry: Open Ocean
 Coccolithophore abundance and production and their impacts on particulate inorganic carbon cycling in the western North Pacific
Coccolithophore abundance and production and their impacts on particulate inorganic carbon cycling in the western North Pacific
 A time series analysis of transparent exopolymer particle distributions and C : N stoichiometry in the subtropical North Pacific: a key process in net community production and preformed nitrate anomalies?
A time series analysis of transparent exopolymer particle distributions and C : N stoichiometry in the subtropical North Pacific: a key process in net community production and preformed nitrate anomalies?
 Marine snow morphology drives sinking and attenuation in the ocean interior
Marine snow morphology drives sinking and attenuation in the ocean interior
 An upper-mesopelagic-zone carbon budget for the subarctic North Pacific
An upper-mesopelagic-zone carbon budget for the subarctic North Pacific
 Ocean alkalinity enhancement in an open-ocean ecosystem: biogeochemical responses and carbon storage durability
Ocean alkalinity enhancement in an open-ocean ecosystem: biogeochemical responses and carbon storage durability
 Relationships between the concentration of particulate organic nitrogen and the inherent optical properties of seawater in oceanic surface waters
Relationships between the concentration of particulate organic nitrogen and the inherent optical properties of seawater in oceanic surface waters
 Inadequacies in the representation of sub-seasonal phytoplankton dynamics in Earth system models
Nitrogen dynamics and nitrate stable isotopes indicate nitrogen loss in the Bay of Bengal
Interactions between ocean alkalinity enhancement and phytoplankton in an Earth System Model
A tracer study for the development of in-water monitoring, reporting, and verification (MRV) of ship-based ocean alkalinity enhancement
Inadequacies in the representation of sub-seasonal phytoplankton dynamics in Earth system models
Nitrogen dynamics and nitrate stable isotopes indicate nitrogen loss in the Bay of Bengal
Interactions between ocean alkalinity enhancement and phytoplankton in an Earth System Model
A tracer study for the development of in-water monitoring, reporting, and verification (MRV) of ship-based ocean alkalinity enhancement
 Composite model-based estimate of the ocean carbon sink from 1959 to 2022
Composite model-based estimate of the ocean carbon sink from 1959 to 2022
 Phytoplankton community structure in relation to iron and macronutrient fluxes from subsurface waters in the western North Pacific during summer
Phytoplankton community structure in relation to iron and macronutrient fluxes from subsurface waters in the western North Pacific during summer
 Intense and localized export of selected marine snow types at eddy edges in the South Atlantic Ocean
Intense and localized export of selected marine snow types at eddy edges in the South Atlantic Ocean
 Spatial distributions of iron and manganese in surface waters of the Arctic's Laptev and East Siberian seas
Spatial distributions of iron and manganese in surface waters of the Arctic's Laptev and East Siberian seas
 Climate-driven shifts in Southern Ocean primary producers and biogeochemistry in CMIP6 models
Oceanic enrichment of ammonium and its impacts on phytoplankton community composition under a high-emissions scenario
Climate-driven shifts in Southern Ocean primary producers and biogeochemistry in CMIP6 models
Oceanic enrichment of ammonium and its impacts on phytoplankton community composition under a high-emissions scenario
 Ocean acidification trends and carbonate system dynamics across the North Atlantic subpolar gyre water masses during 2009–2019
From small scale variability to mesoscale stability in surface ocean pH: implications for air-sea CO2 equilibration
Ocean acidification trends and carbonate system dynamics across the North Atlantic subpolar gyre water masses during 2009–2019
From small scale variability to mesoscale stability in surface ocean pH: implications for air-sea CO2 equilibration
 Sedimentary organic matter signature hints at the phytoplankton-driven biological carbon pump in the central Arabian Sea
Sedimentary organic matter signature hints at the phytoplankton-driven biological carbon pump in the central Arabian Sea
 Hydrological cycle amplification imposes spatial patterns on the climate change response of ocean pH and carbonate chemistry
Hydrological cycle amplification imposes spatial patterns on the climate change response of ocean pH and carbonate chemistry
 Assessing the tropical Atlantic biogeochemical processes in the Norwegian Earth System Model
Assessing the tropical Atlantic biogeochemical processes in the Norwegian Earth System Model
 Evolution of oxygen and stratification and their relationship in the North Pacific Ocean in CMIP6 Earth system models
Evolution of oxygen and stratification and their relationship in the North Pacific Ocean in CMIP6 Earth system models
 Evaluation of CMIP6 model performance in simulating historical biogeochemistry across the southern South China Sea
Evaluation of CMIP6 model performance in simulating historical biogeochemistry across the southern South China Sea
 Drivers of decadal trends in the ocean carbon sink in the past, present, and future in Earth system models
Drivers of decadal trends in the ocean carbon sink in the past, present, and future in Earth system models
 Anthropogenic carbon storage and its decadal changes in the Atlantic between 1990–2020
Anthropogenic carbon storage and its decadal changes in the Atlantic between 1990–2020
 Ocean alkalinity enhancement impacts: regrowth of marine microalgae in alkaline mineral concentrations simulating the initial concentrations after ship-based dispersions
Ocean alkalinity enhancement impacts: regrowth of marine microalgae in alkaline mineral concentrations simulating the initial concentrations after ship-based dispersions
 Climatic controls on metabolic constraints in the ocean
Climatic controls on metabolic constraints in the ocean
 Effects of grain size and seawater salinity on magnesium hydroxide dissolution and secondary calcium carbonate precipitation kinetics: implications for ocean alkalinity enhancement
Effects of grain size and seawater salinity on magnesium hydroxide dissolution and secondary calcium carbonate precipitation kinetics: implications for ocean alkalinity enhancement
 Short-term response of Emiliania huxleyi growth and morphology to abrupt salinity stress
Short-term response of Emiliania huxleyi growth and morphology to abrupt salinity stress
 Assessing the impact of CO2-equilibrated ocean alkalinity enhancement on microbial metabolic rates in an oligotrophic system
Assessing the impact of CO2-equilibrated ocean alkalinity enhancement on microbial metabolic rates in an oligotrophic system
 Phosphomonoesterase and phosphodiesterase activities in the eastern Mediterranean in two contrasting seasonal situations
Phosphomonoesterase and phosphodiesterase activities in the eastern Mediterranean in two contrasting seasonal situations
 Net primary production annual maxima in the North Atlantic projected to shift in the 21st century
Net primary production annual maxima in the North Atlantic projected to shift in the 21st century
 Testing the influence of light on nitrite cycling in the eastern tropical North Pacific
Testing the influence of light on nitrite cycling in the eastern tropical North Pacific
 Loss of nitrogen via anaerobic ammonium oxidation (anammox) in the California Current system during the late Quaternary
Loss of nitrogen via anaerobic ammonium oxidation (anammox) in the California Current system during the late Quaternary
 Technical note: Assessment of float pH data quality control methods – a case study in the subpolar northwest Atlantic Ocean
Technical note: Assessment of float pH data quality control methods – a case study in the subpolar northwest Atlantic Ocean
 Linking northeastern North Pacific oxygen changes to upstream surface outcrop variations
Linking northeastern North Pacific oxygen changes to upstream surface outcrop variations
 Underestimation of multi-decadal global O2 loss due to an optimal interpolation method
Underestimation of multi-decadal global O2 loss due to an optimal interpolation method
 Reviews and syntheses: expanding the global coverage of gross primary production and net community production measurements using Biogeochemical-Argo floats
Reviews and syntheses: expanding the global coverage of gross primary production and net community production measurements using Biogeochemical-Argo floats
 Characteristics of surface physical and biogeochemical parameters within mesoscale eddies in the Southern Ocean
Characteristics of surface physical and biogeochemical parameters within mesoscale eddies in the Southern Ocean
 Seasonal dynamics and annual budget of dissolved inorganic carbon in the northwestern Mediterranean deep-convection region
Seasonal dynamics and annual budget of dissolved inorganic carbon in the northwestern Mediterranean deep-convection region
 The fingerprint of climate variability on the surface ocean cycling of iron and its isotopes
The fingerprint of climate variability on the surface ocean cycling of iron and its isotopes
 Reconstructing the ocean's mesopelagic zone carbon budget: sensitivity and estimation of parameters associated with prokaryotic remineralization
Reconstructing the ocean's mesopelagic zone carbon budget: sensitivity and estimation of parameters associated with prokaryotic remineralization
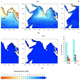
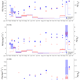 Seasonal cycles of biogeochemical fluxes in the Scotia Sea, Southern Ocean: a stable isotope approach
Seasonal cycles of biogeochemical fluxes in the Scotia Sea, Southern Ocean: a stable isotope approach
 Absence of photophysiological response to iron addition in autumn phytoplankton in the Antarctic sea-ice zone
Absence of photophysiological response to iron addition in autumn phytoplankton in the Antarctic sea-ice zone
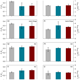
 Optimal parameters for the ocean's nutrient, carbon, and oxygen cycles compensate for circulation biases but replumb the biological pump
Optimal parameters for the ocean's nutrient, carbon, and oxygen cycles compensate for circulation biases but replumb the biological pump
 Importance of multiple sources of iron for the upper-ocean biogeochemistry over the northern Indian Ocean
Importance of multiple sources of iron for the upper-ocean biogeochemistry over the northern Indian Ocean
 Exploring the role of different data types and timescales in the quality of marine biogeochemical model calibration
Exploring the role of different data types and timescales in the quality of marine biogeochemical model calibration
 All about nitrite: exploring nitrite sources and sinks in the eastern tropical North Pacific oxygen minimum zone
All about nitrite: exploring nitrite sources and sinks in the eastern tropical North Pacific oxygen minimum zone
 Fossil coccolith morphological attributes as a new proxy for deep ocean carbonate chemistry
Fossil coccolith morphological attributes as a new proxy for deep ocean carbonate chemistry
 Reconstructing ocean carbon storage with CMIP6 Earth system models and synthetic Argo observations
Reconstructing ocean carbon storage with CMIP6 Earth system models and synthetic Argo observations
Yuye Han, Zvi Steiner, Zhimian Cao, Di Fan, Junhui Chen, Jimin Yu, and Minhan Dai
Biogeosciences, 22, 3681–3697, https://doi.org/10.5194/bg-22-3681-2025, https://doi.org/10.5194/bg-22-3681-2025, 2025
Short summary
Short summary
Our results suggest coccolithophore calcite accounts for a major fraction of PIC (particulate inorganic carbon) standing stocks in the western North Pacific, with a markedly higher contribution in the oligotrophic subtropical gyre than in the Kuroshio–Oyashio transition region, which highlights the importance of coccolithophores for PIC production in the pelagic ocean, particularly in oligotrophic ocean waters.
Kieran Curran, Tracy A. Villareal, and Robert T. Letscher
Biogeosciences, 22, 3515–3531, https://doi.org/10.5194/bg-22-3515-2025, https://doi.org/10.5194/bg-22-3515-2025, 2025
Short summary
Short summary
This work provides a 2-year record of marine organic gel concentrations from an open-ocean site in the subtropical North Pacific Ocean north of Hawaii. These microscopic gels are investigated to understand their importance as an understudied component of organic matter cycling by marine microbes. We find an important role of gel cycling during the summer months, helping explain previously contradictory estimates of nutrient supply and demand for the subtropical ocean.
Yawouvi Dodji Soviadan, Miriam Beck, Joelle Habib, Alberto Baudena, Laetitia Drago, Alexandre Accardo, Remi Laxenaire, Sabrina Speich, Peter Brandt, Rainer Kiko, and Stemmann Lars
Biogeosciences, 22, 3485–3501, https://doi.org/10.5194/bg-22-3485-2025, https://doi.org/10.5194/bg-22-3485-2025, 2025
Short summary
Short summary
Key parameters representing the gravity flux in global models are sinking speed and vertical attenuation of exported material. We calculate, for the first time, these parameters in situ in the ocean for six intermittent blooms followed by export events using high-resolution (3 d) time series of 0–1000 m depth profiles from imaging sensors mounted on an Argo float. We show that sinking speed depends not only on size but also on the morphology of the particles, with density being an important property.
Brandon M. Stephens, Montserrat Roca-Martí, Amy E. Maas, Vinícius J. Amaral, Samantha Clevenger, Shawnee Traylor, Claudia R. Benitez-Nelson, Philip W. Boyd, Ken O. Buesseler, Craig A. Carlson, Nicolas Cassar, Margaret Estapa, Andrea J. Fassbender, Yibin Huang, Phoebe J. Lam, Olivier Marchal, Susanne Menden-Deuer, Nicola L. Paul, Alyson E. Santoro, David A. Siegel, and David P. Nicholson
Biogeosciences, 22, 3301–3328, https://doi.org/10.5194/bg-22-3301-2025, https://doi.org/10.5194/bg-22-3301-2025, 2025
Short summary
Short summary
The ocean’s mesopelagic zone (MZ) plays a crucial role in the global carbon cycle. This study combines new and previously published measurements of organic carbon supply and demand collected in August 2018 in the MZ of the subarctic North Pacific Ocean. Supply was insufficient to meet demand in August, but supply entering into the MZ in the spring of 2018 could have met the August demand. Results suggest observations over seasonal timescales may help to close MZ carbon budgets.
Allanah Joy Paul, Mathias Haunost, Silvan Urs Goldenberg, Jens Hartmann, Nicolás Sánchez, Julieta Schneider, Niels Suitner, and Ulf Riebesell
Biogeosciences, 22, 2749–2766, https://doi.org/10.5194/bg-22-2749-2025, https://doi.org/10.5194/bg-22-2749-2025, 2025
Short summary
Short summary
Ocean alkalinity enhancement (OAE) is being assessed for its potential to absorb atmospheric CO2 and store it for a long time. OAE still needs comprehensive assessment of its safety and effectiveness. We studied an idealised OAE application in a natural low-nutrient ecosystem over 1 month. Our results showed that biogeochemical functioning remained mostly stable but that the long-term capability for storing carbon may be limited at high alkalinity concentration.
Alain Fumenia, Hubert Loisel, Rick A. Reynolds, and Dariusz Stramski
Biogeosciences, 22, 2461–2484, https://doi.org/10.5194/bg-22-2461-2025, https://doi.org/10.5194/bg-22-2461-2025, 2025
Short summary
Short summary
Particulate organic nitrogen (PON) plays a central role in ocean biogeochemistry, yet limited in situ data hinder a full understanding of PON variability and associated processes. Measurements of optical properties offer an alternative for assessing PON across diverse marine environments. Our analysis reveals strong relationships between PON and optical properties, supporting a promising means to assess PON from optical measurements performed in situ or conducted from remote-sensing platforms.
Madhavan Girijakumari Keerthi, Olivier Aumont, Lester Kwiatkowski, and Marina Levy
Biogeosciences, 22, 2163–2180, https://doi.org/10.5194/bg-22-2163-2025, https://doi.org/10.5194/bg-22-2163-2025, 2025
Short summary
Short summary
We assessed how well climate models replicate sub-seasonal changes in ocean chlorophyll observed by satellites. Models struggle to capture these variations accurately. Some overestimate fluctuations and their impact on annual chlorophyll variability, while others underestimate them. The underestimation is likely due to limited model resolution, while the overestimation may come from internal model oscillations.
Gesa Schulz, Kirstin Dähnke, Tina Sanders, Jan Penopp, Hermann W. Bange, Rena Czeschel, and Birgit Gaye
EGUsphere, https://doi.org/10.5194/egusphere-2025-1660, https://doi.org/10.5194/egusphere-2025-1660, 2025
Short summary
Short summary
Oxygen minimum zones (OMZs) are low-oxygen ocean areas that deplete nitrogen, a key marine nutrient. Understanding nitrogen cycling in OMZs is crucial for the global nitrogen cycle. This study examined nitrogen cycling in the OMZ of the Bay of Bengal and East Equatorial Indian Ocean, revealing limited mixing between both regions. Surface phytoplankton consumes nitrate, while deeper nitrification recycles nitrogen. In the BoB’s OMZ (100–300 m), nitrogen loss likely occurs via anammox.
Miriam Seifert, Christopher Danek, Christoph Völker, and Judith Hauck
EGUsphere, https://doi.org/10.5194/egusphere-2025-1495, https://doi.org/10.5194/egusphere-2025-1495, 2025
Short summary
Short summary
Ocean alkalinity enhancement (OAE) can help to remove CO2 from the atmosphere. Yet, our study is the first that investigates the link between OAE and phytoplankton in an Earth System Model. We show that OAE can indirectly decrease primary production, and that biological feedbacks can modify the efficiency of OAE. Our study provides hints for ecological risks of OAE and the quantification of carbon drawdown, and can serve as a kick-start for other models to consider OAE-ecosystem interactions.
Adam V. Subhas, Jennie E. Rheuban, Zhaohui Aleck Wang, Daniel C. McCorkle, Anna P. M. Michel, Lukas Marx, Chloe L. Dean, Kate Morkeski, Matthew G. Hayden, Mary Burkitt-Gray, Francis Elder, Yiming Guo, Heather H. Kim, and Ke Chen
EGUsphere, https://doi.org/10.5194/egusphere-2025-1348, https://doi.org/10.5194/egusphere-2025-1348, 2025
Short summary
Short summary
Ocean alkalinity enhancement (OAE) is a carbon removal approach in which alkaline materials are added to the marine environment, increasing the ocean's ability to store carbon dioxide. We conducted an open-water experiment releasing and tracking a fluorescent water tracer. Under the right conditions, in-water monitoring of OAE does appear to be possible. We conclude with a series of practical recommendations for open-water OAE monitoring.
Jens Terhaar
Biogeosciences, 22, 1631–1649, https://doi.org/10.5194/bg-22-1631-2025, https://doi.org/10.5194/bg-22-1631-2025, 2025
Short summary
Short summary
The ocean is a major natural carbon sink. Despite its importance, estimates of the ocean carbon sink remain uncertain. Here, I present a hybrid model estimate of the ocean carbon sink from 1959 to 2022. By combining ocean models in hindcast mode and Earth system models, I keep the strength of each approach and remove the respective weaknesses. This composite model estimate is similar in magnitude to the best estimate of the Global Carbon Budget but 70 % less uncertain.
Huailin Deng, Koji Suzuki, Ichiro Yasuda, Hiroshi Ogawa, and Jun Nishioka
Biogeosciences, 22, 1495–1508, https://doi.org/10.5194/bg-22-1495-2025, https://doi.org/10.5194/bg-22-1495-2025, 2025
Short summary
Short summary
Iron (Fe) and nitrate are vital for primary production in the North Pacific. Sedimentary Fe is carried by North Pacific Intermediate Water to the North Pacific, but the nutrient return path and its effect on phytoplankton are unclear. By combining Fe and macronutrient fluxes with phytoplankton composition, this study firstly revealed that Fe supply from the subsurface greatly controls diatom abundance and identified the nutrient return path in the subarctic gyre and Kuroshio–Oyashio transition area.
Alexandre Accardo, Rémi Laxenaire, Alberto Baudena, Sabrina Speich, Rainer Kiko, and Lars Stemmann
Biogeosciences, 22, 1183–1201, https://doi.org/10.5194/bg-22-1183-2025, https://doi.org/10.5194/bg-22-1183-2025, 2025
Short summary
Short summary
The open ocean helps mitigate climate change by storing CO2 via the biological carbon pump (BCP), which involves processes like organic carbon production at the surface and transferring it to the deep ocean via various pathways. By deploying an autonomous platform, we found significant marine snow accumulation from the surface to the mesopelagic zone in frontal regions between eddies. We suggest that the coupling of hydrodynamics at eddy edges and biological activity may enhance this process.
Naoya Kanna, Kazutaka Tateyama, Takuji Waseda, Anna Timofeeva, Maria Papadimitraki, Laura Whitmore, Hajime Obata, Daiki Nomura, Hiroshi Ogawa, Youhei Yamashita, and Igor Polyakov
Biogeosciences, 22, 1057–1076, https://doi.org/10.5194/bg-22-1057-2025, https://doi.org/10.5194/bg-22-1057-2025, 2025
Short summary
Short summary
This article presents data on iron and manganese, essential micronutrients for primary producers in the Arctic Laptev and East Siberian seas (LESS). There, observations were made through international cooperation with the Nansen and Amundsen Basin Observational System expedition during the late summer of 2021. The results from this study indicate that the major sources controlling the iron and manganese distributions on the LESS continental margins are river discharge and shelf sediment input.
Ben J. Fisher, Alex J. Poulton, Michael P. Meredith, Kimberlee Baldry, Oscar Schofield, and Sian F. Henley
Biogeosciences, 22, 975–994, https://doi.org/10.5194/bg-22-975-2025, https://doi.org/10.5194/bg-22-975-2025, 2025
Short summary
Short summary
The Southern Ocean is a rapidly warming environment, with subsequent impacts on ecosystems and biogeochemical cycling. This study examines changes in phytoplankton and biogeochemistry using a range of climate models. Under climate change, the Southern Ocean will be warmer, more acidic and more productive and will have reduced nutrient availability by 2100. However, there is substantial variability between models across key productivity parameters. We propose ways of reducing this uncertainty.
Pearse J. Buchanan, Juan J. Pierella Karlusich, Robyn E. Tuerena, Roxana Shafiee, E. Malcolm S. Woodward, Chris Bowler, and Alessandro Tagliabue
EGUsphere, https://doi.org/10.5194/egusphere-2024-3639, https://doi.org/10.5194/egusphere-2024-3639, 2025
Short summary
Short summary
Ammonium is a form of nitrogen that may become more important for growth of marine primary producers (i.e., phytoplankton) in the future. Because some phytoplankton taxa have a greater affinity for ammonium than others, the relative increase in ammonium could cause shifts in community composition. We quantify ammonium enrichment, identify its drivers, and isolate the possible effect on phytoplankton community composition under a high emissions scenario.
David Curbelo-Hernández, Fiz F. Pérez, Melchor González-Dávila, Sergey V. Gladyshev, Aridane G. González, David González-Santana, Antón Velo, Alexey Sokov, and J. Magdalena Santana-Casiano
Biogeosciences, 21, 5561–5589, https://doi.org/10.5194/bg-21-5561-2024, https://doi.org/10.5194/bg-21-5561-2024, 2024
Short summary
Short summary
The study evaluated CO2–carbonate system dynamics in the North Atlantic subpolar gyre during 2009–2019. Significant ocean acidification, largely due to rising anthropogenic CO2 levels, was found. Cooling, freshening, and enhanced convective processes intensified this trend, affecting calcite and aragonite saturation. The findings contribute to a deeper understanding of ocean acidification and improve our knowledge about its impact on marine ecosystems.
Louise Delaigue, Gert-Jan Reichart, Chris Galley, Yasmina Ourradi, and Matthew Paul Humphreys
EGUsphere, https://doi.org/10.5194/egusphere-2024-2853, https://doi.org/10.5194/egusphere-2024-2853, 2024
Short summary
Short summary
Our study analyzed pH in ocean surface waters to understand how they fluctuate with changes in temperature, salinity, and biological activities. We found that temperature mainly controls daily pH variations, but biological processes also play a role, especially in affecting CO2 levels between the ocean and atmosphere. Our research shows how these factors together maintain the balance of ocean chemistry, which is crucial for predicting changes in marine environments.
Medhavi Pandey, Haimanti Biswas, Daniel Birgel, Nicole Burdanowitz, and Birgit Gaye
Biogeosciences, 21, 4681–4698, https://doi.org/10.5194/bg-21-4681-2024, https://doi.org/10.5194/bg-21-4681-2024, 2024
Short summary
Short summary
We analysed sea surface temperature (SST) proxy and plankton biomarkers in sediments that accumulate sinking material signatures from surface waters in the central Arabian Sea (21°–11° N, 64° E), a tropical basin impacted by monsoons. We saw a north–south SST gradient, and the biological proxies showed more organic matter from larger algae in the north. Smaller algae and zooplankton were more numerous in the south. These trends were related to ocean–atmospheric processes and oxygen availability.
Allison Hogikyan and Laure Resplandy
Biogeosciences, 21, 4621–4636, https://doi.org/10.5194/bg-21-4621-2024, https://doi.org/10.5194/bg-21-4621-2024, 2024
Short summary
Short summary
Rising atmospheric CO2 influences ocean carbon chemistry, leading to ocean acidification. Global warming introduces spatial patterns in the intensity of ocean acidification. We show that the most prominent spatial patterns are controlled by warming-driven changes in rainfall and evaporation, not by the direct effect of warming on carbon chemistry and pH. These evaporation and rainfall patterns oppose acidification in saltier parts of the ocean and enhance acidification in fresher regions.
Shunya Koseki, Lander R. Crespo, Jerry Tjiputra, Filippa Fransner, Noel S. Keenlyside, and David Rivas
Biogeosciences, 21, 4149–4168, https://doi.org/10.5194/bg-21-4149-2024, https://doi.org/10.5194/bg-21-4149-2024, 2024
Short summary
Short summary
We investigated how the physical biases of an Earth system model influence the marine biogeochemical processes in the tropical Atlantic. With four different configurations of the model, we have shown that the versions with better SST reproduction tend to better represent the primary production and air–sea CO2 flux in terms of climatology, seasonal cycle, and response to climate variability.
Lyuba Novi, Annalisa Bracco, Takamitsu Ito, and Yohei Takano
Biogeosciences, 21, 3985–4005, https://doi.org/10.5194/bg-21-3985-2024, https://doi.org/10.5194/bg-21-3985-2024, 2024
Short summary
Short summary
We explored the relationship between oxygen and stratification in the North Pacific Ocean using a combination of data mining and machine learning. We used isopycnic potential vorticity (IPV) as an indicator to quantify ocean ventilation and analyzed its predictability, a strong O2–IPV connection, and predictability for IPV in the tropical Pacific. This opens new routes for monitoring ocean O2 through few observational sites co-located with more abundant IPV measurements in the tropical Pacific.
Winfred Marshal, Jing Xiang Chung, Nur Hidayah Roseli, Roswati Md Amin, and Mohd Fadzil Bin Mohd Akhir
Biogeosciences, 21, 4007–4035, https://doi.org/10.5194/bg-21-4007-2024, https://doi.org/10.5194/bg-21-4007-2024, 2024
Short summary
Short summary
This study stands out for thoroughly examining CMIP6 ESMs' ability to simulate biogeochemical variables in the southern South China Sea, an economically important region. It assesses variables like chlorophyll, phytoplankton, nitrate, and oxygen on annual and seasonal scales. While global assessments exist, this study addresses a gap by objectively ranking 13 CMIP6 ocean biogeochemistry models' performance at a regional level, focusing on replicating specific observed biogeochemical variables.
Jens Terhaar
Biogeosciences, 21, 3903–3926, https://doi.org/10.5194/bg-21-3903-2024, https://doi.org/10.5194/bg-21-3903-2024, 2024
Short summary
Short summary
Despite the ocean’s importance in the carbon cycle and hence the climate, observing the ocean carbon sink remains challenging. Here, I use an ensemble of 12 models to understand drivers of decadal trends of the past, present, and future ocean carbon sink. I show that 80 % of the decadal trends in the multi-model mean ocean carbon sink can be explained by changes in decadal trends in atmospheric CO2. The remaining 20 % are due to internal climate variability and ocean heat uptake.
Reiner Steinfeldt, Monika Rhein, and Dagmar Kieke
Biogeosciences, 21, 3839–3867, https://doi.org/10.5194/bg-21-3839-2024, https://doi.org/10.5194/bg-21-3839-2024, 2024
Short summary
Short summary
We calculate the amount of anthropogenic carbon (Cant) in the Atlantic for the years 1990, 2000, 2010 and 2020. Cant is the carbon that is taken up by the ocean as a result of humanmade CO2 emissions. To determine the amount of Cant, we apply a technique that is based on the observations of other humanmade gases (e.g., chlorofluorocarbons). Regionally, changes in ocean ventilation have an impact on the storage of Cant. Overall, the increase in Cant is driven by the rising CO2 in the atmosphere.
Stephanie Delacroix, Tor Jensen Nystuen, August E. Dessen Tobiesen, Andrew L. King, and Erik Höglund
Biogeosciences, 21, 3677–3690, https://doi.org/10.5194/bg-21-3677-2024, https://doi.org/10.5194/bg-21-3677-2024, 2024
Short summary
Short summary
The addition of alkaline minerals into the ocean might reduce excessive anthropogenic CO2 emissions. Magnesium hydroxide can be added in large amounts because of its low seawater solubility without reaching harmful pH levels. The toxicity effect results of magnesium hydroxide, by simulating the expected concentrations from a ship's dispersion scenario, demonstrated low impacts on both sensitive and local assemblages of marine microalgae when compared to calcium hydroxide.
Precious Mongwe, Matthew Long, Takamitsu Ito, Curtis Deutsch, and Yeray Santana-Falcón
Biogeosciences, 21, 3477–3490, https://doi.org/10.5194/bg-21-3477-2024, https://doi.org/10.5194/bg-21-3477-2024, 2024
Short summary
Short summary
We use a collection of measurements that capture the physiological sensitivity of organisms to temperature and oxygen and a CESM1 large ensemble to investigate how natural climate variations and climate warming will impact the ability of marine heterotrophic marine organisms to support habitats in the future. We find that warming and dissolved oxygen loss over the next several decades will reduce the volume of ocean habitats and will increase organisms' vulnerability to extremes.
Charly A. Moras, Tyler Cyronak, Lennart T. Bach, Renaud Joannes-Boyau, and Kai G. Schulz
Biogeosciences, 21, 3463–3475, https://doi.org/10.5194/bg-21-3463-2024, https://doi.org/10.5194/bg-21-3463-2024, 2024
Short summary
Short summary
We investigate the effects of mineral grain size and seawater salinity on magnesium hydroxide dissolution and calcium carbonate precipitation kinetics for ocean alkalinity enhancement. Salinity did not affect the dissolution, but calcium carbonate formed earlier at lower salinities due to the lower magnesium and dissolved organic carbon concentrations. Smaller grain sizes dissolved faster but calcium carbonate precipitated earlier, suggesting that medium grain sizes are optimal for kinetics.
Rosie M. Sheward, Christina Gebühr, Jörg Bollmann, and Jens O. Herrle
Biogeosciences, 21, 3121–3141, https://doi.org/10.5194/bg-21-3121-2024, https://doi.org/10.5194/bg-21-3121-2024, 2024
Short summary
Short summary
How quickly do marine microorganisms respond to salinity stress? Our experiments with the calcifying marine plankton Emiliania huxleyi show that growth and cell morphology responded to salinity stress within as little as 24–48 hours, demonstrating that morphology and calcification are sensitive to salinity over a range of timescales. Our results have implications for understanding the short-term role of E. huxleyi in biogeochemical cycles and in size-based paleoproxies for salinity.
Laura Marín-Samper, Javier Arístegui, Nauzet Hernández-Hernández, Joaquín Ortiz, Stephen D. Archer, Andrea Ludwig, and Ulf Riebesell
Biogeosciences, 21, 2859–2876, https://doi.org/10.5194/bg-21-2859-2024, https://doi.org/10.5194/bg-21-2859-2024, 2024
Short summary
Short summary
Our planet is facing a climate crisis. Scientists are working on innovative solutions that will aid in capturing the hard to abate emissions before it is too late. Exciting research reveals that ocean alkalinity enhancement, a key climate change mitigation strategy, does not harm phytoplankton, the cornerstone of marine ecosystems. Through meticulous study, we may have uncovered a positive relationship: up to a specific limit, enhancing ocean alkalinity boosts photosynthesis by certain species.
France Van Wambeke, Pascal Conan, Mireille Pujo-Pay, Vincent Taillandier, Olivier Crispi, Alexandra Pavlidou, Sandra Nunige, Morgane Didry, Christophe Salmeron, and Elvira Pulido-Villena
Biogeosciences, 21, 2621–2640, https://doi.org/10.5194/bg-21-2621-2024, https://doi.org/10.5194/bg-21-2621-2024, 2024
Short summary
Short summary
Phosphomonoesterase (PME) and phosphodiesterase (PDE) activities over the epipelagic zone are described in the eastern Mediterranean Sea in winter and autumn. The types of concentration kinetics obtained for PDE (saturation at 50 µM, high Km, high turnover times) compared to those of PME (saturation at 1 µM, low Km, low turnover times) are discussed in regard to the possible inequal distribution of PDE and PME in the size continuum of organic material and accessibility to phosphodiesters.
Jenny Hieronymus, Magnus Hieronymus, Matthias Gröger, Jörg Schwinger, Raffaele Bernadello, Etienne Tourigny, Valentina Sicardi, Itzel Ruvalcaba Baroni, and Klaus Wyser
Biogeosciences, 21, 2189–2206, https://doi.org/10.5194/bg-21-2189-2024, https://doi.org/10.5194/bg-21-2189-2024, 2024
Short summary
Short summary
The timing of the net primary production annual maxima in the North Atlantic in the period 1750–2100 is investigated using two Earth system models and the high-emissions scenario SSP5-8.5. It is found that, for most of the region, the annual maxima occur progressively earlier, with the most change occurring after the year 2000. Shifts in the seasonality of the primary production may impact the entire ecosystem, which highlights the need for long-term monitoring campaigns in this area.
Nicole M. Travis, Colette L. Kelly, and Karen L. Casciotti
Biogeosciences, 21, 1985–2004, https://doi.org/10.5194/bg-21-1985-2024, https://doi.org/10.5194/bg-21-1985-2024, 2024
Short summary
Short summary
We conducted experimental manipulations of light level on microbial communities from the primary nitrite maximum. Overall, while individual microbial processes have different directions and magnitudes in their response to increasing light, the net community response is a decline in nitrite production with increasing light. We conclude that while increased light may decrease net nitrite production, high-light conditions alone do not exclude nitrification from occurring in the surface ocean.
Zoë Rebecca van Kemenade, Zeynep Erdem, Ellen Christine Hopmans, Jaap Smede Sinninghe Damsté, and Darci Rush
Biogeosciences, 21, 1517–1532, https://doi.org/10.5194/bg-21-1517-2024, https://doi.org/10.5194/bg-21-1517-2024, 2024
Short summary
Short summary
The California Current system (CCS) hosts the eastern subtropical North Pacific oxygen minimum zone (ESTNP OMZ). This study shows anaerobic ammonium oxidizing (anammox) bacteria cause a loss of bioavailable nitrogen (N) in the ESTNP OMZ throughout the late Quaternary. Anammox occurred during both glacial and interglacial periods and was driven by the supply of organic matter and changes in ocean currents. These findings may have important consequences for biogeochemical models of the CCS.
Cathy Wimart-Rousseau, Tobias Steinhoff, Birgit Klein, Henry Bittig, and Arne Körtzinger
Biogeosciences, 21, 1191–1211, https://doi.org/10.5194/bg-21-1191-2024, https://doi.org/10.5194/bg-21-1191-2024, 2024
Short summary
Short summary
The marine CO2 system can be measured independently and continuously by BGC-Argo floats since numerous pH sensors have been developed to suit these autonomous measurements platforms. By applying the Argo correction routines to float pH data acquired in the subpolar North Atlantic Ocean, we report the uncertainty and lack of objective criteria associated with the choice of the reference method as well the reference depth for the pH correction.
Sabine Mecking and Kyla Drushka
Biogeosciences, 21, 1117–1133, https://doi.org/10.5194/bg-21-1117-2024, https://doi.org/10.5194/bg-21-1117-2024, 2024
Short summary
Short summary
This study investigates whether northeastern North Pacific oxygen changes may be caused by surface density changes in the northwest as water moves along density horizons from the surface into the subsurface ocean. A correlation is found with a lag that about matches the travel time of water from the northwest to the northeast. Salinity is the main driver causing decadal changes in surface density, whereas salinity and temperature contribute about equally to long-term declining density trends.
Takamitsu Ito, Hernan E. Garcia, Zhankun Wang, Shoshiro Minobe, Matthew C. Long, Just Cebrian, James Reagan, Tim Boyer, Christopher Paver, Courtney Bouchard, Yohei Takano, Seth Bushinsky, Ahron Cervania, and Curtis A. Deutsch
Biogeosciences, 21, 747–759, https://doi.org/10.5194/bg-21-747-2024, https://doi.org/10.5194/bg-21-747-2024, 2024
Short summary
Short summary
This study aims to estimate how much oceanic oxygen has been lost and its uncertainties. One major source of uncertainty comes from the statistical gap-filling methods. Outputs from Earth system models are used to generate synthetic observations where oxygen data are extracted from the model output at the location and time of historical oceanographic cruises. Reconstructed oxygen trend is approximately two-thirds of the true trend.
Robert W. Izett, Katja Fennel, Adam C. Stoer, and David P. Nicholson
Biogeosciences, 21, 13–47, https://doi.org/10.5194/bg-21-13-2024, https://doi.org/10.5194/bg-21-13-2024, 2024
Short summary
Short summary
This paper provides an overview of the capacity to expand the global coverage of marine primary production estimates using autonomous ocean-going instruments, called Biogeochemical-Argo floats. We review existing approaches to quantifying primary production using floats, provide examples of the current implementation of the methods, and offer insights into how they can be better exploited. This paper is timely, given the ongoing expansion of the Biogeochemical-Argo array.
Qian Liu, Yingjie Liu, and Xiaofeng Li
Biogeosciences, 20, 4857–4874, https://doi.org/10.5194/bg-20-4857-2023, https://doi.org/10.5194/bg-20-4857-2023, 2023
Short summary
Short summary
In the Southern Ocean, abundant eddies behave opposite to our expectations. That is, anticyclonic (cyclonic) eddies are cold (warm). By investigating the variations of physical and biochemical parameters in eddies, we find that abnormal eddies have unique and significant effects on modulating the parameters. This study fills a gap in understanding the effects of abnormal eddies on physical and biochemical parameters in the Southern Ocean.
Caroline Ulses, Claude Estournel, Patrick Marsaleix, Karline Soetaert, Marine Fourrier, Laurent Coppola, Dominique Lefèvre, Franck Touratier, Catherine Goyet, Véronique Guglielmi, Fayçal Kessouri, Pierre Testor, and Xavier Durrieu de Madron
Biogeosciences, 20, 4683–4710, https://doi.org/10.5194/bg-20-4683-2023, https://doi.org/10.5194/bg-20-4683-2023, 2023
Short summary
Short summary
Deep convection plays a key role in the circulation, thermodynamics, and biogeochemical cycles in the Mediterranean Sea, considered to be a hotspot of biodiversity and climate change. In this study, we investigate the seasonal and annual budget of dissolved inorganic carbon in the deep-convection area of the northwestern Mediterranean Sea.
Daniela König and Alessandro Tagliabue
Biogeosciences, 20, 4197–4212, https://doi.org/10.5194/bg-20-4197-2023, https://doi.org/10.5194/bg-20-4197-2023, 2023
Short summary
Short summary
Using model simulations, we show that natural and anthropogenic changes in the global climate leave a distinct fingerprint in the isotopic signatures of iron in the surface ocean. We find that these climate effects on iron isotopes are often caused by the redistribution of iron from different external sources to the ocean, due to changes in ocean currents, and by changes in algal growth, which take up iron. Thus, isotopes may help detect climate-induced changes in iron supply and algal uptake.
Chloé Baumas, Robin Fuchs, Marc Garel, Jean-Christophe Poggiale, Laurent Memery, Frédéric A. C. Le Moigne, and Christian Tamburini
Biogeosciences, 20, 4165–4182, https://doi.org/10.5194/bg-20-4165-2023, https://doi.org/10.5194/bg-20-4165-2023, 2023
Short summary
Short summary
Through the sink of particles in the ocean, carbon (C) is exported and sequestered when reaching 1000 m. Attempts to quantify C exported vs. C consumed by heterotrophs have increased. Yet most of the conducted estimations have led to C demands several times higher than C export. The choice of parameters greatly impacts the results. As theses parameters are overlooked, non-accurate values are often used. In this study we show that C budgets can be well balanced when using appropriate values.
Anna Belcher, Sian F. Henley, Katharine Hendry, Marianne Wootton, Lisa Friberg, Ursula Dallman, Tong Wang, Christopher Coath, and Clara Manno
Biogeosciences, 20, 3573–3591, https://doi.org/10.5194/bg-20-3573-2023, https://doi.org/10.5194/bg-20-3573-2023, 2023
Short summary
Short summary
The oceans play a crucial role in the uptake of atmospheric carbon dioxide, particularly the Southern Ocean. The biological pumping of carbon from the surface to the deep ocean is key to this. Using sediment trap samples from the Scotia Sea, we examine biogeochemical fluxes of carbon, nitrogen, and biogenic silica and their stable isotope compositions. We find phytoplankton community structure and physically mediated processes are important controls on particulate fluxes to the deep ocean.
Asmita Singh, Susanne Fietz, Sandy J. Thomalla, Nicolas Sanchez, Murat V. Ardelan, Sébastien Moreau, Hanna M. Kauko, Agneta Fransson, Melissa Chierici, Saumik Samanta, Thato N. Mtshali, Alakendra N. Roychoudhury, and Thomas J. Ryan-Keogh
Biogeosciences, 20, 3073–3091, https://doi.org/10.5194/bg-20-3073-2023, https://doi.org/10.5194/bg-20-3073-2023, 2023
Short summary
Short summary
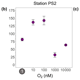
Despite the scarcity of iron in the Southern Ocean, seasonal blooms occur due to changes in nutrient and light availability. Surprisingly, during an autumn bloom in the Antarctic sea-ice zone, the results from incubation experiments showed no significant photophysiological response of phytoplankton to iron addition. This suggests that ambient iron concentrations were sufficient, challenging the notion of iron deficiency in the Southern Ocean through extended iron-replete post-bloom conditions.
Benoît Pasquier, Mark Holzer, Matthew A. Chamberlain, Richard J. Matear, Nathaniel L. Bindoff, and François W. Primeau
Biogeosciences, 20, 2985–3009, https://doi.org/10.5194/bg-20-2985-2023, https://doi.org/10.5194/bg-20-2985-2023, 2023
Short summary
Short summary
Modeling the ocean's carbon and oxygen cycles accurately is challenging. Parameter optimization improves the fit to observed tracers but can introduce artifacts in the biological pump. Organic-matter production and subsurface remineralization rates adjust to compensate for circulation biases, changing the pathways and timescales with which nutrients return to the surface. Circulation biases can thus strongly alter the system’s response to ecological change, even when parameters are optimized.
Priyanka Banerjee
Biogeosciences, 20, 2613–2643, https://doi.org/10.5194/bg-20-2613-2023, https://doi.org/10.5194/bg-20-2613-2023, 2023
Short summary
Short summary
This study shows that atmospheric deposition is the most important source of iron to the upper northern Indian Ocean for phytoplankton growth. This is followed by iron from continental-shelf sediment. Phytoplankton increase following iron addition is possible only with high background levels of nitrate. Vertical mixing is the most important physical process supplying iron to the upper ocean in this region throughout the year. The importance of ocean currents in supplying iron varies seasonally.
Iris Kriest, Julia Getzlaff, Angela Landolfi, Volkmar Sauerland, Markus Schartau, and Andreas Oschlies
Biogeosciences, 20, 2645–2669, https://doi.org/10.5194/bg-20-2645-2023, https://doi.org/10.5194/bg-20-2645-2023, 2023
Short summary
Short summary
Global biogeochemical ocean models are often subjectively assessed and tuned against observations. We applied different strategies to calibrate a global model against observations. Although the calibrated models show similar tracer distributions at the surface, they differ in global biogeochemical fluxes, especially in global particle flux. Simulated global volume of oxygen minimum zones varies strongly with calibration strategy and over time, rendering its temporal extrapolation difficult.
John C. Tracey, Andrew R. Babbin, Elizabeth Wallace, Xin Sun, Katherine L. DuRussel, Claudia Frey, Donald E. Martocello III, Tyler Tamasi, Sergey Oleynik, and Bess B. Ward
Biogeosciences, 20, 2499–2523, https://doi.org/10.5194/bg-20-2499-2023, https://doi.org/10.5194/bg-20-2499-2023, 2023
Short summary
Short summary
Nitrogen (N) is essential for life; thus, its availability plays a key role in determining marine productivity. Using incubations of seawater spiked with a rare form of N measurable on a mass spectrometer, we quantified microbial pathways that determine marine N availability. The results show that pathways that recycle N have higher rates than those that result in its loss from biomass and present new evidence for anaerobic nitrite oxidation, a process long thought to be strictly aerobic.
Amanda Gerotto, Hongrui Zhang, Renata Hanae Nagai, Heather M. Stoll, Rubens César Lopes Figueira, Chuanlian Liu, and Iván Hernández-Almeida
Biogeosciences, 20, 1725–1739, https://doi.org/10.5194/bg-20-1725-2023, https://doi.org/10.5194/bg-20-1725-2023, 2023
Short summary
Short summary
Based on the analysis of the response of coccolithophores’ morphological attributes in a laboratory dissolution experiment and surface sediment samples from the South China Sea, we proposed that the thickness shape (ks) factor of fossil coccoliths together with the normalized ks variation, which is the ratio of the standard deviation of ks (σ) over the mean ks (σ/ks), is a robust and novel proxy to reconstruct past changes in deep ocean carbon chemistry.
Katherine E. Turner, Doug M. Smith, Anna Katavouta, and Richard G. Williams
Biogeosciences, 20, 1671–1690, https://doi.org/10.5194/bg-20-1671-2023, https://doi.org/10.5194/bg-20-1671-2023, 2023
Short summary
Short summary
We present a new method for reconstructing ocean carbon using climate models and temperature and salinity observations. To test this method, we reconstruct modelled carbon using synthetic observations consistent with current sampling programmes. Sensitivity tests show skill in reconstructing carbon trends and variability within the upper 2000 m. Our results indicate that this method can be used for a new global estimate for ocean carbon content.
Cited articles
Abram, N. J., Mulvaney, R., Vimeux, F., Phipps, S. J., Turner, J., and England, M. H.:
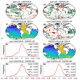
Evolution of the Southern Annular Mode during the past millennium, Nat. Clim. Change, 4, 564–569, https://doi.org/10.1038/nclimate2235, 2014. a, b
Alldredge, A. L. and Gotschalk, C. C.: Direct observations of the mass flocculation of diatom blooms: characteristics, settling velocities and formation of diatom aggregates, Deep-Sea Res. Pt. I, 36, 159–171, https://doi.org/10.1016/0198-0149(89)90131-3, 1989. a
Anderson, M. J. and Willis, T. J.: Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology, Ecology, 84, 511–525, https://doi.org/10.1890/0012-9658(2003)084[0511:CAOPCA]2.0.CO;2, 2003. a
Arblaster, J. M. and Meehl, G. A.: Contributions of external forcings to southern annular mode trends, J. Clim., 19, 2896–2905, https://doi.org/10.1175/JCLI3774.1, 2006. a, b, c, d
Arndt, J.E., Schenke, H.W., Jakobsson, M., Nitsche, F.O., Buys, G., Goleby, B., Rebesco, M., Bohoyo, F., Hong, J., Black, J., and Greku, R.: The International Bathymetric Chart of the Southern Ocean (IBCSO) Version 1.0 – A new bathymetric compilation covering circum‐Antarctic waters, Geophys. Res. Lett., 40, 3111–3117, https://doi.org/10.1002/grl.50413, 2013. a
Arrigo, K. R., Robinson, D. H., Worthen, D., Dunbar, R. B., DiTullio, G. R., VanWoert, M., and Lizotte, M. P.: Phytoplankton Community Structure and the Drawdown of Nutrients and CO2 in the Southern Ocean, Science, 283, 365–367. https://doi.org/10.1126/science.283.5400.365, 1999. a, b
Arrigo, K. R., van Dijken, G. L., and Bushinsky, S.: Primary production in the Southern Ocean, 1997–2006, J. Geophys. Res.-Ocean., 113, 1997–2006, https://doi.org/10.1029/2007JC004551, 2008. a, b, c, d
Belgrano, A., Lindahl, O., and Hernroth, B.: North Atlantic Oscillation primary productivity and toxic phytoplankton in the Gullmar Fjord, Sweden (1985–1996), P. Roy. Soc. B, 266, 425–430, https://doi.org/10.1098/rspb.1999.0655, 1999. a
Blanchot, J., Rodier, M., and Le Bouteiller, A.: Effect of el niño southern oscillation events on the distribution and abundance of phytoplankton in the Western Pacific Tropical Ocean along 165∘ E, J. Plankton Res., 14, 137–156, https://doi.org/10.1093/plankt/14.1.137, 1992. a
Blenckner, T. and Hillebrand, H.: North Atlantic Oscillation signatures in aquatic and terrestrial ecosystems – A meta-analysis, Glob. Change Biol., 8, 203–212, https://doi.org/10.1046/j.1365-2486.2002.00469.x, 2002. a
Bopp, L., Resplandy, L., Orr, J. C., Doney, S. C., Dunne, J. P., Gehlen, M., Halloran, P., Heinze, C., Ilyina, T., Séférian, R., Tjiputra, J., and Vichi, M.: Multiple stressors of ocean ecosystems in the 21st century: projections with CMIP5 models, Biogeosciences, 10, 6225–6245, https://doi.org/10.5194/bg-10-6225-2013, 2013. a, b
Boyce, D. G., Lewis, M. R., and Worm, B.: Global phytoplankton decline over the past century, Nature, 466, 591–596, https://doi.org/10.1038/nature09268, 2010. a, b, c
Boyce, D., Lewis, M., and Worm, B.: Boyce et al. reply, Nature, 472, E8–E9, https://doi.org/10.1038/nature09953, 2011. a
Boyd, C. M., Heyraud, M., and Boyd, C. N.: Feeding of the Antarctic krill Euphausia superba, J. Crustacean Biol., 4, 123–141, https://doi.org/10.1163/1937240X84X00543, 1984. a
Boyd, P. W. and Trull, T. W.: Understanding the export of biogenic particles in oceanic waters: Is there consensus?, Prog. Oceanogr., 72, 276–312, https://doi.org/10.1016/j.pocean.2006.10.007, 2007. a
Bray, J. R. and Curtis, J. T.: An Ordination of the Upland Forest Communities of Southern Wisconsin, Ecol. Monogr., 27, 325–349, https://doi.org/10.2307/1942268, 1957. a
Burckle, L. H., Jacobs, S. S., and McLaughlin, R. B.: Late austral spring diatom distribution between
New Zealand and the Ross Ice Shelf, Antarctica: Hydrographic and sediment correlations,
Micropaleontology, 74–81, https://doi.org/10.2307/1485528, 1987. a
Cadée, G. C., González, H., and Schnack-Schiel, S. B.: Krill diet affects faecal string settling, in: Weddell Sea Ecology, Springer, Berlin, Germany, 1992. a
Carranza, M. M. and Gille, S. T.: Southern Ocean wind‐driven entrainment enhances satellite chlorophyll‐a through the summer, J. Geophys. Res.-Ocean., 120, 304–323, https://doi.org/10.1002/2014JC010203, 2015. a, b, c, d
Cavicchioli, R., Ripple, W. J., Timmis, K. N., Azam, F., Bakken, L. R., Baylis, M., Behrenfeld, M. J.,
Boetius, A., Boyd, P. W., Classen, A. T. and Crowther, T. W.: Scientists’ warning to humanity:
microorganisms and climate change, Nat. Rev. Microbiol., 1, 569–586,
https://doi.org/10.1038/s41579-019-0222-5, 2019. a
Chiba, S., Hirawake, T., Ushio, S., Horimoto, N., Satoh, R., Nakajima, Y., Ishimaru, T., and Yamaguchi,
Y.: An overview of the biological/oceanographic survey by the RTV Umitaka-Maru III off Adelie Land, Antarctica in January–February 1996, Deep-Sea Res. Pt. II, 47, 2589–2613, https://doi.org/10.1016/S0967-0645(00)00037-0, 2000. a
Ciais, P., Sabine, C., Bala, G., Bopp, L., Brovkin, V., Canadell, J., Chhabra, A., DeFries, R., Galloway, J.,
Heimann, M., and Jones, C.: Carbon and other biogeochemical cycles. Climate change 2013: the physical science basis, Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge University Press Cambridge United Kingdom and New York NY USA, 465–570, https://doi.org/10.1017/CBO9781107415324.015, 2013. a, b
Clarke, K. R., Somerfield, P. J., and Gorley, R. N.: Testing of null hypotheses in exploratory community analyses: similarity profiles and biota-environment linkage, J. Exp. Mar. Biol. Ecol., 366, 56–69, https://doi.org/10.1016/j.jembe.2008.07.009, 2008. a
Cohen, J.: Things I have learned (so far) some things you learn aren't so less is more, Am. Psychol., 45, 1304–1312, https://doi.org/10.1037/0003-066X.45.12.1304, 1990. a
Constable, A. J., Melbourne-Thomas, J., Corney, S. P., Arrigo, K. R., Barbraud, C., Barnes, D. K. A., Bindoff, N. L., Boyd, P. W., Brandt, A., Costa, D. P., and Davidson, A. T.: Climate change and Southern Ocean ecosystems I: How changes in physical habitats directly affect marine biota, Glob. Change Biol., 20, 3004–3025, https://doi.org/10.1111/gcb.12623, 2014. a
Dargie, T. C. D.: On the integrated interpretation of indirect site ordinations: a case study using semi-arid vegetation in southeastern Spain, Vegetatio, 55, 37–55, https://doi.org/10.1007/BF00039980, 1984. a
Davidson, A. T., McKinlay, J., Westwood, K., Thomson, P. G., van den Enden, R., de Salas, M., Wright,
S., Johnson, R., and Berry, K.: Enhanced CO2 concentrations change the structure of Antarctic marine microbial communities, Mar. Ecol. Prog. Ser., 552, 93–113, https://doi.org/10.3354/meps11742, 2016. a
Deppeler, S. L. and Davidson, A. T.: Southern Ocean phytoplankton in a changing climate, Front. Mar. Sci., 4, 1–28, https://doi.org/10.3389/fmars.2017.00040, 2017. a
DiFiore, P. J., Sigman, D. M., Trull, T. W., Lourey, M. J., Karsh, K., Cane, G., and Ho, R.: Nitrogen isotope constraints on subantarctic biogeochemistry, J. Geophys. Res.-Ocean., 111, 1–19, https://doi.org/10.1029/2005JC003216, 2006. a
Dixon, P.: VEGAN, a package of R functions for community ecology. J. Veg. Sci., 14, 631–637, https://doi.org/10.1111/j.1654-1103.2003.tb02228.x, 2003. a
Fitch, D. T. and Moore, J. K.: Wind speed influence on phytoplankton bloom dynamics in the Southern Ocean Marginal Ice Zone, J. Geophys. Res.-Ocean., 112, 1–13, https://doi.org/10.1029/2006JC004061, 2007. a, b, c
Fryxell, G. A.: Marine phytoplankton at the Weddell Sea ice edge: Seasonal changes at the specific level, Polar Biol., 10, 6–7, https://doi.org/10.1007/BF00238285, 1989. a
Gibbons, J. D. and Pratt, J. W.: P-values: interpretation and methodology, Am. Stat., 29, 20–25, https://doi.org/10.1080/00031305.1975.10479106, 1975. a
Gillett, N. P. and Fyfe, J. C.: Annular mode changes in the CMIP5 simulations, Geophys. Res. Lett., 40, 1189–1193, https://doi.org/10.1002/grl.50249, 2013. a, b, c
Gomi, Y., Taniguchi, A., and Fukuchi, M.: Temporal and spatial variation of the phytoplankton assemblage in the eastern Indian sector of the Southern Ocean in summer 2001/2002, Polar Biol., 30, 817–827, https://doi.org/10.1007/s00300-006-0242-2, 2007. a
Gong, D. and Wang, S.: Definition of Antarctic oscillation index, Geophys. Res. Lett., 26, 459–462, https://doi.org/10.1029/1999GL900003, 1999. a, b
Greaves, B. L., Davidson, A. T., and Fraser, A. D.: The Southern Annular Mode (SAM) influences phytoplankton communities in the seasonal ice zone of the Southern Ocean, Ver. 1, Australian Antarctic Data Centre, https://doi.org/10.26179/5d9181f7308bd, 2019. a
Green, S. E. and Sambrotto, R. N.: Plankton community structure and export of C, N, P and Si in the Antarctic Circumpolar Current, Deep-Sea Res. Pt. II, 53, 620–643, https://doi.org/10.1016/j.dsr2.2006.01.022, 2006. a
Hall, A. and Visbeck, M.: Synchronous variability in the Southern Hemisphere atmosphere, sea ice, and ocean resulting from the annular mode, J. Clim., 15, 3043–3057, https://doi.org/10.1175/1520-0442(2002)015<3043:SVITSH>2.0.CO;2, 2002. a, b
Harris, R. M. B., Beaumont, L. J., Vance, T. R., Tozer, C. R., Remenyi, T. A., Perkins-Kirkpatrick, S. E., Mitchell, P.J., Nicotra, A.B., McGregor, S., Andrew, N.R. Letnic, M., Kearney, M. R., Wernberg, T.,
Hutley, L. B., Chambers, L. E., Fletcher, M.-S., Keatley, M. R., Woodward, C. A., Williamson, G., Duke,
N. C., and Bowman, D. M. J. S.: Biological responses to the press and pulse of climate trends and extreme events, Nat. Clim. Change, 8, 579–587, https://doi.org/10.1038/s41558-018-0187-9, 2018. a
Henson, S. A., Yool, A., and Sanders, R. Variability in efficiency of particulate organic carbon export: A model study, Global Biogeochem. Cy., 29, 33–45, https://doi.org/10.1002/2014GB004965, 2015. a, b, c
Hines, K. M., Bromwich, D. H., and Marshall, G. J.: Artificial surface pressure trends in the NCEP-NCAR reanalysis over the Southern Ocean and Antartica, J. Clim., 13, 3940–3952, https://doi.org/10.1175/1520-0442(2000)013<3940:ASPTIT>2.0.CO;2, 2000. a
Ho, M., Kiem, A. S., and Verdon-Kidd, D. C.: The Southern Annular Mode: a comparison of indices, Hydrol. Earth Syst. Sci., 16, 967–982, https://https://doi.org/10.5194/hess-16-967-2012, 2012. a, b, c
Hoegh-Guldberg, O. and Bruno, J. F.: The impact of climate change on the world's marine ecosystems, Science, 328, 1523–1528, https://doi.org/10.1126/science.1189930, 2010. a
Hydes, D. J., Aoyama, M., Aminot, A., Bakker, K., Becker, S., Coverly, S., Daniel, A., Dickson, A. G.,
Grosso, O., Kerouel, R., van Ooijen, J., Sato, K., Tanhua, T., Woodward, E. M. S., and Zhang, J. Z.:
Determination of Dissolved Nutrients (N, P, SI) in Seawater With High Precision and Inter-Comparability Using Gas-Segmented Continuous Flow Analysers, in: The GOSHIP repeat hydrography
manual: a collection of expert reports and guidelines, edited by: Hood, E. M., Sabine, C. L., and Sloyan, B. M., IOCCP report number 14, ICPO publication series number 134, UNESCO-IOC, Paris,
France, available at: http://www.go-ship.org/HydroMan.html (last access: 15 January 2020), 2010. a
Clem, K. R., Crosta,
X., de Lavergne, C., Eisenman, I., England, M. H., Fogt, R. L., Frankcombe, L. M., Marshall, G. J.,
Masson-Delmotte, V., Morrison, A. K., Orsi, A. J., Raphael, M. N., Renwick, J. A., Schneider, D. P.,
Simpkins, G. R., Steig, E. J., Stenni, B., Swingedouw, D., and Vance, T. R.: Assessing recent trends in high-latitude Southern Hemisphere surface climate, Nat. Clim. Change, 6, 917–926, https://doi.org/10.1038/nclimate3103, 2016. a
Kahru, M., Brotas, V., Manzano-Sarabia, M., and Mitchell, B. G.: Are phytoplankton blooms occurring earlier in the Arctic?, Glob. Change Biol., 17, 1733–1739, https://doi.org/10.1111/j.1365-2486.2010.02312.x, 2011. a, b
Kawaguchi, S., Ichii, T., and Naganobu, M.: Green krill, the indicator of micro-and nano-size phytoplankton availability to krill, Polar Biol., 22, 133–136, 1999. a
Kohyama, T. and Hartmann, D. L.: Antarctic sea ice response to weather and climate modes of variability, J. Clim.e, 29, 721–741. https://doi.org/10.1175/JCLI-D-15-0301.1, 2016. a
Kwok, R. and Comiso, J. C.: Southern Ocean climate and sea ice anomalies associated with the Southern Oscillation, J. Clim., 15, 487–501, https://doi.org/10.1175/1520-0442(2002)015<0487:SOCASI>2.0.CO;2, 2002. a
Lampitt, R. S. and Antia, A. N.: Particle flux in deep seas: Regional characteristics and temporal variability, Deep-Sea Res. Pt. I, 44, 1377–1403, https://doi.org/10.1016/S0967-0637(97)00020-4, 1997. a
Lannuzel, D., Schoemann, V., de Jong, J., Tison, J. L., and Chou, L.: Distribution and biogeochemical behaviour of iron in the East Antarctic sea ice, Mar. Chem., 106, 18–32, https://doi.org/10.1016/j.marchem.2006.06.010, 2007. a
Lefebvre, W., Goosse, H., Timmermann, R., and Fichefet, T.: Influence of the Southern Annular Mode on the sea ice-ocean system, J. Geophys. Res.-Ocean., 109, 1–12, https://doi.org/10.1029/2004JC002403, 2004. a, b
Legendre, P. and Anderson, M. J.: Distance‐based redundancy analysis: testing multispecies responses in multifactorial ecological experiments, Ecol. Monogr., 69, 1–24, https://doi.org/10.1890/0012-9615(1999)069[0001:DBRATM]2.0.CO;2, 1999. a
Legendre, P., Oksanen, J., and ter Braak, C. J.: Testing the significance of canonical axes in redundancy analysis, Methods Ecol. Evol., 2, 269–277, https://doi.org/10.1111/j.2041-210X.2010.00078.x, 2011. a
Lenton, A. and Matear, R. J.: Role of the Southern Annular Mode (SAM) in Southern Ocean CO2 uptake, Global Biogeochem. Cy., 21, 1-17, https://doi.org/10.1029/2006GB002714, 2007. a
Lohbeck, K. T., Riebesell, U., and Reusch, T. B. H.: Gene expression changes in the coccolithophore Emiliania huxleyi after 500 generations of selection to ocean acidification, P. Roy. Soc. B, 281, 1–7, https://doi.org/10.1098/rspb.2014.0003, 2014. a
Lovenduski, N. S., Gruber, N., Doney, S. C., and Lima, I. D.: Enhanced CO2 outgassing in the Southern Ocean from a positive phase of the Southern Annular Mode, Global Biogeochem. Cy., 21, 1–14, https://doi.org/10.1029/2006GB002900, 2007. a
Mackas, D. L.: Does blending of chlorophyll data bias temporal trend?, Nature, 472, E4–E5, https://doi.org/10.1038/nature09951, 2011. a
Mackintosh, A. N., Anderson, B. M., Lorrey, A. M., Renwick, J. A., Frei, P., and Dean, S. M.: Regional cooling caused recent New Zealand glacier advances in a period of global warming, Nat. Commun., 8, 1–13, https://doi.org/10.1038/ncomms14202, 2017. a
Marshall, G. J.: Trends in the Southern Annular Mode from Observations and Reanalyses, J. Clim., 16, 4134–4143, https://doi.org/10.1175/1520-0442(2003)016<4134:TITSAM>2.0.CO;2, 2003. a, b
Marshall, G. J.: Half-century seasonal relationships between the Southern Annular mode and Antarctic temperatures, Int. J. Climatol., 27, 373–383, https://doi.org/10.1002/joc.1407, 2007. a, b
Martin, A., McMinn, A., Heath, M., Hegseth, E. N., and Ryan, K. G.: The physiological response to increased temperature in over-wintering sea ice algae and phytoplankton in McMurdo Sound, Antarctica and Tromsø Sound, Norway, J. Exp. Mar. Biol. Ecol., 428, 57–66, https://doi.org/10.1016/j.jembe.2012.06.006, 2012. a
Massom, R. A. and Stammerjohn, S. E.: Antarctic sea ice change and variability – Physical and ecological implications, Polar Sci., 4, 149–186, https://doi.org/10.1016/j.polar.2010.05.001, 2010. a, b
McMinn, A., Ashworth, C., and Ryan, K.: Growth and Productivity of Antarctic Sea Ice Algae under PAR and UV Irradiances, Bot. Mar., 42, 401–407, https://doi.org/10.1515/BOT.1999.046, 1999. a
McMinn, A. and Martin, A.: Dark survival in a warming world, P. Roy. Soc. B, 280, 20122909, https://doi.org/10.1098/rspb.2012.2909, 2013. a, b
Meredith, M. P., Murphy, E. J., Hawker, E. J., King, J. C., and Wallace, M. I.: On the interannual variability of ocean temperatures around South Georgia, Southern Ocean: Forcing by El Niño/Southern Oscillation and the Southern Annular Mode, Deep-Sea Res. Pt. II, 55, 2007–2022, https://doi.org/10.1016/j.dsr2.2008.05.020, 2008. a, b
Mo, K. C.: Relationships between low-frequency variability in the Southern Hemisphere and sea surface temperature anomalies, J. Clim., 13, 3599–3610, https://doi.org/10.1175/1520-0442(2000)013<3599:rblfvi>2.0.co;2, 2000. a
Moline, M. A., Claustre, H., Frazer, T. K., Schofield, O., and Vernet, M.: Alteration of the food web along the Antarctic Peninsula in response to a regional warming trend, Glob. Change Biol., 10, 1973–1980, https://doi.org/10.1111/j.1365-2486.2004.00825.x, 2004. a
Moore, J. K. and Abbott, M. R.: Phytoplankton chlorophyll distributions and primary production in the Southern Ocean, J. Geophys. Res.-Ocean., 105, 28709–28722, https://doi.org/10.1029/1999JC000043, 2000. a, b
Nakagawa, S.: A farewell to Bonferroni: the problems of low statistical power and publication bias, Behav. Ecol., 15, 1044–1045, https://doi.org/10.1093/beheco/arh107, 2004. a
Nakagawa, S. and Cuthill, I. C.: Effect size, confidence interval and statistical significance: a practical guide for biologists, Biol. Rev., 82, 591–605, https://doi.org/10.1111/j.1469-185X.2007.00027.x, 2007. a
Nehring, S.: Establishment of thermophilic phytoplankton species in the North Sea: biological indicators of climatic changes?, Short communication, ICES J. Mar. Sci., 55, 818–823, https://doi.org/10.1006/jmsc.1998.0389, 1998. a, b
NOAA: Teleconnection Pattern Calculation Procedures, Climate Prediction Center Internet Team, available at: https://www.cpc.ncep.noaa.gov/products/precip/CWlink/daily_ao_index/history/method.shtml#var (last access: 15 June 2017), 2005. a
OBIS: Ocean Biogeographic Information System. Intergovernmental Oceanographic Commission of UNESCO, available at: http://www.iobis.org, last access: 18 February 2020.
Ottersen, G., Planque, B., Belgrano, A., Post, E., Reid, P. C., and Stenseth, N. C.: Ecological effects of the North Atlantic Oscillation, Oecologia, 128, 1–14, https://doi.org/10.1007/s004420100655, 2001. a, b
Parkinson, C. L.: A 40-y record reveals gradual Antarctic sea ice increases followed by decreases at rates far exceeding the rates seen in the Arctic, P. Natl. Acad. Sci. USA, 116, 14414–14423, https://doi.org/10.1073/pnas.1906556116, 2019. a, b
R Core Team: R: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing, Vienna, Austria, 2016. a
Rigual-Hernández, A. S., Trull, T. W., Bray, S. G., Closset, I., and Armand, L. K.: Seasonal dynamics in diatom and particulate export fluxes to the deep sea in the Australian sector of the southern Antarctic Zone, J. Mar. Syst., 142, 62–74, https://doi.org/10.1016/j.jmarsys.2014.10.002, 2015. a
Roach, L. A., Smith, M. M., and Dean, S. M.: Quantifying growth of pancake sea ice floes using images from drifting buoys, J. Geophys. Res.-Ocean., 123, 2851–2866, https://doi.org/10.1002/2017JC013693, 2018. a
Rodgers, J. L. and Nicewander, W. A.: Thirteen Ways to Look at the Correlation Coefficient, Am. Stat., 42, 59–66, https://doi.org/10.1080/00031305.1988.10475524, 1988. a
Saenz, B. T. and Arrigo, K. R.: Annual primary production in Antarctic sea ice during 2005-2006 from a sea ice state estimate, J. Geophys. Res.-Ocean., 119, 3645–3678, https://doi.org/10.1002/2013JC009677, 2014. a
Sarthou, G., Timmermans, K. R., Blain, S., and Tréguer, P.: Growth physiology and fate of diatoms in the ocean: a review, J. Sea Res., 53, 25–42, https://doi.org/10.1016/j.seares.2004.01.007, 2005. a
Savidge, G., Priddle, J., Gilpin, L. C., Bathmann, U., Murphy, E. J., Owens, N. J. P., Pollard, R. T.,
Turner, D. R., Veth, C., and Boyd, P.: An assessment of the role of the marginal ice zone in the carbon cycle of the Southern Ocean, Antarct. Sci., 8, 349–358, https://doi.org/10.1017/S0954102096000521, 1996. a
Scheffers, B. R., De Meester, L., Bridge, T. C. L., Hoffmann, A. A., Pandolfi, J. M., Corlett, R. T., Butchart, S. H. M., Pearce-Kelly, P., Kovacs, K. M., Dudgeon, D., Pacifici, M., Rondinini, C., Foden, W.
B., Martin, T. G., Mora, C., Bickford, D., and Watson, J. E. M.: The broad footprint of climate change from genes to biomes to people, Science, 354, aaf7671, https://doi.org/10.1126/science.aaf7671, 2016. a
Schiermeier, Q.: Atmospheric science: fixing the sky, Nature, 460, 792–795, https://doi.org/10.1038/460792a, 2009. a
Schlüter, L., Lohbeck, K. T., Gutowska, M. A., Gröger, J. P., Riebesell, U., and Reusch, T. B. H.: Adaptation of a globally important coccolithophore to ocean warming and acidification, Nat. Clim. Change, 4, 1024–1030, https://doi.org/10.1038/nclimate2379, 2014. a
Scott, F. J. and Marchant, H. J. (Eds): Antarctic marine protists, Australian Biological Resources Study, Canberra and Hobart, Australia, 541 pp., https://doi.org/10.1017/s0032247405244819, 2005. a
Sen Gupta, A. and England, M. H.: Coupled ocean–atmosphere–ice response to variations in the Southern Annular Mode, J. Clim., 19, 4457–4486, https://doi.org/10.1175/JCLI3843.1, 2006. a, b
Smetacek, V. and Nicol, S.: Polar ocean ecosystems in a changing world, Nature, 437, 362–368, https://doi.org/10.1038/nature04161, 2005. a
Solomon, S., Ivy, D. J., Kinnison, D., Mills, M. J., Neely, R. R., and Schmidt, A.: Emergence of healing in the Antarctic ozone layer, Science, 353, 269–274, https://doi.org/10.1126/science.aae0061, 2016. a, b
Son, S. W., Tandon, N. F., Polvani, L. M., and Waugh, D. W.: Ozone hole and Southern Hemisphere climate change, Geophys. Res. Lett., 36, 1–5, https://doi.org/10.1029/2009GL038671, 2009. a
Soppa, M., Völker, C., and Bracher, A.: Diatom Phenology in the Southern Ocean: Mean Patterns, Trends and the Role of Climate Oscillations, Remote Sens., 8, 1–7, https://doi.org/10.3390/rs8050420, 2016. a, b
Spreen, G., Kaleschke, L., and Heygster, G.: Sea ice remote sensing using AMSR-E 89-GHz channels, J. Geophys. Res.-Ocean., 113, C02S03, https://doi.org/10.1029/2005JC003384, 2008. a
Squire, V. A.: Ocean wave interactions with sea ice: a reappraisal, Annu. Rev. Fluid Mech., 52, 37–60, https://doi.org/10.1146/annurev-fluid-010719-060301, 2020. a
Steinacher, M., Joos, F., Frölicher, T. L., Bopp, L., Cadule, P., Cocco, V., Doney, S. C., Gehlen, M., Lindsay, K., Moore, J. K., Schneider, B., and Segschneider, J.: Projected 21st century decrease in marine productivity: a multi-model analysis, Biogeosciences, 7, 979–1005, https://doi.org/10.5194/bg-7-979-2010, 2010. a
Swart, N. C. and Fyfe, J. C.: Observed and simulated changes in the Southern Hemisphere surface westerly wind-stress, Geophys. Res. Lett., 39, 1–6, https://doi.org/10.1029/2012GL052810, 2012. a, b
Swart, N. C., Fyfe, J. C., Gillett, N., and Marshall, G. J.: Comparing Trends in the Southern Annular Mode and Surface Westerly Jet, J. Clim., 28, 8840–8859, https://doi.org/10.1175/JCLI-D-15-0334.1, 2015. a
Świło, M., Majewski, W., Minzoni, R. T., and Anderson, J. B.: Diatom assemblages from coastal settings of West Antarctica, Mar. Micropaleontol., 125, 95–109, https://doi.org/10.1016/j.marmicro.2016.04.001, 2016. a
Takahashi, T., Sutherland, S. C., Wanninkhof, R., Sweeney, C., Feely, R. A., Chipman, D. W., Hales, B.,
Friederich, G., Chavez, F., Sabine, C., Watson, A., Bakker, D. C. E., Schuster, U., Metzl, N., Yoshikawa-Inoue, H., Ishii, M., Midorikawa, T., Nojiri, Y., Körtzinger, A., Steinhoff, T., Hoppema, M., Olafsson,
J., Arnarson, T. S., Tilbrook, B., Johannessen, T., Olsen, A., Bellerby, R., Wong, C. S., Delille, B., Bates, N. R., and de Baar, H. J. W.: Climatological mean and decadal change in surface ocean pCO2, and net sea–air CO2 flux over the global oceans, Deep-Sea Res. Pt. II, 56, 554–577, https://doi.org/10.1016/j.dsr2.2008.12.009, 2009. a, b
Taljaard, J. J.: Development, Distribution and Movement of Cyclones and Anticyclones in the Southern Hemisphere During the IGY, J. Appl. Meteorol., 6, 973–987, https://doi.org/10.1175/1520-0450(1967)006<0973:DDAMOC>2.0.CO;2, 1967. a
Taylor, F. and Sjunneskog, C.: Postglacial marine diatom record of the Palmer Deep, Antarctic
Peninsula (ODP Leg 178, Site 1098), 2. Diatom assemblages, Paleoceanography, 17, 1–12, https://doi.org/10.1029/2000PA000564, 2002. a
Ter Braak, C. J. and Verdonschot, P. F.: Canonical correspondence analysis and related multivariate methods in aquatic ecology, Aquat. Sci., 57, 255–289, https://doi.org/10.1007/BF00877430, 1995. a
Thompson, D. W., Lee, S., and Baldwin, M. P.: Atmospheric processes governing the northern hemisphere annular mode/North Atlantic oscillation, Geoph. Monog. Series, 134, 81–112, 2003. a
Thompson, D. W. J. and Solomon, S.: Interpretation of Recent Southern Hemisphere Climate Change, Science, 296, 895–899, https://doi.org/10.1126/science.1069270, 2002. a, b
Tomas, C. R. (Ed.): Identifying marine phytoplankton, Academic press, San Diego, California, 858 pp., 1997. a
Turner, J., Bracegirdle, T. J., Phillips, T., Marshall, G. J., and Hosking, J. S.: An initial assessment of Antarctic sea ice extent in the CMIP5 models, J. Clim., 26, 1473–1484, 2013. a
Turner, J., Summerhayes, C., Sparrow, M., Mayewski, P., Convey, P., di Prisco, G., Gutt, J., Hodgson, D., Speich, S., Worby, T., Bo, S., and Klepikov, A.: Antarctic climate change and the environment – 2015 update, Antarctic Treaty Consultative Meeting, Sofia, Bulgaria, June 2015, IP 92, 2015. a
Waters, R. L., Van Den Enden, R., and Marchant, H. J.: Summer microbial ecology off East Antarctica (80–150∘ E): protistan community structure and bacterial abundance, Deep-Sea Res. Pt. II, 47, 2401–2435, https://doi.org/10.1016/S0967-0645(00)00030-8, 2000. a
Webb, T. and Bryson, R. A.: Late-and postglacial climatic change in the northern Midwest, USA: quantitative estimates derived from fossil pollen spectra by multivariate statistical analysis, Quaternary Res., 2, 70–115, https://doi.org/10.1016/0033-5894(72)90005-1, 1972. a
Whitaker, D. and Christman, M.: clustsig: Significant Cluster Analysis, R package version 1.1., 2014. a
Wilson, D. L., Smith Jr, W. O., and Nelson, D. M.: Phytoplankton bloom dynamics of the western Ross Sea ice edge – I. Primary productivity and species-specific production, Deep-Sea Res. Pt. I, 33, 1375–1387, https://doi.org/10.1016/0198-0149(86)90041-5, 1986. a, b
Wright, S. W., van den Enden, R. L., Pearce, I., Davidson, A. T., Scott, F. J., and Westwood, K. J.: Phytoplankton community structure and stocks in the Southern Ocean (30–80∘ E) determined by CHEMTAX analysis of HPLC pigment signatures, Deep-Sea Res. Pt. II, 57, 758–778, https://doi.org/10.1016/j.dsr2.2009.06.015, 2010. a
Short summary
We observed that variation in the Southern Annular Mode (SAM) over 11 years showed a relationship with the species composition of hard-shelled phytoplankton in the seasonal ice zone (SIZ) of the Southern Ocean. Phytoplankton in the SIZ are productive during the southern spring and summer when the area is ice-free, with production feeding most Antarctic life. The SAM is known to be increasing with climate change, and changes in phytoplankton in the SIZ may have implications for higher life forms.
We observed that variation in the Southern Annular Mode (SAM) over 11 years showed a...




Altmetrics
Final-revised paper
Preprint